
Abstract
CC chemokine receptor 3 (CCR3) belongs to the G protein-coupled receptor family and is highly expressed in eosinophils and basophils. CCR3 is essential for recruiting eosinophils into the lung. Moreover, CCR3 was found in the serum of colorectal cancer patients higher than in the control group. Therefore, CCR3 will be a useful target for asthma and colorectal cancer diagnosis. This study developed a specific and sensitive monoclonal antibody (mAb) for mouse CCR3 (mCCR3), which is useful for flow cytometry using the Cell-Based Immunization and Screening method. The established anti-mCCR3 mAb, C3Mab-3 (rat IgG2a, kappa), reacted with mCCR3-overexpressed Chinese hamster ovary-K1 (CHO/mCCR3) cells through flow cytometry. C3Mab-3 also reacted with P388 (mouse lymphoid neoplasma) and J774-1 (mouse macrophage-like) cells, which express mCCR3 endogenously. Kinetic analyses using flow cytometry indicated that KDs of C3Mab-3 for CHO/mCCR3, P388, and J774-1 cells were 4.3 × 10−8 M, 2.6 × 10−7 M, and 2.4 × 10−7 M, respectively. C3Mab-3 could be a valuable tool for elucidating mCCR3-related biological response using flow cytometry.
Introduction
CC chemokine receptor 3
Therefore, chemokines and their receptors are attractive diagnostics and therapeutic targets, and some chemokine- and chemokine receptor-targeted drugs have been launched. For example, plerixafor is the CXCR4 receptor antagonist. Plerixafor is an immunostimulant and is used for patients with lymphoma and multiple myeloma. Mogamulizumab, a monoclonal antibody (mAb) against CCR4, is used as an anticancer drug.(12,13) Maraviroc, a CCR5 antagonist, is used as an antiretroviral drug that blocks HIV entry.(10,14,15)
Chemokines are classified into four subfamilies: CC, CXC, CX3C, and XC.(16,17) CC type chemokines, such as eotaxin family (CCL11, CCL24, and CCL26), Monocyte Chemotactic Protein-3 (MCP-3; CCL7), and regulated on activation, normal T cell expressed and secreted (RANTES; CCL5), are known as ligands for CCR3. CCR3 support HIV Env-mediated syncytia formation, and eotaxin inhibits the efficiency of HIV-1 infection.(2,18,19) CCR3 also contributes to ocular allergy.(20) Moreover, CCR3 is essential for the recruitment of eosinophils into the lung and airway hyperresponsiveness in a murine model of allergic asthma.(21–24)
A recent study indicated that the CCR3 concentration in the serum of colorectal cancer patients was significantly higher than the control group.(25) Therefore, CCR3 is one of the most important chemokine receptors in asthma and will be useful as a biomarker for diagnosing colorectal cancer. Moreover, single and repeated intrathecal administration of SB328437, CCR3 antagonists, decreased mechanical and thermal hypersensitivity caused by nerve injury, and blockade of CCR3 by SB328437 enhanced morphine and buprenorphine effectiveness; therefore, CCR3 will be a promising target for neuropathic pain therapy.(26)
The development of specific anti-CCR3 mAbs has been required for allergic and HIV therapy and pathological diagnosis of age-related macular degeneration. However, the development of anti-GPCR mAbs has been challenging because GPCRs have complicated three-dimensional structures, and have a small extracellular region. Purification of GPCRs for an antigen is complex.(27) In this study, a novel anti-mouse CCR3 (mCCR3) mAb was developed using the cell-based immunization and screening (CBIS) method(28–33) and investigated its usefulness for flow cytometry analysis, and determined the dissociation constant between the novel anti-mCCR3 mAb and mCCR3-expressed cell lines.
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1, LN229, and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA). P388 (mouse lymphoid neoplasma) and J774-1 (mouse macrophage-like) cells were obtained from the Cell Resource Center for Biomedical Research Institute of Development, Aging and Cancer Tohoku University (Miyagi, Japan). Synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding mCCR3 (Accession No. NM_009914.4) was subcloned into a pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). mCCR3 plasmid was transfected into CHO-K1 and LN229 cells using the Neon transfection system (Thermo Fisher Scientific Inc., Waltham, MA). Stable transfectants were established through cell sorting using a cell sorter (SH800; Sony Corp., Tokyo, Japan) and cultivation in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA).
