
Abstract
We investigated the potential for vancomycin-susceptible Enterococcus (VSE) from multi-component salads to disseminate vanA from four clinical vancomycin-resistant enterococci to 14 streptogramin-resistant enterococci (SRE) of food and animal origin. Strains were selected from a previous study based on the presence of streptogramin susceptibility and/or vanA, vanB, vatD, vatE, agg, cpd, and gelE genes. Transconjugants were isolated on brain heart infusion agar containing vancomycin and selective antibiotics. Thirty-nine matings using a 1:10 donor-recipient ratio for filter and broth methods resulted in transfer of vanA between an agg+cpd+gelE+ Enterococcus faecalis donor and an agg−cpd−gelE− streptogramin-susceptible Enterococcus faecium salad recipient at a frequency of 10−8 per recipient by filter method. Secondary mating of the transconjugant with SR/VSE strains resulted in a two- to four-log-fold greater frequency of transfer. Reverse transcription–polymerase chain reaction revealed vanA RNA products in the transconjugant cultivated in nutrient broth and salad at 37°C in the presence and absence of recipient filtrate. This study demonstrated that native salad VSE disseminated vanA to SRE carrying agg, cpd, and/or gelE. An increase in transfer efficiency resulted from secondary conjugation using the native vancomycin- and streptogramin-susceptible salad strain as the donor.
Introduction
The vanA gene, conferring high-level resistance to vancomycin, is carried on transposon Tn1546 and can be transferred via a conjugative plasmid. 17 Dissemination of vanA via enterococcal probiotic or starter strains has been demonstrated in vitro.10,20 High levels of streptogramin-resistant enterococci (SRE) isolated among VanA-type Enterococcus from poultry and human sources, as well as transfer of vanA to food-associated Enterococcus faecalis strain during sausage and cheese fermentations in the absence of selected antibiotic pressure is known to occur.7,16 The emergence of VRE over the past decades leaves few treatment options for patients, resulting in significant increases in length of inpatient stay and cost of patient care. 25 Dissemination of glycopeptide resistance elements has been well documented, but the role of commensal bacteria in the dissemination of antibiotic resistance genes via foods is limited. 31 The recent horizontal transfer of vanA from clinical VRE to methicillin-resistant Staphylococcus aureus (MRSA) has produced novel clinical pathogens with high-level resistance to vancomycin.3,30
Although fecal carriage of VRE among healthy people in the United States is relatively low, a relatively large European community reservoir exists in both humans and animals due to the former agricultural use of avoparcin.8,26 Concerns that similar resistance to current antibiotics used to treat VRE, including the streptogramin compound quinupristin-dalfopristin (Q/D; Synercid), will emerge due to the continued use of streptogramin growth-promoting antibiotics in the United States such as virginiamycin. 27 Evolutionary genetic studies of VRE and vancomycin-susceptible enterococci (VSE) from human and nonhuman sources have indicated that a common genetic lineage exists among hospital-related Enterococcus faecium globally. 32
In a previous study, we characterized the vancomycin and streptogramin resistance profiles of commensal enterococci isolated from multi-component prepared deli salads. 4 This work examined the potential for enterococci isolated from multi-component salads to serve as an intermediary in the dissemination of vanA between clinical enterococci and enterococci of food and animal origin. Mating experiments were conducted in broth, on filters, and in salad at 37°C to test the conjugative potential of transconjugant enterococci. The expression of VanA in a transconjugant strain was tested by reverse transcriptase–polymerase chain reaction (RT-PCR).
Materials and Methods
Bacterial strains
Eighteen donor and recipient enterococci were chosen from a pool of isolates that included hospitalized inpatient, animal, and food strains. Clinical isolates were selected among those obtained from positive blood cultures of hospitalized inpatients at Robert Wood Johnson University Hospital, New Brunswick, New Jersey, from September through October 2005; food enterococci were obtained from multi-component deli salads, and animal enterococci isolated from cows or milk from cows with clinical mastitis and were a generous gift from Dr. Joseph Hogan (OARDC Mastitis Laboratory, Ohio State University).
