
Abstract
The epidermal growth factor receptor (EGFR) contributes to tumor malignancy through gene amplification and/or protein overexpression. In our previous study, we developed an anti-human EGFR (hEGFR) monoclonal antibody, clone EMab-134 (mouse IgG1, kappa), which specifically detects both hEGFR and dog EGFR (dEGFR). The defucosylated mouse IgG2a version of EMab-134 (134-mG2a-f) exhibits antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) in dEGFR-overexpressed Chinese hamster ovary-K1 (CHO/dEGFR) cells and antitumor activities in mouse xenografts of CHO/dEGFR cells. In this study, the reactivity of 134-mG2a-f against a canine mammary gland tumor cell line (SNP) was examined by flow cytometry and immunocytochemistry. Furthermore, 134-mG2a-f highly exerted ADCC and CDC for SNP. The administration of 134-mG2a-f significantly suppressed the SNP xenograft growth. These results suggest that 134-mG2a-f exerts antitumor effects against dEGFR-expressing canine mammary gland tumors, and could be valuable as part of an antibody treatment regimen for them.
Introduction
Female breast cancer has the highest number of worldwide new cases of all diagnosed cancers, with 2,261,419 cases (11.7% of new all cancer cases) and 684,996 deaths (6.9% of the total new cancer deaths) in 2020.(1) Breast cancer deaths have steadily decreased, but their diagnostic rates and deaths are globally high.(2)
Canines are one of the animals that spontaneously develop cancers, including breast cancers. In terms of histological, genetic, and biological features, dog has strong similarities to human.(3–6) Dogs share carcinogenic factors, such as aging, exposure to carcinogens, obesity, and environmental risk factors with humans.(7,8) Dogs grow five to eight times faster than humans, making them suitable as models for age-related gene profiling and pathogenesis research.(6)
The gene amplification of MYC oncogene and the loss of tumor suppressor gene PTEN have been found in both human and canine breast cancers.(9) The tumor suppressor gene P53 abnormalities also have been found in several canine tumors, including mammary tumors.(10,11) Dysregulations in these genes are often found in human cancers. Among mutations found in human breast cancers, BRCA1, IGF2R, and USH2A mutations have also been confirmed in dog cancer.(12) Estrogen receptor (ER)+ canine mammary tumors have been reported as complex carcinomas that include both luminal and myoepithelial cells. In contrast, ER-tumors are classified into simple carcinomas arise from genomic aberrations.(12)
Response to cancer treatment, acquisition of drug resistance, metastasis, and recurrence were similarly observed in both dogs and humans.(13) Due to its similarity, human cancer research is expected to be applied into canine.(7,14) It has been reported that inhibition of programmed cell death-1 (PD-1) pathway is one of the therapeutic strategies in canine metastatic oral malignant melanoma.(15)
Epidermal growth factor receptor (EGFR) is a type I glycoprotein that forms a dimer by binding its ligand and regulates cell proliferation and survival. Overexpression and mutations of EGFR activate multiple intracellular cascades, such as RAS-MAPK and PI3K/AKT pathways.(16) EGFR is closely associated with various cancer development and malignancy in both human and dog.(17–21) EGFR expression also correlates with poor prognosis in several canine cancers, including mammary cancer.(22–24) Human EGFR (hEGFR) and dog EGFR (dEGFR) have 91% amino acid homology.(25) Therefore, the development of therapeutic methods targeting EGFR is suitable as a cancer treatment for dog.
In our previous study, we developed an anti-hEGFR monoclonal antibody (mAb), clone EMab-134 (mouse IgG1, kappa).(26) The 134-mG2a, an IgG2a type of EMab-134, exerted antitumor activities in model of human oral squamous cell carcinoma.(27) Furthermore, a 134-mG2a-f, defucosylated type of 134-mG2a, possesses antitumor effects in mouse xenograft models of dEGFR-expressed cells.(28) The aim of this study is to investigate whether 134-mG2a-f possesses antibody-dependent cellular cytotoxicity (ADCC), complement-dependent cytotoxicity (CDC), and antitumor activities using a canine mammary gland tumor model.