CHO-K1, P3U1, mCCR3-overexpressed CHO-K1 (CHO/mCCR3), P388, and J774-1 were cultured in a Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan). LN229 and mCCR3-overexpressed LN229 (LN229/mCCR3) were cultured in Dulbecco's modified Eagle's medium (DMEM; Nacalai Tesque, Inc.). RPMI 1640 and DMEM were supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific Inc.), 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Cells were grown in a humidified incubator at 37°C with an atmosphere of 5% CO2 and 95% air.
Antibodies
An anti-mCCR3 mAb (clone J073E5) was bought from BioLegend (San Diego, CA). Secondary Alexa Fluor 488-conjugated anti-rat IgG was bought from Cell Signaling Technology, Inc. (Danvers, MA).
Production of hybridomas
Sprague Dawley (SD) rats (6 weeks old) were purchased from CLEA Japan (Tokyo, Japan). Animals were housed under specific pathogen-free conditions. All animal experiments were conducted according to relevant guidelines and regulations to minimize animal suffering and distress in the laboratory. Animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit No. 2019NiA-001). A rat was monitored daily for health during the full 4-week duration of the experiment. A reduction of more than 25% of total body weight was defined as a humane endpoint. A rat was euthanized through cervical dislocation, and death was verified through respiratory and cardiac arrest.
To develop mAbs against mCCR3, the CBIS method was employed. In brief, one SD rat was immunized with LN229/mCCR3 cells (1 × 109) by the intraperitoneal route together with the Inject Alum (Thermo Fisher Scientific Inc.). The procedure included three additional immunization, followed by a final booster intraperitoneal injection administered 2 days before the harvest of spleen cells. Harvested spleen cells were subsequently fused with P3U1 cells using PEG1500 (Roche Diagnostics, Indianapolis, IN). The hybridomas were grown in an RPMI 1640 medium supplemented with hypoxanthine, aminopterin, and thymidine for selection (Thermo Fisher Scientific Inc.). Flow cytometry was used to screen the supernatants with CHO/mCCR3, CHO-K1, P388, and J774-1 cells.
Flow cytometry
Cells were harvested after a brief exposure to 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). They were washed with 0.1% bovine serum albumin in phosphate-buffered saline and treated with primary mAbs for 30 minutes at 4°C. The cells were then treated with Alexa Fluor 488-conjugated anti-rat IgG (1:2000; Cell Signaling Technology, Inc.). Fluorescence data were obtained using BD FACSLyric (BD Biosciences, Franklin Lakes, NJ).
Determination of dissociation constants (KDs) by flow cytometry
CHO/mCCR3, P388, and J774-1 were suspended in 100 μL of serially diluted anti-mCCR3 mAbs, and then 50 μL of Alexa Fluor 488-conjugated anti-rat IgG (1:200; Cell Signaling Technology, Inc.) was added. Fluorescence data were obtained using the BD FACSLyric. The KDs were calculated by fitting saturation binding curves to built-in one-site binding models in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA).
Results
The development of anti-mCCR3 mAbs using the CBIS method
To develop anti-mCCR3 mAbs, we employed CBIS method. CBIS method includes the immunization of mCCR3-overexpressed cells and the high-throughput hybridoma screening using flow cytometry (Fig. 1). In this study, one rat was immunized with LN229/mCCR3 cells (Fig. 1A). The spleen was excised from the rat, and splenocytes were fused with P3U1 cells (Fig. 1B). Developed hybridomas were seeded into 96-well plates and cultivated for 6 days. Culture supernatants positive for CHO/mCCR3 were screened using flow cytometry (Fig. 1C).