Identification and characterization of bacterial isolates
All isolates were identified to the species level with a 99% or greater probability using a Microscan Gram-Positive/MIC Combo Panel and Walk/Away 96 SI system (Dade Behring, Sacramento, CA) and antimicrobial susceptibilities were interpreted according to Clinical and Laboratory Standards Institute guidelines for broth microdilution methods. 6 Species identification was verified by detection of sodA using species-specific primers. The presence of vancomycin resistance determinants vanA, vanB, virginiamycin acetyltransferase genes vatD, vatE and the selected genes agg (aggregation substance), cpd (sex pheromone), gelE (gelatinase), efaAfm (adhesin) were determined in a previous study. 3 Additionally, sodA (superoxide dismutase) was included as a housekeeping gene for RT-PCR. High-level vancomycin resistance for all enterococci and endpoints for quinupristin-dalfopristin resistance for E. faecium were determined by E-test (AB Biodisk, Piscataway, NJ) on Mueller-Hinton agar (Difco, Sparks, MD). Minimal inhibitory concentration of virginiamycin and fusidic acid was determined by standard agar diffusion testing on Mueller-Hinton medium containing 0.125 to 32 μg/ml virginiamycin M1 (Sigma, St. Louis, MO) or 20 μg/ml fusidic acid (Axxora LLC, San Diego, CA). Agar diffusion and Etests for minimal inhibitory concentration (MIC) determination were performed in duplicate. A previously described multiplex PCR procedure was used for detecting antibiotic resistance genes and virulence determinants. 4 Isolates and transconjugants were screened for plasmids using the Wizard Genomic Purification Kit (Promega, Madison, WI). Control strains for antimicrobial susceptibility and PCR included E. faecium ATCC 51559 (vanA+efaAfm+), E. faecalis ATTC 51299 (vanB+agg+gelE+cpd+), E. faecium CVM 3001 (vatE+), and E. faecium CVM 3002 (vatD+). In mating experiments, the rifampin- and fusidic acid-resistant strain E. faecalis JH2-2 was included as a reference recipient. 19 E. faecium CVM 3001 and CVM 3002 were also used as virginiamycin-resistant recipient strains in secondary mating experiments.
Bacteriocin production
Bacteriocin production was determined by the well-diffusion assay. In brief, cell-free supernatants (CFS) of isolates C2, S4, T2G, S6, B12, CVM3001, CVM3002, T7A, T7B, T7D, and T7E were prepared from brain heart infusion (BHI) broth cultures incubated at 10°C, 22°C, and 37°C overnight. The cultures were centrifuged (10 min, 5,400 rpm), supernatant decanted, and passed through a 0.2 μm filters. Each isolate listed above served as an indicator organism. Micrococcus luteus used as a standard based on its known sensitivity to bacteriocins. The negative control was BHI and nisin (10 mg/ml) (Sigma, 2.5% bacteriocin preparation [106 IU/g] dissolved in ddH2O) as the positive control. The indicator organism for the well-diffusion and disk diffusion assays was inoculated onto TSA (containing 6% yeast extract) as a lawn using a sterile swab, and allowed to air-dry for 5 min. For the well diffusion assay 10-mm wells were punched into the agar using a sterile glass tube, and 200 μl of CFS was added. The plates were kept at room temperature for 2 hr to allow for absorption of the supernatant, and then incubated at 37°C overnight. For the disk diffusion assay, 6 mm sterile filter paper disks were placed onto the surface of the inoculated agar plates. A 20-μl drop of CFS was dispensed onto individual disks and the plates kept at room temperature for 1 hr to allow absorption of the supernatant, and then incubated at 37°C overnight.
