
Abstract
Monoclonal antibodies (MAbs) against epizootic hemorrhagic disease virus (EHDV) were produced by immunizing BALB/c mice with rec-VP7-EHDV2; 66 clones producing MAbs able to recognize the VP7-EHDV with a strong reaction were obtained and tested in indirect enzyme-linked immunosorbent assay (i-ELISA) against the whole epizootic hemorrhagic disease (EHD) virus serotype 2; potential cross-reactions with related orbiviruses, as Bluetongue virus (BTV) and African horse sickness virus (AHSV), were investigated as well by i-ELISA, Western blot, and immunofluorescence. Fifty-three MAbs were specific for EHDV (VP7 recombinant protein and whole virus) and 13 reacted also with the VP7 of BTV. None of the MAbs reacted with AHSV. MAbs specific for EHDV were further characterized in a competitive ELISA (c-ELISA): 20 among them were found useful to develop a c-ELISA for the detection of antibodies against EHDV in bovine sera. The availability of this extensive set of MAbs provides the opportunity to develop a c-ELISA for the serological diagnosis of EHDV and to tune new methods for the isolation and identification of the virus in biological samples and cell cultures. The experimentation protocol was approved by the Italian Ministry of Health (number 639/2018-PR, Resp. to Prot. BDF16.13#295833199#).
Introduction
Epizootic hemorrhagic disease (EHD) is a noncontagious infectious viral disease affecting domestic and wild ruminants, transmitted by vector insects of the genus Culicoides. It mainly affects white-tailed deer (Odocoileus virginianus) and occasionally cattle, sheep, and goats. In susceptible species, clinical manifestations of the disease are similar to those induced by Bluetongue virus (BTV) infection. Sheep and goats usually do not develop evident symptoms.(1,2)
Epizootic hemorrhagic disease virus (EHDV) belongs to the genus Orbivirus, family Reoviridae, and is closely related to BTV and African horse sickness virus (AHSV). At least seven serotypes are currently recognized around the world, but to date, the exact number of serotypes is not known.(1–3)
The viral particles consist of a double capsid with icosahedral symmetry; the genome consists of 10 genomic segments (S1 to S10) of double-stranded RNA encoding 7 structural proteins (VP1–VP7) and at least 4 nonstructural proteins (NS1, NS2, NS3, and NS3A).(1) The structural proteins VP2 and VP5 constitute the outer capsidic layer and are mainly responsible for binding with receptors and penetration into host cells.(4) The internal capsid is composed of two proteins VP7 and VP3.(1,5)
VP7 is the serogroup-specific orbivirus immunodominant viral protein, it is highly conserved among EHDV serotypes and shows a high percentage of identity at the amino acid level(6); for this reason, it is the ideal protein for the development of diagnostic tests.
Serological methods currently recommended for the diagnosis of EHD are competitive enzyme-linked immunosorbent assay (c-ELISA) and virus neutralization; complement fixation test and agar gel immunodiffusion could also be used in particular situations.(2)
EHD was first described in New Jersey (USA) in a white-tailed deer in 1955.(7) The virus is endemic in many areas of North America, Australia, and some Asian countries. In the last decade, EHDV has been detected in several countries of the Mediterranean basin, including Israel, Morocco, Algeria, Tunisia, and Turkey.(1,8–12)
Unlike BTV and AHSV, EHDV has never been reported in the European Union (EU); however, EHD outbreaks in the Mediterranean basin represent a serious risk for the EU. In fact, EHDV shares the same biological niche and transmission cycle as BTV whose forays into Southern Europe have been linked to viral circulation in North Africa.(13–15)
This study describes the production of a panel of monoclonal antibodies (MAbs) against the EHDV VP7 protein; these MAbs could be an important tool for the serological diagnosis of EHD, in particular for the development of a c-ELISA and for the identification of the virus in biological samples and cell cultures.
Materials and Methods
Ethics approval
Animal experimentation was carried out in compliance with the Italian national law, that is, Legislative Decree March 4, 2014, number 26,(16) implementing Directive 2010/63/UE of the European Parliament and the Council of the European Union on the protection of animals used for scientific purposes.(17) The protocol was approved by the Italian Ministry of Health with number 639/2018-PR (Resp. to Prot. BDF16.13#295833199#).