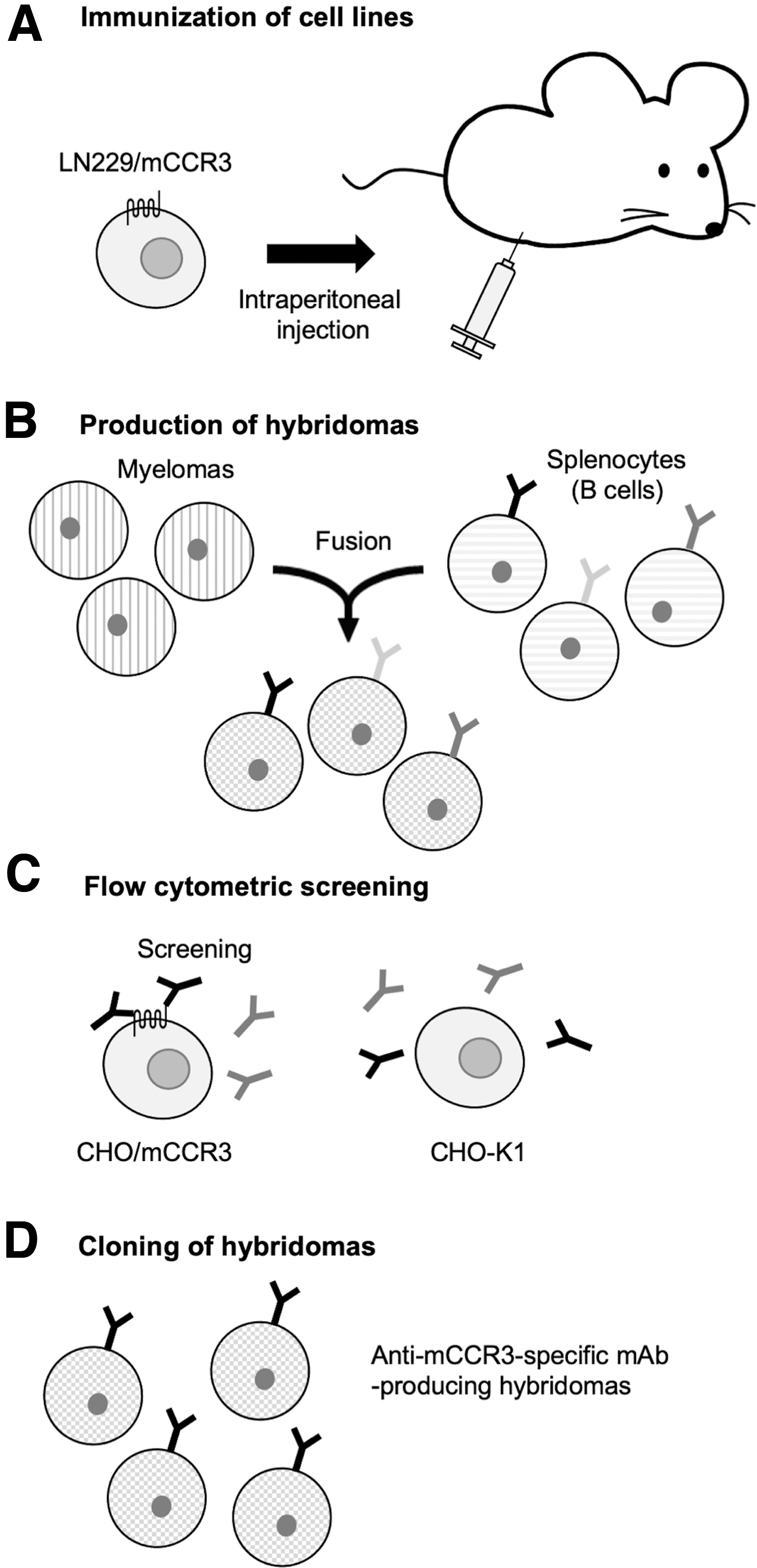
Schematic illustration of the production of anti-mCCR3 mAbs.
The first screening approach identified strong signals from CHO/mCCR3 cells and weak or no signals from CHO-K1 cells in 30 of 956 wells (3.14%). The second screening procedure identified strong signals from P388 and J774-1 cells in 26 of the 30 hybridoma supernatants identified in the previous step (86.7%). After limiting dilution and several additional screenings, an anti-mCCR3 mAb, C3Mab-3 (rat IgG2a, kappa) was finally established (Fig. 1D).