Preparation of cells with and without E. faecium S4 culture filtrate
Culture filtrate from E. faecium S4 was prepared for testing the effect of primary recipient pheromones in mating experiments. Cells were grown at 37°C in BHI broth to OD600 = 1. Cultures were centrifuged for 10 min at 5,400 rpm; supernatant was collected by filtration through a 0.22 μm-pore size Millipore membrane and diluted 50% with BHI broth. Donor and transconjugant cells were grown in BHI broth without antibiotics overnight and standardized to ∼109 CFU/ml (OD600 = 0.3–0.4). Five hundred microliters of the cell suspension or 250 μl cell suspension plus 250 μl E. faecium S4 filtrate was added to 4.5 ml sterile BHI broth and 1.5 g simulated salad, briefly vortexed, and incubated at 37°C for 4 hr without agitation. After incubation, 2 ml of BHI broth was added to tubes containing salad samples. Broth and salad tubes were vortexed, centrifuged (5 min; 10,000 rpm), and supernatant decanted. The pellet was suspended in 100 μl of RNAlater (Ambion, Austin, TX) and stored at −20°C.
Preparation of multi-component salad
Blended salad was prepared using the following commercial products: 60 g Hellmann's Light Mayonnaise (Unilever, Englewood Cliffs, NJ), 120 g DelMonte Diced New Potatoes (DelMonte, San Francisco, CA), and 130 g Swanson White Chicken Breast packed in water (Campbell Soup, Camden, NJ). Ingredients were processed in a sterile stomacher bag for 2 min, and 1.5 g aliquots were aseptically transferred to sterile 50 ml tubes. Control samples were incubated at 37°C and examined for sterility following subculture and incubation at 37°C on BHI agar for 24, 48, and 72 hr.
Mating experiments
All matings were performed in duplicate at 37°C. Broth matings were done according to the procedure by Clewell et al. 5 Briefly, overnight cells grown in BHI broth were mixed in a 1:10 donor–recipient ratio (50 μl donor and 500 μl recipient in 4.5 ml BHI broth) and incubated with agitation (150 rpm) at the appropriate temperature for 4 hr. Filter matings were performed using a 1:10 donor–recipient ratio diluted in BHI broth and spread onto a 0.22 μm pore size filter (Millipore, Billerica, MA) on BHI agar, then incubated overnight at the appropriate temperature. Cells were harvested by washing filters in 1 ml BHI and then spread-plated on media containing 6 μg/ml vancomycin HCl (Sigma), and the appropriate selective antibiotic: 20 μg/ml fusidic acid and 20 μg/ml rifampin (Sigma) and/or 32 μg/ml nitrofurantoin (Alfa Aesar, Ward Hill, MA) for primary transconjugants or 10 μg/ml tetracycline HCl (Acros Organics, Morris Plains, NJ) for secondary transconjugants. For additional verification that colonies were recipient rather than donor transconjugants, 3% gelatin was included in selective agar plates for recipient strains that exhibited the gelatinase phenotype. Plates were incubated in ambient air at 37°C for 72 hr and examined daily. From any plate exhibiting growth, one to six colonies were selected at random and subcultured to BHI agar. Presumptive transconjugants were identified to the species level by Microscan and the selected genetic determinants (vanA, vanB, vatD, vatE, agg, cpd, gelE, efaAfm, and sodA were verified by PCR.
