
Abstract
The study aimed to analyze drug resistance and mutations and genetic evolution of influenza A and influenza B viruses during the 2013 and 2014 influenza season in Beijing, China. RNA was extracted from pharyngeal or nasal swabs of 28 patients, and determination of influenza genotypes was performed by using real-time reverse-transcription polymerase chain reaction. Influenza A virus samples were sequenced with the neuraminidase (NA) gene and M2 matrix protein gene to determine the NA inhibitor (NAI) resistance and amantadine resistance mutations, and influenza B virus samples were sequenced with the NA gene and hemagglutinin (HA) gene to analyze NAI resistance mutations. As a result, the enrolled subjects consisted of 19 patients with the A(H1N1)pdm09 subtype, four with A(H3N2) subtype and five with influenza B virus. All of the 23 samples with influenza A viruses harbored amantadine resistance mutation S31N in M2 matrix protein. V241I, a compensatory NAI resistance mutation, was detected in all of the 19 A(H1N1)pdm09 viruses. No other NAI resistance mutation was observed in both influenza A and B viruses. The NA gene of the five influenza B virus strains was classified as B-Victoria lineage, while the HA gene of five strains was classified as B-Yamagata lineage. In summary, all influenza A viruses from patients in Beijing in the 2013–2014 season were resistant to amantadine agent. Both influenza A and B viruses kept sensitive to NAIs. Lineage recombination was detected in influenza B virus strains and may impair the efficacy of influenza vaccination.
Introduction
T
Currently, there have been two classes of antiviral drugs available for treating influenza A virus infection. They are M2 ion channel blocker adamantanes (amantadine and rimantadine) and NA inhibitors (NAIs, including oseltamivir, zanamivir, peramivir and laninamivir). Oseltamivir has been the most commonly used medication in clinical practice since its approval in 1999. However, the emergence and spread of antiviral resistance viruses pose a challenge to the treatment of influenza virus. Amantadine resistance mutations include L26F, V27A, A30T, S31N, and G34E, among which S31N is the most frequently detected mutation in circulating influenza type A viruses.7,8 Nearly all influenza A viruses circulating were resistant to amantadine since 2009. 9 Compared with amantadine resistance mutant, the oseltamivir resistance mutant was detected at a lower rate but spread faster. 10 The most common NAI resistance mutations are limited to H275Y and N295S for the N1 subtype, E119V and R292K for N2 subtype viruses, and D197N/E/Y and I221T/V for the B type.11,12
The prevalence of NAI drug resistance and epidemic strain has been changing along with time. In the 2008–2009 season, almost 100% of the seasonal H1N1 viruses were H275Y mutants with oseltamivir resistance in Japan and other countries. 5 Oseltamivir-resistant seasonal H1N1 virus was not detected in the 2009–2010 season around the world, and the A(H1N1)pdm09 strain became prevalent and was sensitive to oseltamivir; in 2011–2014, the dominant subtype turned to be A(H3N2) and influenza B. 13 These reports highlighted the importance of close monitoring of evolutionary traits and antiviral susceptibility profile of influenza viruses. However, data were limited in drug resistance and genetic evolution of influenza A and B viruses in Beijing during 2013 and 2014.
In the present study, we aimed to investigate (1) the prevalence of influenza type A and type B viruses to amantadines and NAIs during the 2013 and 2014 influenza season in Beijing, China and (2) the molecular epidemiology and phylogenetic characteristics of influenza viruses.
Materials and Methods
Sample collection
A total of 28 patients who visited Beijing 302 Hospital in Beijing city were included in this study. The inclusion criteria for enrollment were fever (≥38°C) plus either cough or sore throat within the past 72 hours before patient presentation. The swabs were transported immediately in virus transport media tubes and then frozen at −80°C for further analysis. This study was conducted with the approval of the Ethics Committees of Beijing 302 Hospital.
RNA extraction and RT-PCR for detection of influenza A and B virus
RNA was extracted from 200 μl of preservation solution for swab samples, using the QIAamp MinElute Virus Spin Kit (Qiagen) according to the protocol. The type of influenza viruses was determined by real-time reverse-transcription quantitative PCR (Roche).
Amplification and sequencing of NA and M2 matrix protein gene
Reverse transcription of influenza A virus samples was performed by using Invitrogen SuperScript® III one-step RT-PCR with Platinum Taq Kit (Invitrogen). Upstream and downstream universal primers of influenza A virus were MBT-uni12 and MBT-uni13. 14 N1- and N2-specific primers were used to subtype influenza A viruses. The M2 matrix protein gene was amplified and sequenced by M2U12 and M2D12. For identifying the influenza B virus, reverse transcription was performed by using Invitrogen SuperScript® III First-Strand Synthesis System for RT-PCR kit (Invitrogen). NA and HA genes of influenza B virus were sequenced by two-round nested PCR. Primers used in this study are shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr).