Flow cytometry analysis
We performed flow cytometry using C3Mab-3 against CHO/mCCR3, CHO-K1, P388, and J774-1. C3Mab-3 dose-dependently recognized CHO/mCCR3 cells at 10, 1, and 0.1 μg/mL (Fig. 2A, left panel), but not parental CHO-K1 cells (Fig. 2B, left panel). Another anti-mCCR3 mAb (clone J073E5 from BioLegend: positive control) also recognized CHO/mCCR3 in a dose-dependent manner (Fig. 2A, right panel), but not CHO-K1 (Fig. 2B, right panel). Although C3Mab-3 did not react with CHO/mCCR3 at 0.01 μg/mL, J073E5 reacted with it even at 0.01 μg/mL (Fig. 2A). C3Mab-3 and J073E5 also recognized P388 cells (Fig. 2C) and J774-1 cells (Fig. 2D) expressing endogenous mCCR3 in a dose-dependent manner. These results suggested that C3Mab-3 is specific for mCCR3 and is useful for detecting endogenous mCCR3 using flow cytometry.
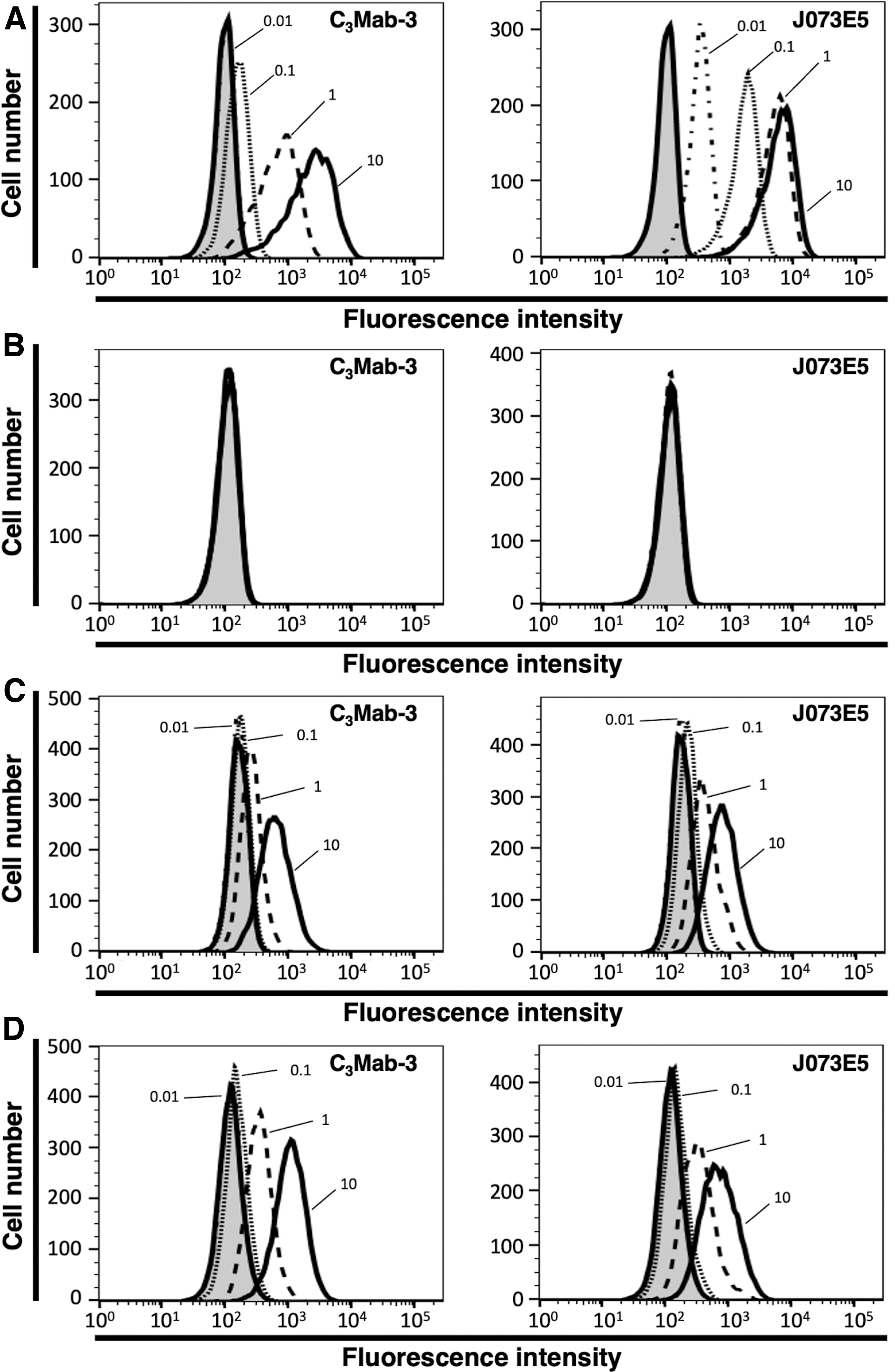
Flow cytometry using anti-mCCR3 mAbs. CHO/mCCR3
Determination of the KDs using C3Mab-3 against mCCR3-expressing cells by flow cytometry
To assess the KD of C3Mab-3 with mCCR3-expressed cells, we performed kinetic analysis of the interaction of C3Mab-3 with CHO/mCCR3, P388, and J774-1 using flow cytometry. The geometric mean of fluorescence intensity was plotted versus the concentration of C3Mab-3 and fitted by one-site binding models in GraphPad PRISM 8. The KDs of C3Mab-3 for