Materials and Methods
Cell lines
A canine mammary gland tumor cell line, SNP, was obtained from the Cell Resource Center for Biomedical Research Institute of Development, Aging and Cancer at Tohoku University (Miyagi, Japan).(29) SNP was cultured in Roswell Park Memorial Institute-1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific, Inc., Waltham, MA), 100 U/mL of penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Cells were cultured at 37°C in a humidified atmosphere containing 5% CO2.
Antibodies
Anti-hEGFR mAb EMab-134 (mouse IgG1, kappa) was developed as previously described.(26) To generate recombinant EMab-134 (recEMab-134), we subcloned VH and CH of cDNAs of EMab-134 into the pCAG-Neo vector, along with VL and CL cDNAs of EMab-134 into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan), respectively. To generate 134-mG2a, we subcloned VH cDNA of EMab-134 and CH mouse IgG2a into the pCAG-Neo vector, along with VL and CL cDNAs of EMab-134 into the pCAG-Ble vector, respectively.(27) Vectors of EMab-134 and 134-mG2a were transfected into ExpiCHO-S cells and BINDS-09 cells (FUT8-deficient ExpiCHO-S cells) using the ExpiCHO Expression System (Thermo Fisher Scientific, Inc.), respectively.(28) The resulting mAbs, recEMab-134 and 134-mG2a-f, were purified with Protein G-Sepharose (GE Health care Biosciences, Pittsburgh, PA).(28) Mouse IgG (cat. no. I8765) and IgG2a (cat. no. M7769) were purchased from Sigma-Aldrich (St. Louis, MO).
Flow cytometry
SNP cells were harvested by brief exposure to 0.25% trypsin/1 mM ethylenediamine tetraacetic acid (EDTA; Nacalai Tesque, Inc.). After washing with blocking buffer of 0.1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS), cells were treated with 1 μg/mL of recEMab-134, 134-mG2a-f, or control blocking buffer for 30 minutes at 4°C. Then, cells were incubated in Alexa Fluor 488-conjugated anti-mouse IgG at a dilution of 1:1000 (cat no. 4408S; Cell Signaling Technology, Inc., Danvers, MA) for 30 minutes at 4°C. Fluorescence data were collected using the EC800 Cell Analyzer (Sony Corp., Tokyo, Japan).
Determination of binding affinity
SNP cells were suspended in 100 μL of serially diluted 134-mG2a-f (0.006–100 μg/mL) followed by Alexa Fluor 488-conjugated anti-mouse IgG (1:200; Cell Signaling Technology, Inc.). Fluorescence data were collected using the EC800 Cell Analyzer (Sony Corp.). The dissociation constant (KD) was calculated by fitting binding isotherms to built-in one-site binding models in GraphPad Prism 8 (GraphPad Software, Inc., La Jolla, CA).
Immunocytochemical analysis
Subconfluent SNP cells cultured on acid-wash coverslips were fixed in 4% paraformaldehyde in PBS for 10 minutes at room temperature. After quenching with 50 mM NH4Cl in PBSc/m (PBS containing 0.2 mM Ca2+ and 2 mM Mg2+) for 10 minutes, the cells were blocked with blocking buffer (PBSc/m containing 0.5% BSA) for 30 minutes and incubated with 10 μg/mL of recEMab-134, 134-mG2a-f, or control blocking buffer for 1 hour. Then, the cells were labeled with Alexa Fluor 488-conjugated anti-mouse IgG (1:400; Cell Signaling Technology, Inc.) for 45 minutes. The cell nuclei were stained with 4′,6-diamidino-2-phenylindole (Thermo Fisher Scientific, Inc.). A fluorescence microscope BZ-X800 (Keyence, Osaka, Japan) was used to acquire fluorescence images.
Animals
All animal experiments were performed following relevant guidelines and regulations to minimize animal suffering and distress in the laboratory. Animal experiments for ADCC and antitumor activity were approved by the Institutional Committee for Experiments of the Institute of Microbial Chemistry (permit nos. 2021-028 for ADCC assays and 2021-021 for antitumor experiments). Mice were maintained in a specific pathogen-free environment (23°C ± 2°C, 55% ± 5% humidity) on an 11 hours light/13 hours dark cycle with food and water supplied ad libitum across the experimental period. Mice were monitored for health and weight every 2–5 days during the 3-week period of each experiment. We determined the loss of original body weight to a point >25% and/or a maximum tumor size >3000 mm3 as humane endpoints for euthanasia. Mice were euthanized by cervical dislocation; death was verified by respiratory and cardiac arrest.