Drug resistance mutation analysis
The incidences of NAI and amantadine resistance mutation were analyzed as previously described.5,12,15 Primary NAI resistance mutations can reduce the susceptibility of influenza virus to NAIs, including D151E, Y155H, D199N, I223K/R, H275Y, and N295S for oseltamivir resistance of A(H1N1) virus, P120L,Q136K/R, D151E, Y155H, N221D, and I223R for zanamivir resistance of A(H1N1) virus, D151E and N295S for peramivir resistance of A(H1N1) virus; E119V/I, D151G, R292K, and N294S for oseltamivir resistance of A(H3N2) virus, E119I, Q136K/R, D151G, and R292K for zanamivir resistance of A(H3N2) virus, R292K for peramivir resistance of A(H3N2) virus; R150K, D197N/E/Y, I221V/T, and R374K for oseltamivir resistance and zanamivir resistance of influenza B virus, R150K and D197N for peramivir resistance of influenza B virus. Secondary mutations or compensatory mutations do not confer resistance to NAIs directly but compensate for the viral fitness, The V241I and N369K substitutions in the NA of the H275Y mutant A(H1N1)pdm09 viruses were reported to increase its replication and transmission fitness.16,17 Adamantane-resistant mutations include L26F, V27A, A30T, S31N, and G34E for A(H1N1) virus and L26F, V27A, A30T, S31N, and G34E for A(H3N2) virus.
Evolutionary analysis of influenza A and B viruses
Evolutionary analysis of NA gene of influenza A virus was carried out by including sequences announced as the Northern Hemisphere influenza vaccine strains recommended in the 2013–2014 season by the WHO, that is, A/Christchurch/16/2010 and A/Texas/50/2012. All reference sequences were retrieved from NCBI Influenza Virus Resource database.
Evolutionary analysis of NA and HA gene of influenza B virus was performed by including WHO-recommended vaccine strains B/Massachusetts/02/2012 for B/Yamagata lineage and B/Brisbane/33/2008 for B/Victoria lineage. Phylogenetic analysis was performed with the Neighbor-Joining method and Kimura 2-Parameter and by the procedure of Bootstrap 1,000 replicates in the software package Mega 6.0 software.
Results
Type/subtype classification
Three types/subtypes were determined across the study samples. The GenBank accession numbers for 19 A(H1N1) subtype virus sequences and five influenza B virus sequences in this study are listed in Tables 1 and 2, respectively. In addition, there were 4 A(H3N2) virus sequences. The accession numbers of these sequences are KR025189 (A/Beijing/FL29/2013), KR025190 (A/Beijing/FL31/2013), KR025191 (A/Beijing/FL34/2013), and KR025193 (A/Beijing/FL93/2013).
Primary NAI resistance mutations: D151E, H275Y, N295S; Compensatory NAI resistance mutations: V241I.
NAIs, neuraminidase inhibitors.
Adamantane and NAI resistance mutations
In this study, 28 patients with influenza virus infection consisted of 19 patients with A(H1N1)pdm09, 4 with influenza A(H3N2) and 5 with influenza B virus.
For amantadine resistance mutation, S31N was detected in all 23 patients with influenza A virus infection, but other amantadine resistance mutations, that is, L26F, V27A, A30T, and G34E, were not detected.
For A(H1N1)pdm09 subtype, no mutation that confers NAI resistance was detected. However, 19 patients were detected with V241I mutation, which was previously identified as compensatory mutations (Table 1). No NAI resistance-associated mutations were detected in A(H3N2) subtype and influenza B virus.
Phylogenetic analyses of NA and HA gene in different types/subtypes of influenza virus
Phylogenetic analysis showed that the average genetic similarity of NA gene in A(H1N1)pdm09 strains between tested strains and the vaccine strain A/Christchurch/16/2010 was 99.02%. The NA gene of A(H1N1)pdm09 strains in Beijing from 2013 to 2014 was allocated to respective clades year by year (Fig. 1).