RNA isolation and RT-PCR
For determination of vanA expression in primary donor and transconjugant cells, cells were grown to log phase in BHI broth and disrupted by incubation in 15 mg/L TE-lysozyme (Sigma) for 10 min, vortexing every 2 min. Total RNA was extracted using the Qiagen RNeasy Mini Kit (Qiagen, Valencia, CA). Samples were eluted from the column using 50 ml RNase/DNase-free water (Fermentas, Hanover, MD) and DNase-treated by adding 5 μl RQ1 10×DNase reaction buffer (Promega, Madison, WI), 5 μl RQ1 RNase-free DNase (Promega), and incubated 30 min at 37°C. RNA was quantified using spectrophotometric analysis at OD260. DNase-treated RNA samples (100–50 ng) were amplified with 0.5 μl of ImProm-II reverse transcriptase (Promega) in ImProm-II 5×reaction buffer (Promega) with 3 mM MgCl2, 300 mM dNTPs, 0.5 μl of GoTaq DNA polymerase (Promega) added, and 400 mM each forward and reverse primers in a total reaction volume of 30 μl. cDNA was synthesized at 48°C for 45 min, heated at 94°C for 2 min, and then amplified for 35 cycles with denaturing at 94°C for 30 sec, annealing at 55°C for 45 sec, elongation at 72°C for 1 min, and final extension of 7 min at 72°C. Reactions were carried out in a GeneAmp PCR system 2400 (Perkin-Elmer, Waltham, MA). Forward and reverse primer sequences for the species-specific variant of the superoxide dismutase gene sodA were based on previously published data. 18 Electrophoresis of 10 μl of RT-PCR product was performed on a 1.5% agarose gel containing 0.5 μl of ethidium bromide and observed using UV transillumination. E. faecium ATCC 51559 (vanA+) was used as the control strain.
Results
Characterization and selection of strains used in mating studies
All isolates were identified by Microscan to the species level with a 99% or greater probability. Strains used in mating experiments were based on appropriate antimicrobial susceptibility patterns for selection (Table 1). Three high-level vancomycin-resistant E. faecalis and one E. faecium isolates from clinical sources were selected as donors in mating experiments with vancomycin-susceptible recipient enterococci. For detection of transconjugants, E. faecium isolates demonstrating the appropriate resistance patterns (see Table 1) from salad and bovine sources were selected as recipients for secondary matings with the transconjugant. Susceptibility to virginiamycin and Q/D was the same for all isolates. Intrinsic resistance was observed in all E. faecalis, whereas three of the salad and two of the bovine E. faecium were streptogramin resistant. Gelatinase production on Todd-Hewitt agar with 3% gelatin was only observed in E. faecalis. Of those isolates in which gelE was detected, two (C1 and CVM 3002) were negative for gelatinase production. Virginiamycin acetyltransferase genes vatD and vatE were detected in none of the isolates other than E. faecium CVM 3001 and CVM 3002. Virulence determinants agg, cpd, and gelE genes were detected together in all but one (B13) E. faecalis regardless of source. The enterococcal antigen A determinant efaAfm was detected in all E. faecium isolates. In primary and secondary transconjugants, recipient gene characterization remained the same with the exception of vanA or vanB. The presence of plasmid DNA was not detected in any of the mating strains or transconjugants.
Phenotypically negative strains are shown in parentheses.
Rf, rifampin (20 μg/ml); Fd, nitrofurantoin (32 μg/ml); Te, tetracycline (10 μg/ml); Fs, fusidic acid (20 μg/ml); S, susceptible; R, resistant; phenotypes provided for only strains used in mating experiments.
Bacteriocin assay
None of the donor or recipient strains used in conjugation experiments tested positive for bacteriocin production (data not shown).
Mating experiments
Mating protocol was followed as per Fig. 1. Results of broth and filter matings are shown in Table 2. In primary mating experiments, transfer of vanA occurred between E. faecalis C2 to an E. faecium salad recipient (S4) by filter mating with an average conjugation frequency of 10−8 per recipient. Transconjugants were not observed among any of the other primary conjugative pairs either by broth or by filter. Upon secondary mating using the primary transconjugant (T2G) as the donor, transfer of vanA by the filter method was successful with each of the four recipient enterococci tested (S6, B12, CVM 3001, and CVM 3002) and by the broth method with two of the recipients (CVM 3001 and CVM 3002). Average frequency of transfer increased two- to four-log-fold, with an average frequency of transfer ranging from 10−4 to 10−6 per recipient. Antimicrobial resistance profiles for transconjugants varied from parent strains in both primary and secondary matings. In all cases, transconjugants showed an increase in number of resistant MIC values than compared with the donors and recipients (Table 3). In all cases, high-level resistance to vancomycin was maintained.