Antibody-dependent cellular cytotoxicity
A total of five female 5-week-old BALB/c nude mice (weighing 14–17 g) were purchased from Charles River Laboratories, Inc., (Kanagawa, Japan). Spleen cells from five mice were used as the source of natural killer cells for the evaluation of ADCC, as previously reported.(28) Following euthanasia by cervical dislocation, the spleens were removed aseptically, and a syringe was used to force spleen tissues through a sterile cell strainer (352360; BD Falcon, Corning, New York, NY) and obtain single-cell suspensions. Erythrocytes were lysed by 10-s exposure to ice-cold distilled water. Splenocytes were washed with Dulbecco's modified Eagle's medium (DMEM; Nacalai Tesque, Inc.) and resuspended in DMEM with 10% FBS to be used as effector cells. SNP cells were labeled with 10 μg/mL Calcein AM (Thermo Fisher Scientific, Inc.) and resuspended in the same medium.
The target cells (2 × 104 cells/well) were plated in 96-well plates and mixed with effector cells (effector/target cells ratio, 50), 100 μg/mL of 134-mG2a-f or control mouse IgG2a. Following 4.5-hour incubation at 37°C, the release of Calcein into the supernatant was measured in each well. The fluorescence intensity was determined using a microplate reader (Power Scan HT; BioTek Instruments, Inc., Winooski, VT) with an excitation wavelength of 485 nm and an emission wavelength of 538 nm. Cytolytic activity (% lysis) was calculated as follows: % lysis = (E−S)/(M−S) × 100, where “E” is the fluorescence measured in combined cultures of target and effector cells, “S” is the spontaneous fluorescence of target cells only, and “M” is the maximum fluorescence measured following the lysis of all cells with a buffer containing 0.5% Triton X-100, 10 mM Tris-HCl (pH 7.4), and 10 mM of EDTA.
Complement-dependent cytotoxicity
SNP cells were labeled with 10 μg/mL Calcein AM (Thermo Fisher Scientific, Inc.) and resuspended in the medium. They were then plated in 96-well plates at 2 × 104 cells/well with rabbit complement (final dilution 1:10; Low-Tox-M Rabbit Complement; Cedarlane Laboratories, Hornby, Ontario, Canada) and 100 μg/mL of 134-mG2a-f or control mouse IgG2a. Following 4 hours of incubation at 37°C, we measured Calcein release into the supernatant for each well. Fluorescence intensity was calculated as described in the ADCC section above.
Antitumor activity of 134-mG2a-f in xenografts of SNP cells
A total of 16 female BALB/c nude mice (5 weeks old, weighing 14–17 g) were purchased from Charles River Laboratories, Inc., and used in experiments once they reached 7 weeks of age. SNP cells (0.3 mL of 1.33 × 108 cells/mL in DMEM) were mixed with 0.5 mL BD Matrigel Matrix Growth Factor Reduced (BD Biosciences, San Jose, CA); 100 μL of this suspension (5 × 106 cells) was injected subcutaneously into the left flanks of the mice. On day 7 postinoculation, 100 μg of 134-mG2a-f (n = 8) or control mouse IgG (n = 8) in 100 μL PBS were injected intraperitoneally. Additional antibody inoculations were performed on days 11 and 18. At 25 days following cell implantation, all mice were euthanized by cervical dislocation. Tumor diameters and volumes were determined as previously described.(28)
Statistical analyses
All data are expressed as mean ± standard error of the mean. Statistical analysis was conducted with Tukey's test for ADCC and CDC, and Welch's t test for tumor weight. Analysis of variance (ANOVA) and Sidak's multiple comparisons tests were conducted for tumor volume and mouse weight. All calculations were performed using GraphPad Prism 8 (GraphPad Software, Inc.). A p-value of <0.05 was considered statistically significant.