MAb production
Four female BALB/c mice (Charles River Breeding Laboratories), 6/8 weeks of age, were inoculated with rec-VP7-EHDV2 (IZSA&M) expressed in baculovirus-infected Sf9 cells and purified by immobilized metal affinity chromatography. The antigen, diluted with sterile 0.01 M phosphate-buffered saline (PBS), pH 7.4, to a protein concentration of 200 μg/mL, was emulsified with complete Freund adjuvant (Sigma-Aldrich) and administered intraperitoneally (0.5 mL/mouse). The second immunization (100 μg protein/mouse) was performed 14 days later with incomplete Freund adjuvant (Sigma-Aldrich), whereas the subsequent injection (100 μg protein/mouse) was performed 28 days later with sterile PBS only. Three days after the third injection, mice sera were tested for seroconversion in indirect ELISA (i-ELISA), and 42 days later, the first injection, the mice with the highest antibody titers were given an intravenous injection (100 μg protein/mouse).
Mice splenocytes were fused with murine myeloma cells Sp2/O-Ag-14 (ATCC CRL-1581™) using 50% polyethylene glycol solution (PEG 1550; Serva Feinbiochema GmbH & Co).(18) Hybridomas were grown for 2 weeks in Dulbecco's modified Eagle's medium (Sigma-Aldrich) containing 20% fetal bovine serum (Carlo Erba), 2 mM glutamine, 100 × penicillin-streptomycin-amphotericin B, 50 mg/mL gentamicin, 10,000 UI/mL nystatin, and 50 × HAT (HybriMax®; Sigma-Aldrich) and screened by i-ELISA for the presence of antibodies against rec-VP7-EHDV2 (2 μg/mL). Mouse serum samples collected before and after immunization were used as negative and positive controls, respectively. Hybridomas showing optical density values at 450 nm (OD450nm) ≥ 1.000 were cloned, according to the method of limiting dilutions,(19,20) to three sets of limiting dilution to ensure clonality.
Indirect enzyme-linked immunosorbent assay
Cross-reactions of MAbs versus rec-VP7-EHDV2 with related orbiviruses, such as BTV and AHSV, were assessed by i-ELISA. Briefly, 96-well microplates (PolySorp, Nunc Brand Products) were coated with EHDV serotype 2 purified whole virus (20 mg/mL), rec-VP7-BTV2 (IZSA&M) (2 μg/mL),(21) and rec-VP7-AHSV1 (IZSA&M) (2 μg/mL),(18) diluted in 0.05 M carbonate-bicarbonate buffer (CBB), pH 9.6, and incubated overnight (ON) at +4°C.
After blocking with 1% yeast extract in PBS containing 0.05% Tween 20 (PBST) at room temperature (RT) for 1 hour, 100 μL/well of MAb supernatants diluted 1:50 in PBST were added and incubated for 1 hour at 37°C. As secondary antibody, sheep anti-mouse IgG horseradish peroxidase (HRP) conjugated (100 μL/well; GE Healthcare) was used at the dilution of 1:20,000 in PBST at RT for 30 minutes. After washings, 100 μL/well of a chromogenic substrate, 3,3′,5,5′-tetramethylbenzidine (TMB; Surmodics), was added to each well for 10 minutes and the reaction stopped with 0.5 N H2SO4. Microplate reading was performed at 450 nm.
Competitive ELISA
Suitability of produced MAbs for the use in c-ELISA was assessed. Supernatants were twofold diluted in PBST and titrated by i-ELISA as described above: the dilution giving an OD450nm of 2.000 ± 0.100 was established as the titer of each MAb supernatant. Ninety-six-well microplates (Polysorp, Nunc) were coated with 2 μg/mL of rec-VP7-EHDV2 (IZSA&M) (100 μL/well) diluted in CBB and incubated ON at +4°C. Plates were blocked with 1% yeast in PBST for 1 hour at RT and then washed. Fifty microliters per well of an EHDV-positive bovine serum (VN titer 1:20, serotype 2) undiluted and twofold diluted in PBST (from 1:2 to 1:128), 50 μL/wells of an EHDV-negative bovine serum, and 50 μL/wells of PBST (MAb control and blank) were added to the plate and incubated for 1 hour at 37°C.
After further washings, 50 μL/well of each MAb at the established titer was added to the wells, except in blank wells, where 50 μL/well of PBST was added. Plates were incubated at 37°C for 1 hour and then washed. Fifty microliters per well of the Sheep anti-mouse IgG HRP-conjugated (GE Healthcare) diluted 1:20,000 in PBST was added to all the wells and incubated for 30 minutes at RT. Plates were then washed and added with 100 μL/well of TMB (Surmodics) for 10 minutes at RT. Colorimetric reaction was stopped by adding 50 μL/well of 0.5 N H2SO4. OD were acquired at 450 nm.