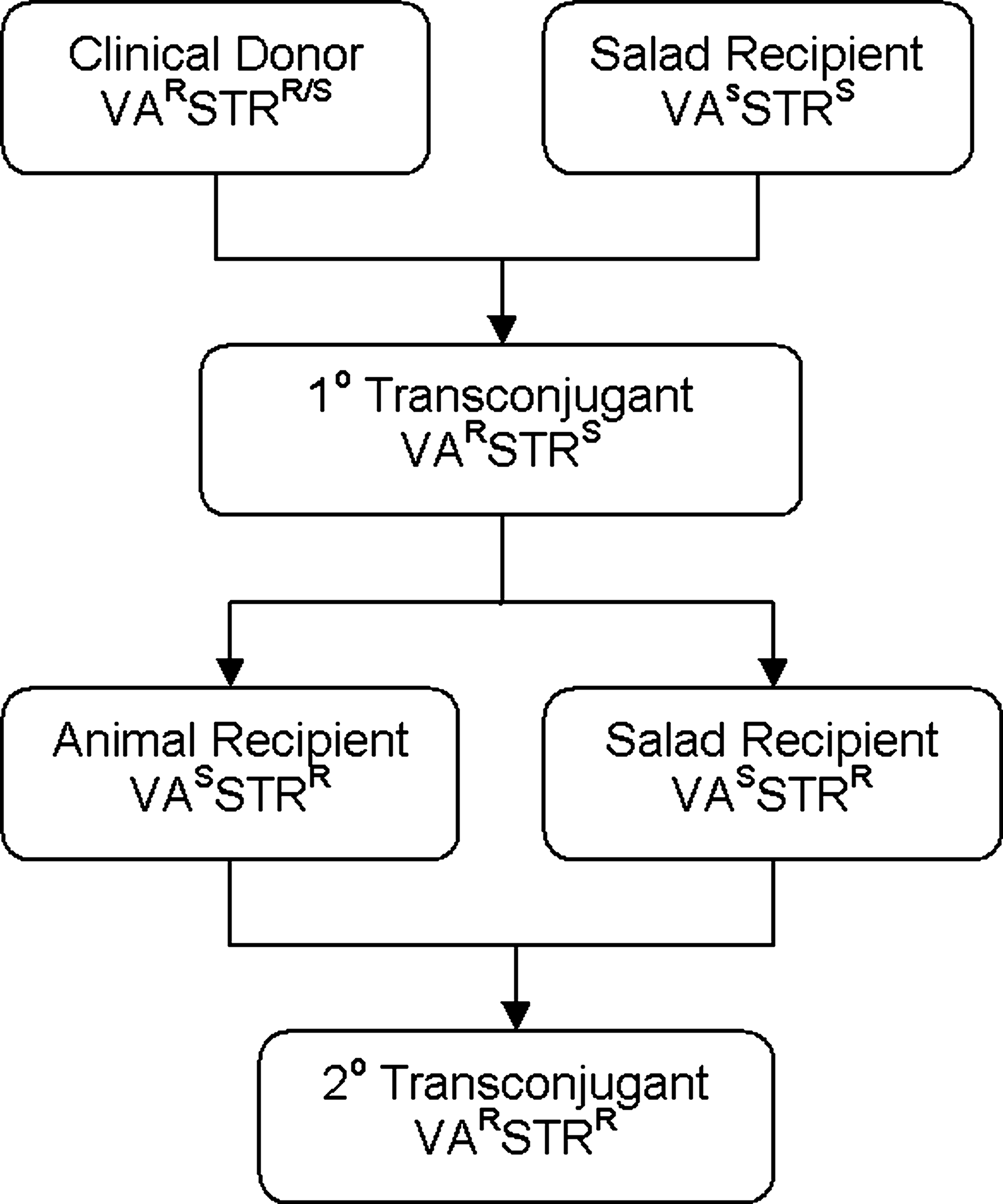
Schematic of mating protocol. VA, vancomycin; STR, streptogramin; S, susceptible; R, resistant.