Results
Flow cytometry analysis against canine mammary gland tumor cell line, SNP cells using recEMab-134 and 134-mG2a-f
In our previous study, an anti-hEGFR mAb, EMab-134, recognized dEGFR-overexpressed CHO/dEGFR cells, indicating that EMab-134 crossreacts with dEGFR.(28) In this study, recEMab-134 also detected canine mammary gland tumor cell line, SNP cells (Fig. 1A). Similarly, the defucosylated mouse IgG2a type of EMab-134, 134-mG2a-f, detected SNP cells (Fig. 1B), indicating that both recEMab-134 and 134-mG2a-f detect endogenous dEGFR-expressed on SNP cells.
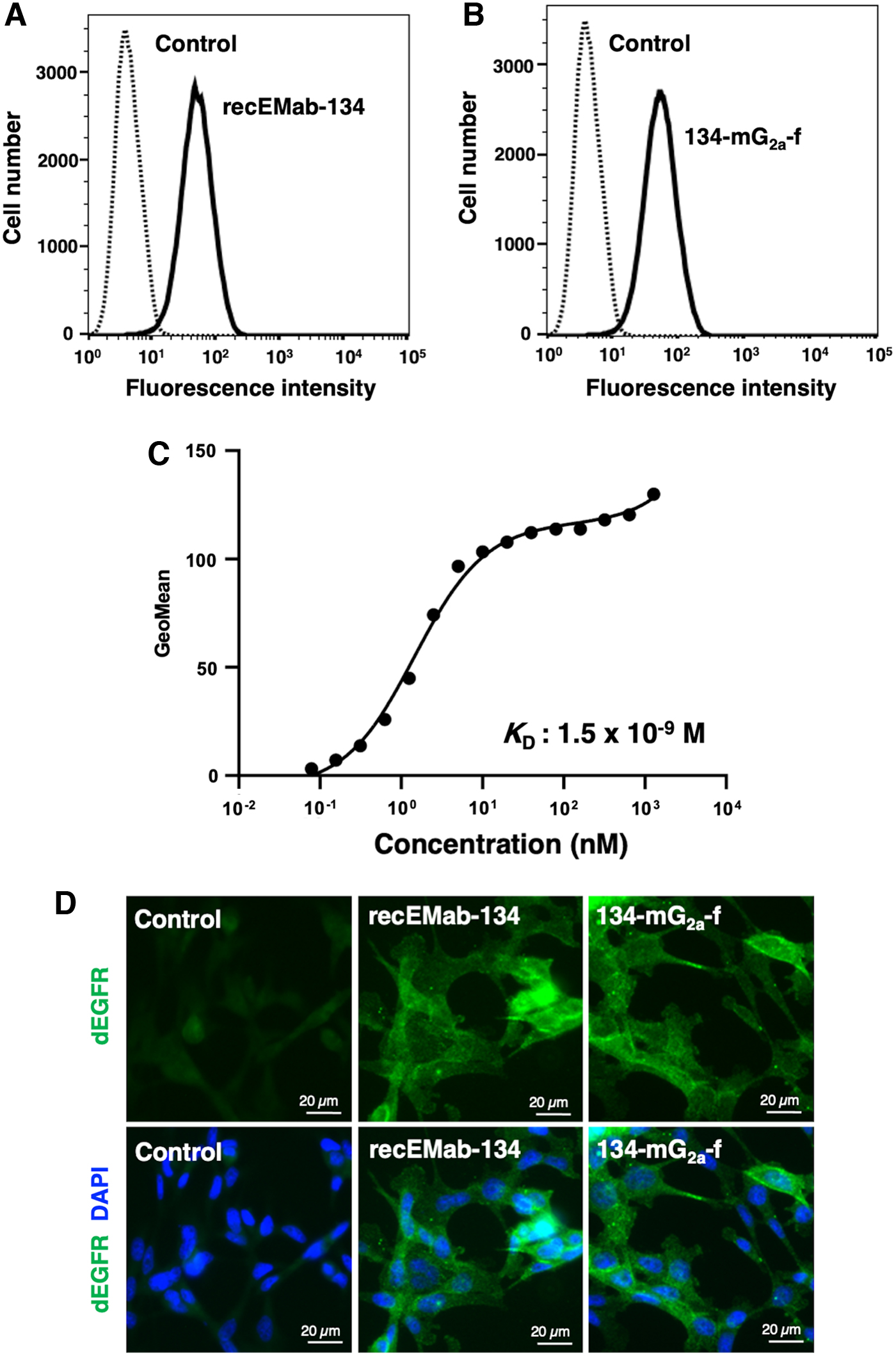
Flow cytometry and immunocytochemistry using recEMab-134 and 134-mG2a-f.