CHO/mCCR3, P388, and J774-1 cells were determined to be 4.3 × 10−8 M (Fig. 3A), 2.6 × 10−7 M (Fig. 3B), and 2.4 × 10−7 M (Fig. 3C), respectively. These results show that C3Mab-3 possesses moderate affinity for CHO/mCCR3 cells and low affinity for P388 and J774-1.
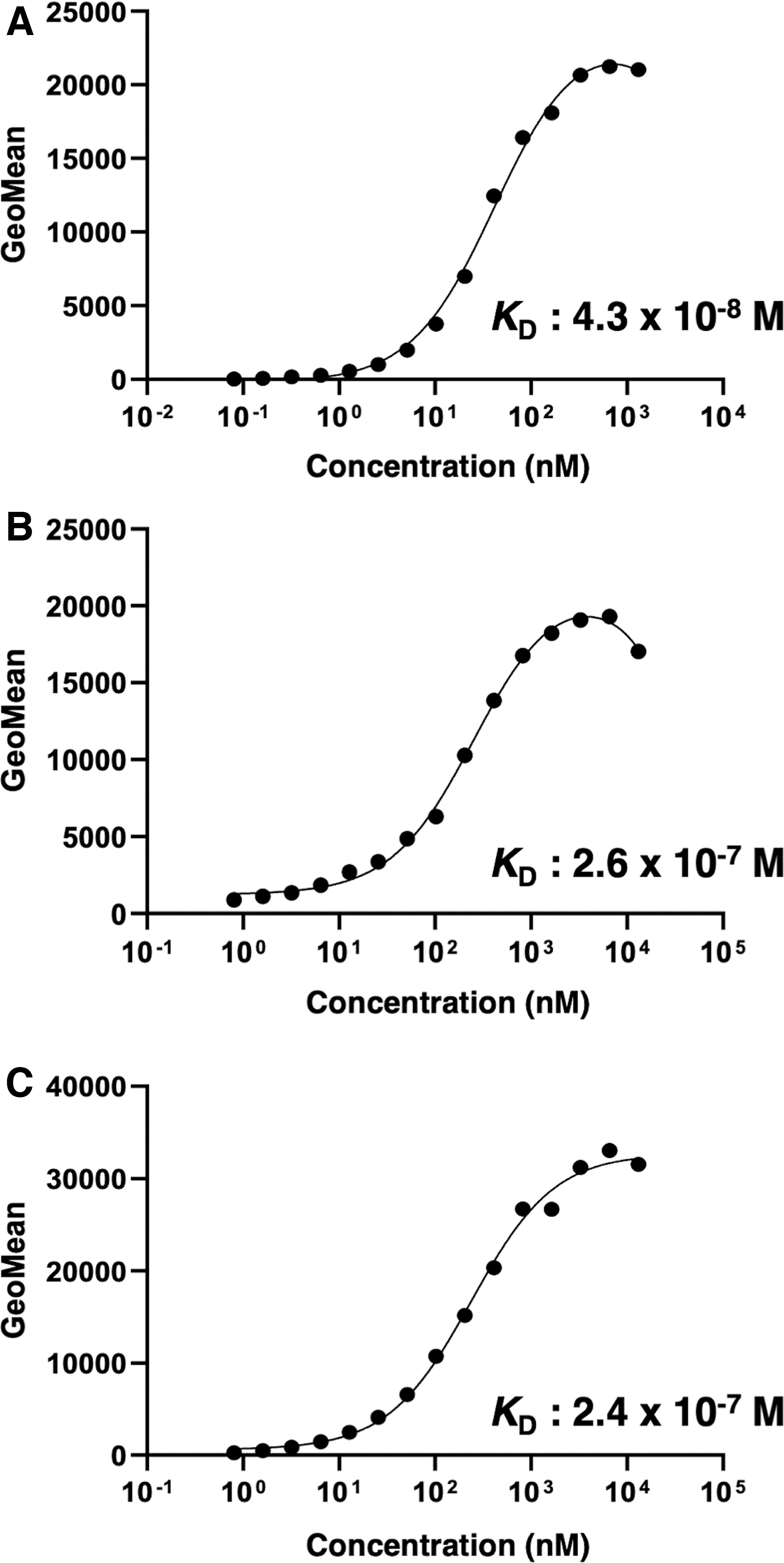
Determination of the KDs of C3Mab-3 against mCCR3-expressing cells. CHO/mCCR3
Discussion
GPCRs respond to different external signals, such as light, hormones, and chemokines, and transmit extracellular signals to intracellular molecules. GPCR-driven signaling pathways are involved in physiological function and the pathogenesis of several diseases. Therefore, GPCRs are therapeutic targets. Therefore, anti-GPCR mAbs are a valuable tool for therapy and diagnosis. To generate the antibodies, purified proteins or synthesized peptides are often used as an immunogen. However, the purification of GPCR is complex, and consumes much time and effort. Although synthesized peptides are easy to obtain, established mAbs using immunizing peptides often do not recognize proteins that have a native conformation.(34)
Previously, we have succeeded in the development of useful mAbs against membrane proteins, including CCR3,(31) CCR8,(32) CCR9,(33) podoplanin,(35–38) CD20,(39,40) CD44,(30) EpCAM,(41) TROP2,(42) EGFR,(43) and HER3,(44) using CBIS method in which