Number of transconjugants per recipient.
NA, not applicable.
Minimal inhibitory concentration breakpoints in μg/ml based on values given in Clinical Laboratory Standards Institute6 for vancomycin (Va), ≥32; ampicillin (Am), ≥16; ciprofloxacin (Cp), ≥4; erythromycin (E), ≥8; levofloxacin (Lx), ≥8; linezolid (Lz), ≥8; penicillin (P), ≥16; rifampin (Rf), >2; synercid (Syn), ≥4; tetracycline (Te), ≥16; fs, E. faecalis; fm, E. faecium.
RNA isolation and RT-PCR
RT-PCR detected vanA RNA in E. faecium T2G grown in broth at 4°C, 10°C, 22°C, and 37°C with and without filtrate (Fig. 2). Additional temperatures were included for comparative purposes. No vanA RNA products were detected in E. faecalis C2. Results were similar for cells grown in salad (data not shown). Species-specific sodA mRNA was detected in isolates from all samples.
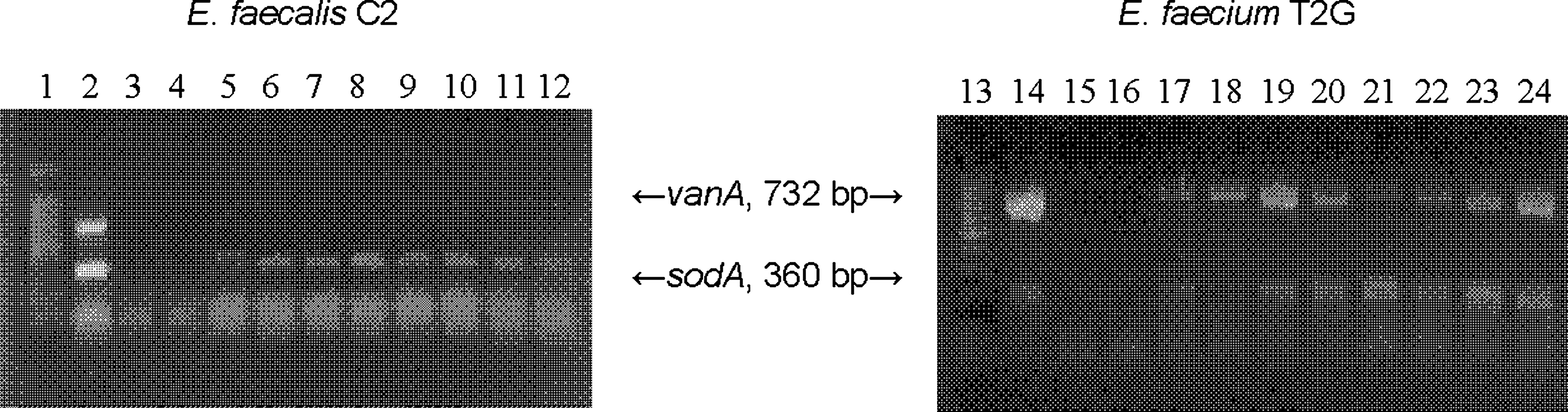
Reverse transcriptase–polymerase chain reaction results for vanA and sodA for Enterococcus faecalis C2 (donor) and Enterococcus faecium T2G (transconjugant) cultivated in brain heart infusion broth without antibiotics. Lane 1, molecular marker (100 bp); lane 2, E. faecium ATCC 51559 (positive polymerase chain reaction control for vanA and sodA); lanes 3/15, E. faecium S4 (negative control; no template); lanes 4/16, E. faecium S4 (negative control; no reverse transcriptase); lanes 5/17, 4°C without filtrate; lanes 6/18, 10°C without filtrate; 7/19, 22°C without filtrate; 8/20, 37°C without filtrate; 9/21, 4°C with filtrate; 10/22, 10°C with filtrate; 11/23, 22°C with filtrate; 12/24, 37°C with filtrate.