Determination of binding affinity
A kinetic analysis of the interactions of 134-mG2a-f with SNP cells was performed via flow cytometry. As shown in Figure 1C, the KD for the interaction of 134-mG2a-f with SNP cells was 1.5 × 10−9 M, suggesting that 134-mG2a-f shows high affinity for SNP cells.
Immunocytochemical analysis against canine mammary gland tumor cell line, SNP cells using recEMab-134 and 134-mG2a-f
RecEMab-134 and 134-mG2a-f were applied to immunocytochemistry in SNP cells. As shown in Figure 1D, recEMab-134 and 134-mG2a-f, but not buffer control, visualized dEGFR in SNP cells, suggesting that recEMab-134 and 134-mG2a-f recognize endogenous dEGFR in SNP cells.
134-mG2a-f-mediated ADCC and CDC in SNP cells
We next investigated whether 134-mG2a-f has ADCC against SNP cells. As shown in Figure 2A, 134-mG2a-f showed ADCC (46.8% cytotoxicity) against SNP cells more potent than mouse IgG2a (12.9% cytotoxicity) and PBS (7.8% cytotoxicity).
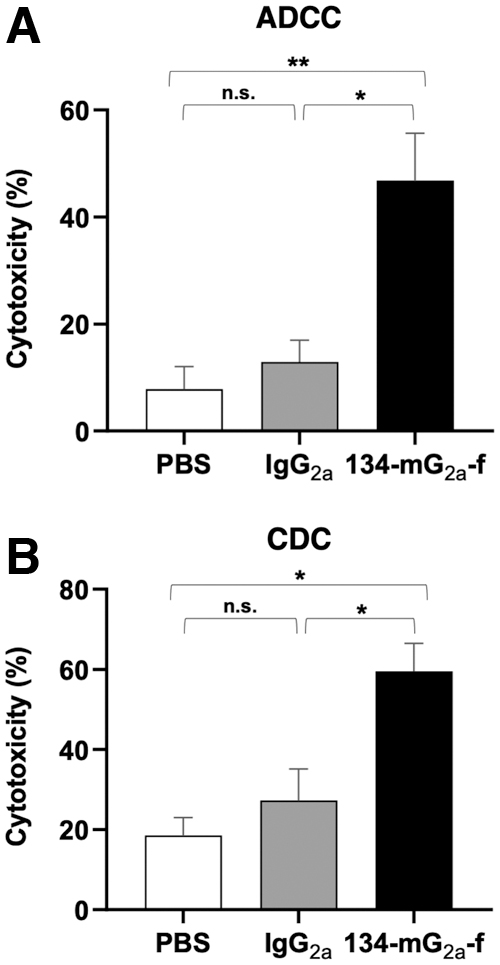
Evaluation of ADCC and CDC elicited by 134-mG2a-f.
We then investigated whether 134-mG2a-f has CDC against SNP cells. As shown in Figure 2B, 134-mG2a-f elicited a higher degree of CDC (59.5% cytotoxicity) in SNP cells compared with that elicited by mouse IgG2a (27.3% cytotoxicity) and PBS (18.5% cytotoxicity). These results demonstrated that 134-mG2a-f exerts ADCC and CDC against dEGFR-expressing SNP cells.