antigen-expressing cell lines are used for immunization and screening. In this study, the novel anti-mCCR3 mAb, C3Mab-3, which is useful for flow cytometry using the CBIS method. C3Mab-3 reacted with endogenous mCCR3 in P388 and J774-1 cells similar to the J073E5 (BioLegend). In contrast, C3Mab-3 reacted with CHO/mCCR3 cells, but this reactivity is weaker than J073E5 with CHO/mCCR3 (Fig. 2).
In a previous study, other anti-mCCR3 mAbs, C3Mab-2, was developed using immunizing CHO/mCCR3 cell lines.(31) The reactivities of C3Mab-3 with CHO/mCCR3, P388, and J774-1 cell lines are comparable with that of C3Mab-2 with these cell lines. In addition, KDs between C3Mab-3 and these cell lines are also comparable with KDs between C3Mab-2 and them. However, C3Mab-2 cannot detect trypsin-treated CHO/mCCR3 (data not shown). Otherwise, C3Mab-3 can react with trypsin-treated CHO/mCCR3. This result indicates that C3Mab-3 and C3Mab-2 recognize different regions on mCCR3. In future studies, the epitope regions of C3Mab-3 and C3Mab-2 will need to be determined. Moreover, it will be confirmed whether C3Mab-3 is useful for Western blotting, immunohistochemistry, and enzyme-linked immunosorbent assay.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers JP22ama121008 (to Yukinari Kato), JP21am0401013 (to Yukinari Kato), and JP21am0101078 (to Yukinari Kato).