Discussion
The clinical, food, and animal strains used for primary mating showed a difference in the incidence of virulence determinants between E. faecalis and E. faecium consistent with previously published data. 10 With the exception of animal isolate B13, all E. faecalis recipients carried the selected virulence genes agg, cpd, and gelE, whereas efaA was detected in all E. faecium. There was no other distinguishing characteristic of B13 compared with the other E. faecalis isolates. Antimicrobial susceptibility patterns for the four clinical donor VRE were similar in that all were susceptible to nitrofurantoin and fusidic acid. All E. faecalis demonstrated characteristic intrinsic streptogramin resistance. Recipient enterococci were selected on the basis of antibiotic resistance patterns for transconjugant selection purposes; hence, all food and animal E. faecium were resistant to nitrofurantoin and/or rifampin. The standard recipient E. faecalis JH2-2 was included for mating experiments, and this was the only isolate in the study that demonstrated resistance to fusidic acid. In primary mating experiments, among 39 donor–recipient pairs, transconjugant generation was detected between one VanA E. faecalis donor, and an E. faecium salad recipient, S4, at an average conjugation frequency of 10−8 per recipient. No transconjugants were detected for any of the other mating pairs, indicating the inability to transfer under in vitro conditions. The relatively low rate of transfer in the absence of bacteriocin production may be due to the absence of selective antibiotic pressure or limited selection of recipient enterococci due to genotypic requirements for selective isolation. In a study of 40 VRE donors from retail poultry carcasses, Novais et al. reported intraspecies transfer of vancomycin resistance between only two mating pairs, an E. faecium donor to E. faecium GE1 and an E. faecalis donor to E. faecalis JH2-2, at a frequency of 10−7 and 10−8, respectively, attributing the low transfer frequency to adaptation of transferable elements to particular hosts. 24
The vanA gene is carried on transposon Tn1546 and can be transferred via a conjugative plasmid and dissemination of vanA via enterococcal probiotic or starter strains has been demonstrated in vitro.10,17,20 In this study, the transfer of vanA from T2G to CVM 3001 and CVM 3002 in the absence of aggregation substance reveals that SRE could readily acquire glycopeptide resistance. This may reflect the alternative mechanisms of horizontal transfer that confer broad host-range resistance. Among enterococci from environmental sources, transfer efficiency for the pheromone-associated plasmid pAD1 is reported to be 10−1 at 37°C in liquid broth cultures. 21 We were unable to detect transconjugants using E. faecalis JH2-2, a known pheromone-producing recipient, indicating that transfer may have been due to a pheromone-independent process.
Conservation of vanA or vanB determinants or differences in binding regions among transposon-like elements in the donors may also have accounted for the low frequency of conjugation. Guardabassi and Dalsgaard found interspecies transfer of vanA among enterococci of environmental origin to range from 10−3 to 10−7 per recipient by filter mating when Tn1546-like elements were structurally similar among enterococci of differing ecological origin. 14 In either case, multi-component salads that may contain commensal Enterococcus could serve as a medium for efficient transfer of vanA between donor and recipient bacteria particularly if clinical strains of resistant Enterococcus were to be introduced, such as in a hospital or nursing home kitchen. Natural acquisition of vancomycin resistance by non-enterococcal bacteria may occur via pheromone dependent or independent response mechanisms.11,29 Recently, an Inc 18-like plasmid conferring vanA was identified in vancomycin-resistant Staphylococcus aureus and VRE isolated from the same patient. 33 The transfer frequency of these broad host-range plasmids does not rely on pheromone responsiveness. In a food matrix that consists of both liquid and solid components as well as enterococci and commensal nonenterococcal bacteria from different sources, dissemination of high-level glycopeptides resistance may be facilitated. We investigated the transfer of vanA from the primary E. faecium transconjugant, T2G, to streptogramin-resistant salad and animal E. faecalis and to E. faecium CVM 3001 and CVM 3002 that carried vatE and vatD streptogramin resistance determinants. In secondary mating, transconjugants were detected by filter mating with all recipients with an increased conjugative frequency ranging from 10−4 to 10−7 per recipient, indicating that a native salad recipient originally carrying no glycopeptide or streptogramin resistance genes, or the selected conjugative virulence determinants agg, cpd, or gelE, could disseminate vanA to more virulent commensal SRE.