Antitumor activities of 134-mG2a-f in the mouse xenografts of SNP cells
In the SNP xenograft models, 134-mG2a-f (100 μg) and control mouse IgG (100 μg) were injected intraperitoneally into mice on days 7, 11, and 18 following the injection of SNP cells. The tumor volume was measured on days 7, 11, 14, 18, 21, and 25 after the injection. The administration of 134-mG2a-f resulted in a significant reduction in tumor development on days 18 (p < 0.05), 21 (p < 0.05), and 25 (p < 0.01) compared with that of the control mouse IgG (Fig. 3A). The administration of 134-mG2a-f resulted in a 31% reduction of tumor volume compared with that of the control mouse IgG on day 25 postinjection. Furthermore, the tumor weight of the 134-mG2a-f-treated mice was significantly lower than that of IgG-treated mice (21% reduction; p < 0.05, Fig. 3B). Total body weights of the two groups did not change significantly (Fig. 3C).
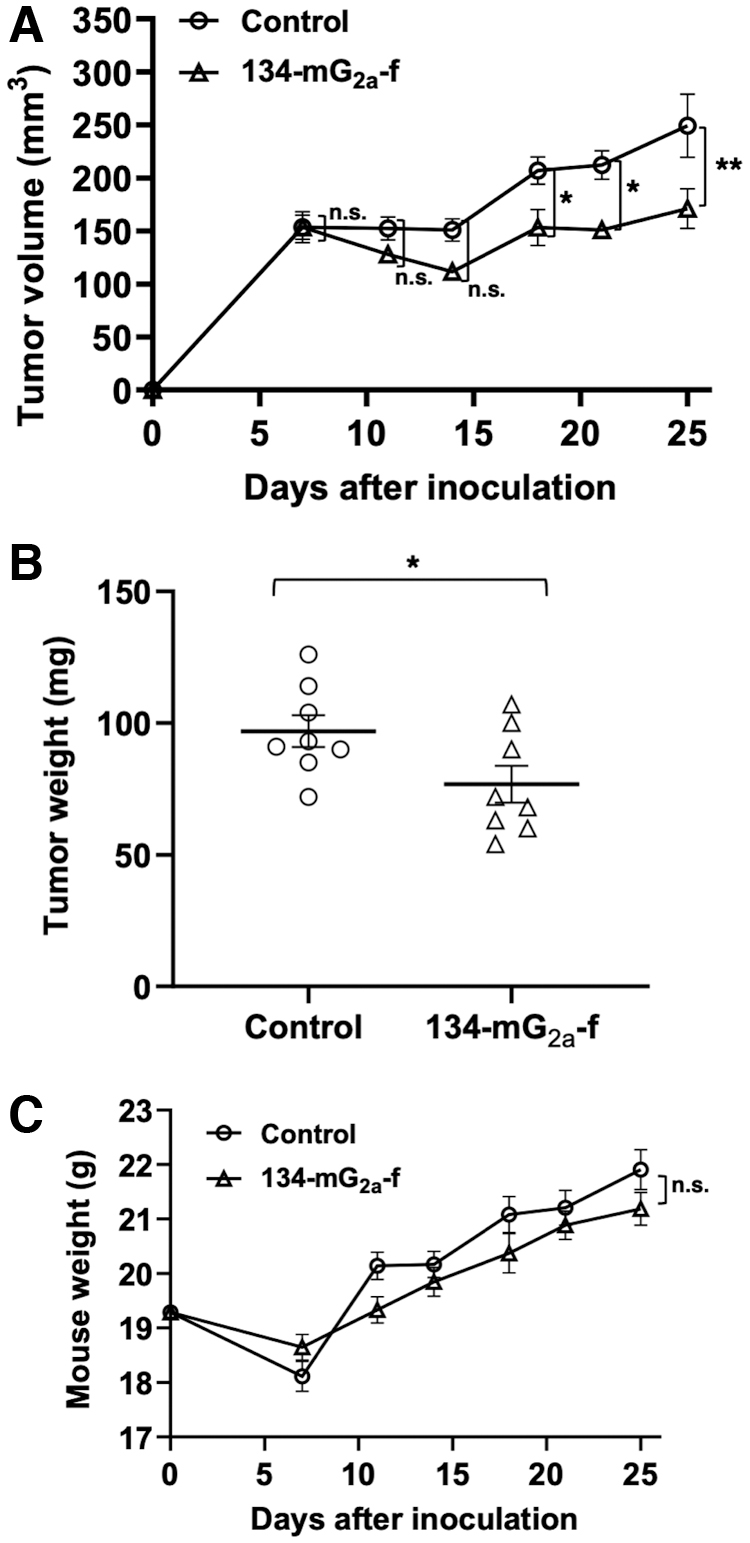
Evaluation of antitumor activity of 134-mG2a-f in SNP xenografts.