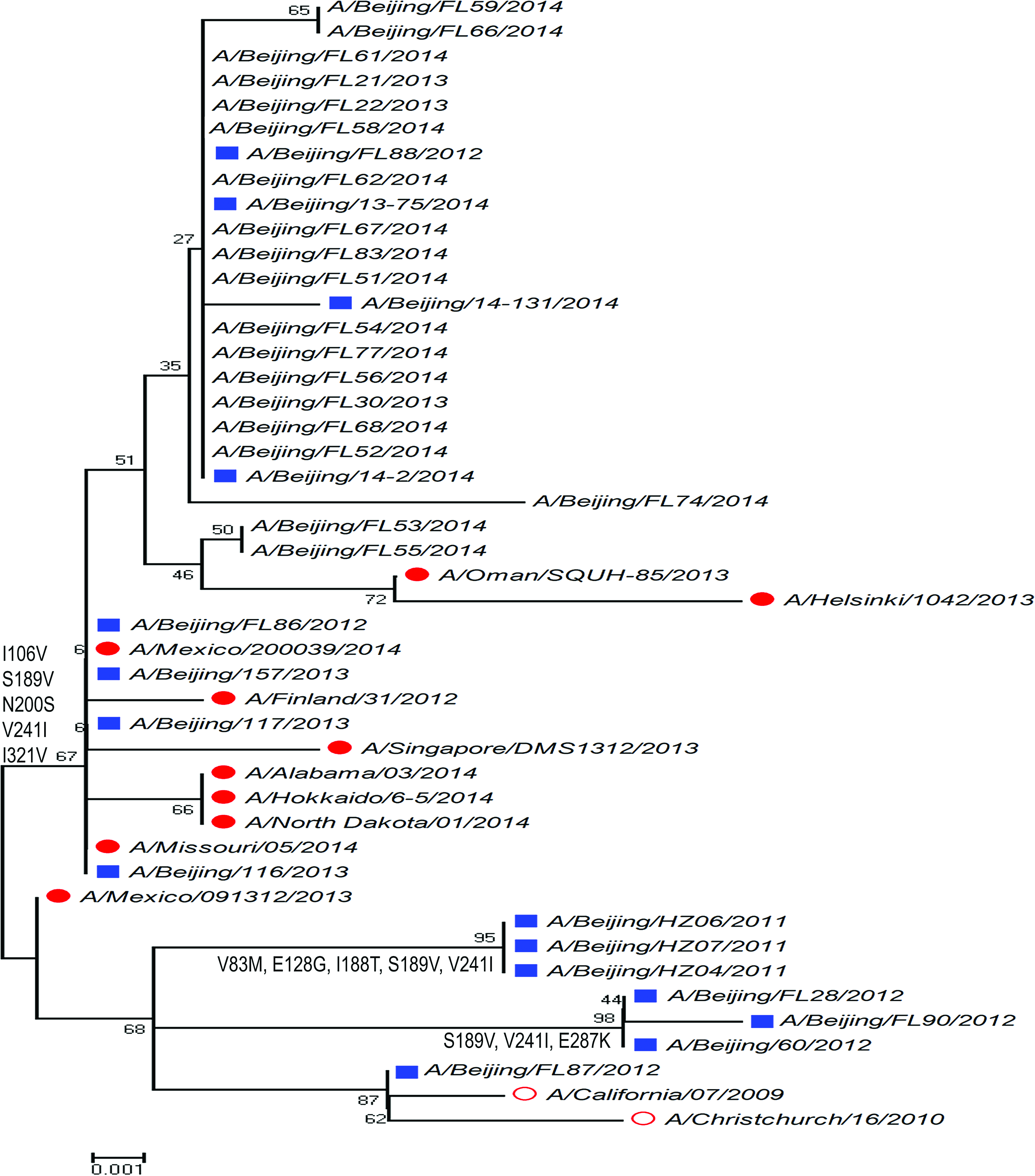
Phylogenetic analysis of H1N1 NA gene from Beijing during the 2013 and 2014 influenza season. Red circle: vaccine candidate strains; solid red circle: strains circulating out of China; blue square: strains circulating in China; normal character: strains isolated from our laboratory. NA, neuraminidase. Color images available online at www.liebertpub.com/mdr
For NA gene of A(H3N2) subtype (Fig. 2), the average genetic similarity between tested strains and the vaccine strain A/Texas/50/2012 was 98.98%. The year-by-year evolutionary trait of A(H3N2) subtype was similar with A(H1N1)pdm09 but less significant. For example, the A/Beijing/FL93/2013 strain was allocated with the strain (A/Beijing/346/2009) circulating in the 2009–2010 influenza season at one evolutionary branch rather than the strain (A/Hangzhou/B754/2013) circulating in the 2013–2014 influenza season.

Phylogenetic analysis of H3N2 NA gene from Beijing during the 2013 and 2014 influenza season. Red circle: WHO-recommended vaccine candidate strains; solid red circle: strains circulating out of China; blue square: strains circulating in China; normal character: strains isolated from our laboratory. WHO, World Health Organization. Color images available online at www.liebertpub.com/mdr
For influenza B virus, phylogenetic analysis of NA gene (Fig. 3) showed that all of the five samples were distributed to the B/Brisbane/33/2008 (B-Victoria lineage) rather than the WHO-recommended vaccine strain B/Massachusetts/2/2012 (B-Yamagata lineage). By contrast, phylogenetic analysis of HA gene (Fig. 4) showed that five samples were distributed to the B-Yamagata lineage.