Among the primary and secondary transconjugants, high-level vancomycin resistance of ≥256 μg/ml was the same as with clinical donor strains. Antimicrobial susceptibility patterns overall resembled the recipient enterococci, with an increase in resistance to four or more additional antibiotics in the transconjugant. One notable pattern was the emergence of penicillin resistance in some transconjugants. In E. faecium, an increased resistance to β-lactam antibiotics can be conferred among E. faecium at low a frequency (10−10 to 10−7) by the VanB-encoding transposon Tn5382; however, the transfer of vanB was not observed in this study.2,15
In this study, the interspecies horizontal transfer of vanA between clinically derived and food-borne Enterococcus resulted in a transconjugant that demonstrated active vanA gene expression at 37°C. Detection of RNA products in transconjugant cultures incubated at 4°C, 10°C, and 22°C indicates that vanA expression could occur over a range of temperatures and food storage conditions. The presence or absence of vanA transcription products differed between the donor and the transconjugant, but was not dependent on temperature or the presence of pheromone that could be present in the S4 filtrate. All cells were cultivated without exposure to vancomycin; therefore, no detection of vanA RT-PCR products in RNA preparations from E. faecalis C2 was most likely due to lack of an inducing agent. However, in transconjugant cells, vanA mRNA was detected in the presence and absence of filtrate regardless of temperature. Thus, the clinical VRE did not express vanA constitutively, whereas expression did occur in transconjugants grown in the absence of vancomycin. Since growth conditions for RNA isolation were the same for both strains, so it is possible that lack of induction was due to accumulated depsipeptide peptidoglycan precursors in donor cells. Alternatively, since the transconjugant resulted from interspecies mating, inherent responses to environmental or biological cues could account for differences in gene expression. 28 Each of the E. faecium recipients that acquired vanA maintained high level resistance to >2 μg/ml of Q/D and ≥16 μg/ml of virginiamycin.
The prevalence of streptogramin-resistant genes, including vatD and vatE, among Q/D-resistant E. faecium isolated from foods, animals, and humans in the United States is low, suggesting an undetermined mechanism for transfer of streptogramin resistance still exists. 9 The mating protocol depicted in Fig. 1 was designed as a model for in situ transformation of commensal bacteria in a mixed-component food system. Our data show that an increase in secondary horizontal transfer of vanA occurs among Enterococcus from prepared deli salad at 37°C. The increase in transfer efficiency upon secondary mating can lead to enhanced resistant profiles of commensal Enterococcus recipients. Introduction of Enterococcus from preparation and/or handling of these types of food matrices may result in the development of multidrug resistant populations of commensal bacteria upon secondary horizontal transfer. This study also demonstrates that in transconjugant commensal E. faecium isolated from prepared salad, expression of vanA can be induced in the absence of selective antibiotic pressure.
Footnotes
Acknowledgments
We thank Dr. David White of the Center for Veterinary Medicine (USFDA) for providing the E. faecium CVM 3001 and CVM 3002 and E. faecalis JH2-2 strains used in the study, and Dr. Melvin Weinstein of Robert Wood Johnson University Hospital for facilitating the acquisition of clinical Enterococcus isolates. E.A.C. would like to thank Dr. Hoan-Jen Pang for useful discussions regarding multiplex PCR and filter mating.
Disclosure Statement
No competing financial interests exist.