Percentage of MAb competition was calculated, after subtraction of the mean of blank OD, by the following formula:
Western blot analysis
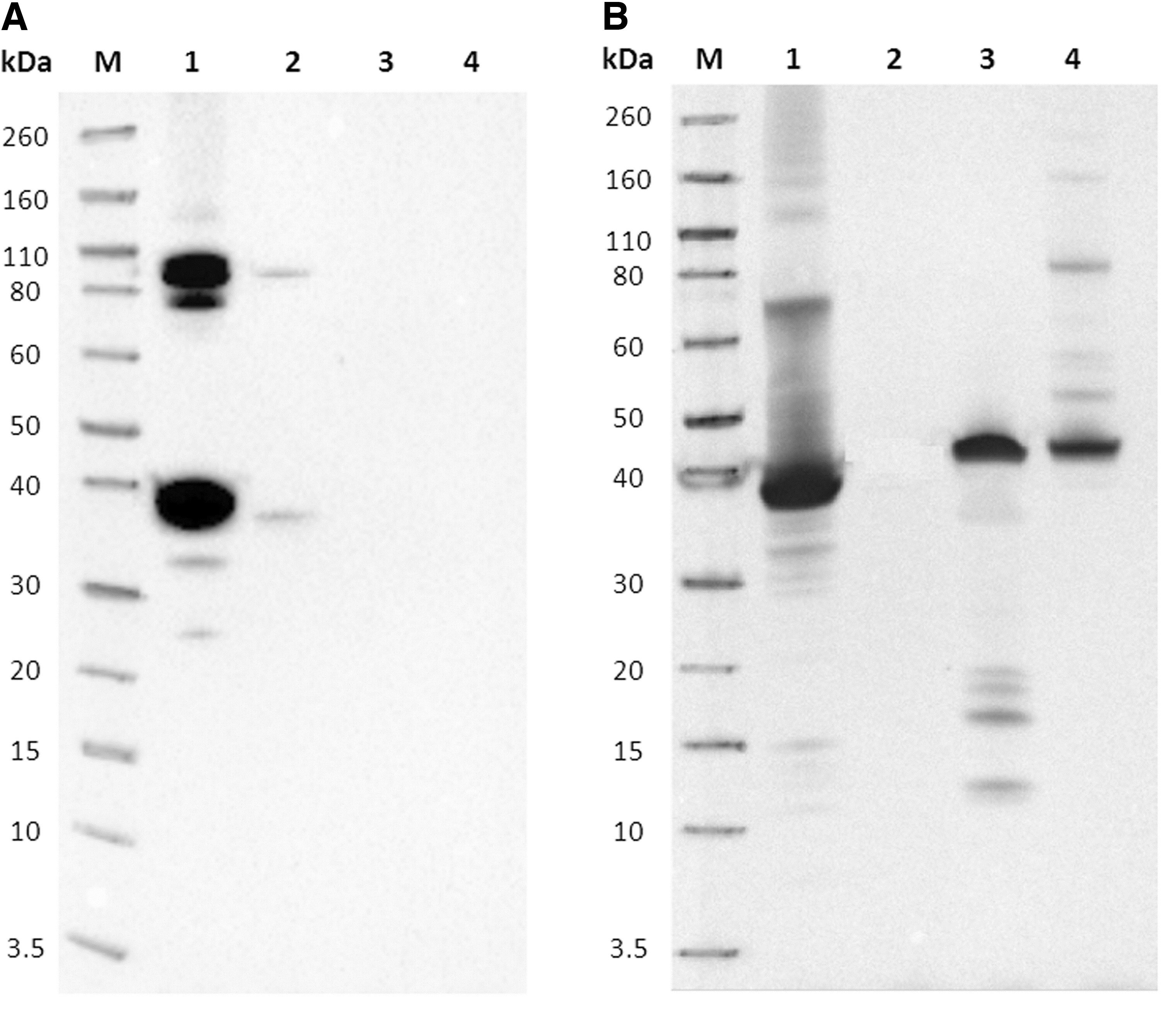
Proteins of EHDV serotype 2 purified whole virus (IZSA&M) (20 μg/20 μL), rec-VP7-EHDV2 (IZSA&M) (20 μg/20 μL), rec-VP7-BTV2 (IZSA&M) (20 μg/20 μL), and rec-VP7-AHSV1 (IZSA&M) (20 μg/20 μL) were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) at 200 V using NuPAGE 4%–12% Bis-Tris Gels Mini (Novex; Life Technologies). Gels were stained with Simply Blue Safe Stain (Life Technologies) to visualize protein bands.