Taken together, these results indicate that the administration of 134-mG2a-f effectively suppresses the tumor growth of SNP xenografts.
Discussion
EGFR is one of the important oncoproteins that promotes tumor development.(16) Therapeutic agents targeting EGFR have been developed and clinically applied.(30,31) The EGFR-targeted antibody drugs, including cetuximab and panitumumab,(32,33) possess similar binding affinities to EGFR, but have different epitopes on the EGFR.(34–36) These mAbs can inhibit the EGFR signaling pathways and induce apoptosis in EGFR-expressing cancer cells and show therapeutic potential for patients with advanced colorectal cancer.(37,38)
Approved-drugs for human are often used to canine cancer treatment due to their similarities.(7,39) Since higher expression levels of EGFR is involved in canine cancer malignancy and poor prognosis, including lung, nasal, mammary, and transitional cell carcinoma,(18,22–24) EGFR-targeting drugs are considered as important therapeutic strategy. We have previously shown that the 134-mG2a-f has an antitumor effect against a dEGFR-overexpressed CHO model.(28) This study also demonstrated antitumor activities of a defucosylated IgG2a type of anti-hEGFR mAb (134-mG2a-f) on in vitro and in vivo in canine mammary gland tumor cells expressed endogenous dEGFR (Figs. 2 and 3). In the future study, we will develop the antibody-based immunotherapeutic strategies such as chimeric antigen receptor (CAR)-T cell therapy using 134-mG2a-f.(40)
Human epidermal growth factor receptor 2 (HER2) is one of the oncogenes that belongs to the EGFR family of receptor tyrosine kinases consisting of EGFR (HER1, ERBB1), HER2 (ERBB2), HER3 (ERBB3), and HER4 (ERBB4).(41) HER2 is frequently confirmed to be overexpressed in human and canine breast cancers.(42) Previously, we have also reported that a specific mAb against HER2 (H2Mab-41) exerts strong antitumor activity against dog HER2-overexpressed cells.(43) Thus, the combination therapy of 134-mG2a-f with H2Mab-41 might be effective for the treatment of canine cancer clinically.
Combination therapy of cetuximab and pembrolizumab is successful for recurrent or metastatic human head and neck squamous cell carcinomas.(44) Pembrolizumab is an immune checkpoint inhibitor that recognizes immunosuppressive molecule PD-1 expressed on T cells, and has been expanded to various human cancers in recent years.(45) Not only immune checkpoint inhibitors but also CAR-T cell therapy is used to treat canine cancers.(46) Therefore, the combination of anti-EGFR antibody therapy and immune checkpoint inhibitor is expected to be more effective in canine cancer treatment. Further studies are required for the investigation of antitumor activity of 134-mG2a-f to spontaneously develop canine cancer and the establishment of technique to diagnose canine cancer, including dEGFR expression and immune microenvironment.
Identified cell surface markers for the canine immune cell subsets such as lymphoid, myeloid, and myeloid-derived suppressor cells are approximately identical to humans.(47,48) Therefore, dogs with an immune system similar to humans have been focused as companion animals suitable for evaluation of cancer immunotherapies.(49) PD-1 is expressed on total of 5%–10% CD4+ T cells and 20%–25% CD8+ T cells in healthy dogs and is upregulated in canine cancer.(50) Since anti-PD-1 and anti-PD-L1 mAbs are used in canine cancer treatment,(15,51,52) the combination of immune checkpoint inhibitors with the other antibody drug is expected to be more effective in canine cancer treatment in the future trials. Due to its genetical and histological similarity, canine cancer research can provide information feedback not only for development of canine cancer treatment but also for human cancer research.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported, in part, by Japan Agency for Medical Research and Development (AMED) under grant nos.: JP21am0401013 (to Y.K.) and JP21am0101078 (to Y.K.).