Separated proteins were also transferred onto nitrocellulose membranes with iBlot Dry Blotting System (Life Technologies). Membranes were blocked with 5% skim milk (Biolife) in PBST for 2 hours at RT and cut into strips. Strips were incubated ON at 4°C with the MAb supernatants diluted in PBST containing 2.5% skim milk. The detection of immune complexes was performed using the Sheep anti-mouse IgG HRP conjugated (GE Healthcare) and a chemiluminescent substrate (ECL Select Western Blotting Detection Reagent; GE Healthcare). Results were analyzed using the ChemiDoc MP (Bio-Rad) and the Image Lab Software, version 4.0 (Bio-Rad).
MAb isotyping and large-scale production
MAbs specific for EHDV (whole virus and VP7 recombinant protein) and suitable for the use in c-ELISA were isotyped using the Mouse-Typer Isotyping Panel (Bio-Rad). MAbs with IgG isotype were produced in vitro on a large scale(18) and purified by affinity chromatography using a column HiTrap rProtein A FF, 5 mL (GE Healthcare), according to the manufacturer's instructions. Purified MAbs were concentrated with 100 kDa cutoff centrifugal filters (Millipore) and resuspended in PBS; the protein concentration was determined by spectrophotometry (absorbance at 280 nm/IgG molar extinction coefficient). Purified MAbs were stored at −20°C until use.
Affinity assay
Affinity assay was performed according to Antes(22) with some modifications. Ninety-six-well microplates (PolySorp, Nunc) were coated with rec-VP7-EHDV2 (IZSA&M) (2 μg/mL in CBB) and blocked as previously described. Fifty microliters per well of each purified MAb serially diluted in PBST from 50 to 0.02 μg/mL was added and incubated for 1 hour at 37°C. Secondary antibody Sheep anti-mouse IgG HRP conjugated (GE Healthcare), TMB (Surmodics), and 0.5 N H2SO4 were applied as previously described.
Avidity assay
Avidity assay was performed according to Antes(22) with some modifications.
Ninety-six-well microplates (PolySorp, Nunc) were coated with rec-VP7-EHDV2 (IZSA&M) serially diluted in CBB from 50 to 0.02 μg/mL and blocked as previously described. Fifty microliters per well of each purified MAb at the concentration of 5 μg/mL in PBST was added (incubation 1 hour at 37°C). Secondary antibody Sheep anti-mouse IgG HRP conjugated (GE Healthcare), TMB (Surmodics), and 0.5 N H2SO4 were applied as previously described.
Immunofluorescence assay
Vero cells were grown in Nunc® Lab-Tek® Chamber Slide™ system (Corning). After reaching cellular confluence, the cells were infected with BTV serotype 2, EHDV serotype 2, and AHSV serotype 1. Vero cells uninfected and infected with BTV serotype 2 and AHSV serotype 1 were used as negative controls. After 48 hours, the glass slides were fixed with cold acetone for 20 minutes, air dried, and stored at 4°C. Thirty microliters of each purified MAb diluted 1:50 in PBS was added on the surface of the test wells and incubated at 37°C for 30 minutes.
The slides were washed two times with PBS for 10 minutes each, and air dried. Antigen-MAb reaction was revealed using 30 μL of fluorescein-conjugated rabbit IgG fraction to mouse IgG (MP Biomedicals), diluted 1:80 in PBS containing 1% Evans Blue (Sigma-Aldrich). Slides were washed as above, rinsed in distilled water, air dried, and evaluated by fluorescence microscopy (40 × magnification, DMI3000B; Leica).
Results
MAb characterization by i-ELISA
Sixty-six clones producing MAbs showing a strong reaction (OD450nm > 2.000) versus rec-VP7-EHDV2, and reacting both with EHDV VP7 recombinant protein and EHD whole virus, were obtained. Among them, 53 MAbs did not cross-react with BTV and AHSV VP7 recombinant proteins and 13 MAbs showed a strong (OD450nm > 1.500) or weak (0.300 < OD450nm < 0.700) cross-reaction with the VP7 of BTV. None of the 66 MAbs reacted with AHSV VP7.
Competitive ELISA
The 53 MAbs specific for EHDV (recombinant VP7 and whole virus) were tested by c-ELISA using a two-fold diluted EHDV-positive bovine serum. Twenty of them were able to compete with antibodies in the EHDV bovine positive serum for the binding to VP7 protein epitopes. Eight of these MAbs (1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, 7E3B5, 8C7B9, and 8E4C3) showed the best results in terms of percentage of competition and were selected for further characterization.
Supernatant titers of selected MAbs are showed in Table 1.
Characterization of Monoclonal Antibodies Versus Epizootic Hemorrhagic Disease Virus
Dilution giving an OD450nm of 2.000 ± 0.100.
c-ELISA, competitive enzyme-linked immunosorbent assay; mAb, monoclonal antibody; OD, optical density.
MAb isotyping and large-scale production
The 20 MAbs useful in c-ELISA showed k light chain and the following isotypes: IgG1 (8 MAbs), IgG2a (4 MAbs), and IgG2b (8 MAbs). MAbs 1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, 7E3B5, 8C7B9, and 8E4C3, showing the best results in c-ELISA, were produced on a large scale and purified by affinity chromatography. Antibody production (mg of purified IgG obtained per liter of supernatant) and isotypes of the eight selected MAbs are shown in Table 1.
Western blotting
MAbs 1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, 7E3B5, 8C7B9, and 8E4C3 recognized, both in rec-VP7-EHDV2 and the EHD whole virus, two protein bands showing apparent molecular weight of about 36 and 110 kDa, corresponding, respectively, to the monomer and the trimer of VP7. Rec-VP7-BTV2 and rec-VP7-AHSV1 were not recognized (Fig. 1A, B).
Affinity and avidity assay
MAbs 7E3B5, 8E4C3, 8C7B9, 2D7D9, and 5A11D12 showed the greatest affinity and avidity; MAb 1G4B6 showed the lowest affinity and avidity. MAbs 1B4B4 and 1B4B6 produced intermediate results (Figs. 2 and 3).
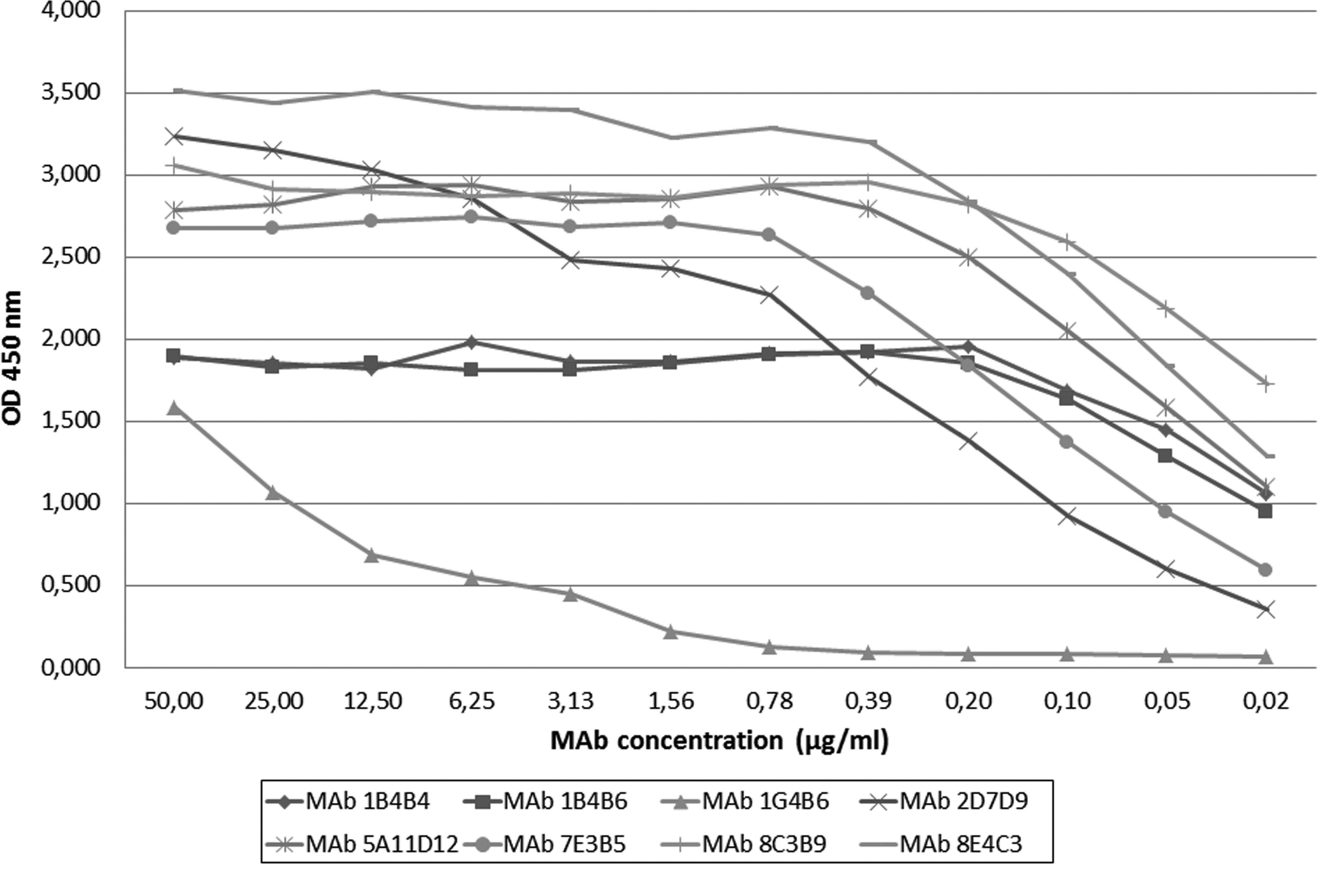
Affinity assay of MAbs 1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, 7E3B5, 8C7B9, and 8E4C3 versus the rec-VP7-EHDV2 (IZSA&M).
Avidity assay of MAbs 1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, 7E3B5, 8C7B9, and 8E4C3 versus the rec-VP7-EHDV2 (IZSA&M).
Immunofluorescence assay
Vero cells infected with EHDV serotype 2 and incubated with MAbs 1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, and 7E3B5 showed a strong specific fluorescence in the cytoplasm (Fig. 4). No fluorescence was observed in the slides coated with noninfected Vero cells and cells infected with BTV serotype 2 and AHSV serotype 1 (Images not shown). EHDV-infected cells incubated with MAbs 8C7B9 and 8E4C3 showed a weak fluorescence (Image not shown).

Immunofluorescence with MAbs 1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, and 7E3B5. Vero cells infected with EHDV serotype 2.
Discussion and Conclusions
EHDV and BTV viruses both belong to the genus Orbivirus, infect ruminants, and show similarities in the life cycle and in the way of transmission.(11,23) Co-infections of animals with the two viruses have been reported in areas where EHDV and BTV co-circulate. In the last years, several BTV serotypes have been detected in European countries and in some countries of the Mediterranean basin, but to date, EHDV outbreaks have been reported only in Tunisia, Israel, Egypt, and Turkey.(24) Circulation of EHDV in North Africa represents a remarkable risk for Europe, since the two viruses also share the same species of vector insects(11,25,26) and show a similar clinical pattern in susceptible species.(2) For these reasons, it is necessary to have good diagnostic tools for detection and discrimination of EHDV and BTV. MAbs represent an important tool to improve the diagnosis of infectious diseases.
In this work, a panel of eight MAbs against the VP7 protein of EHDV was produced. Selection of the most useful MAbs was conducted taking into account the specificity for EHDV and the reaction with both the purified VP7 protein and the whole virus and with both the native and the denatured form of the VP7, to make possible the use of the selected MAbs for different diagnostic methods (ELISA, immunofluorescence, immunohistochemistry, SDS-PAGE, and Western blotting). Regarding the specificity, no clone reacting with AHSV was obtained, but 13 out of the 66 clones showing a strong reaction with the VP7 EHDV reacted with the VP7 of BTV. This is due to the fact that VP7 of EHDV and BTV show a high degree of homology (>80%) in their amino acid sequence.(27)
According to the OIE Manual,(2) competitive ELISA is the preferred serological test for the detection of EHDV antibodies, so the usefulness of MAbs in c-ELISA was assessed. All the eight selected MAbs (1B4B4, 1B4B6, 1G4B6, 2D7D9, 5A11D12, 7E3B5, 8E4C3, and 8C7B9) were able to compete with antibodies of a positive bovine serum and could be used, after conjugation with HRP, to develop a c-ELISA. The eight MAbs resulted specific for EHDV and, being directed versus the VP7 protein, common to all the serotypes, they should be able to recognize all the serotypes of the EHDV and be useful to detect the virus in the event of a suspected outbreak.
The next step of this work will be the development of a c-ELISA using the eight HRP-conjugated MAbs to select the MAb providing the better values of sensibility and specificity of the test.
Authors' Contributions
All authors have contributed to, seen, and approved this final, submitted version of the article. The article is not under consideration in any other journal.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.