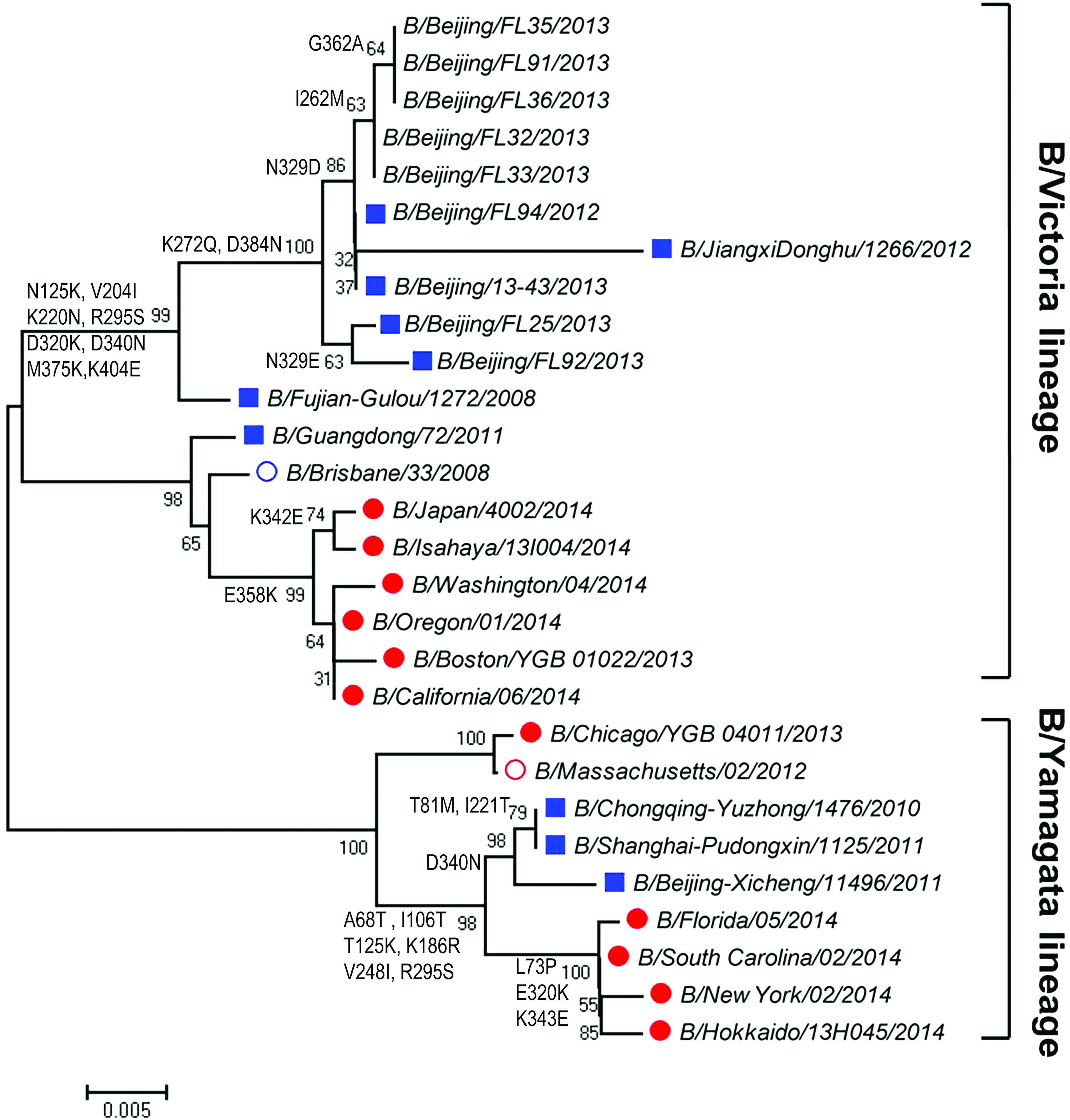
Phylogenetic analysis of NA gene of influenza B virus from Beijing during the 2013 and 2014 influenza season. Red circle: WHO-recommended trivalent vaccine candidate strains; blue circle: WHO-recommended quadrivalent vaccine candidate strains; solid red circle: strains circulating out of China; blue square: strains circulating in China; normal character: strains isolated from our laboratory. Color images available online at www.liebertpub.com/mdr
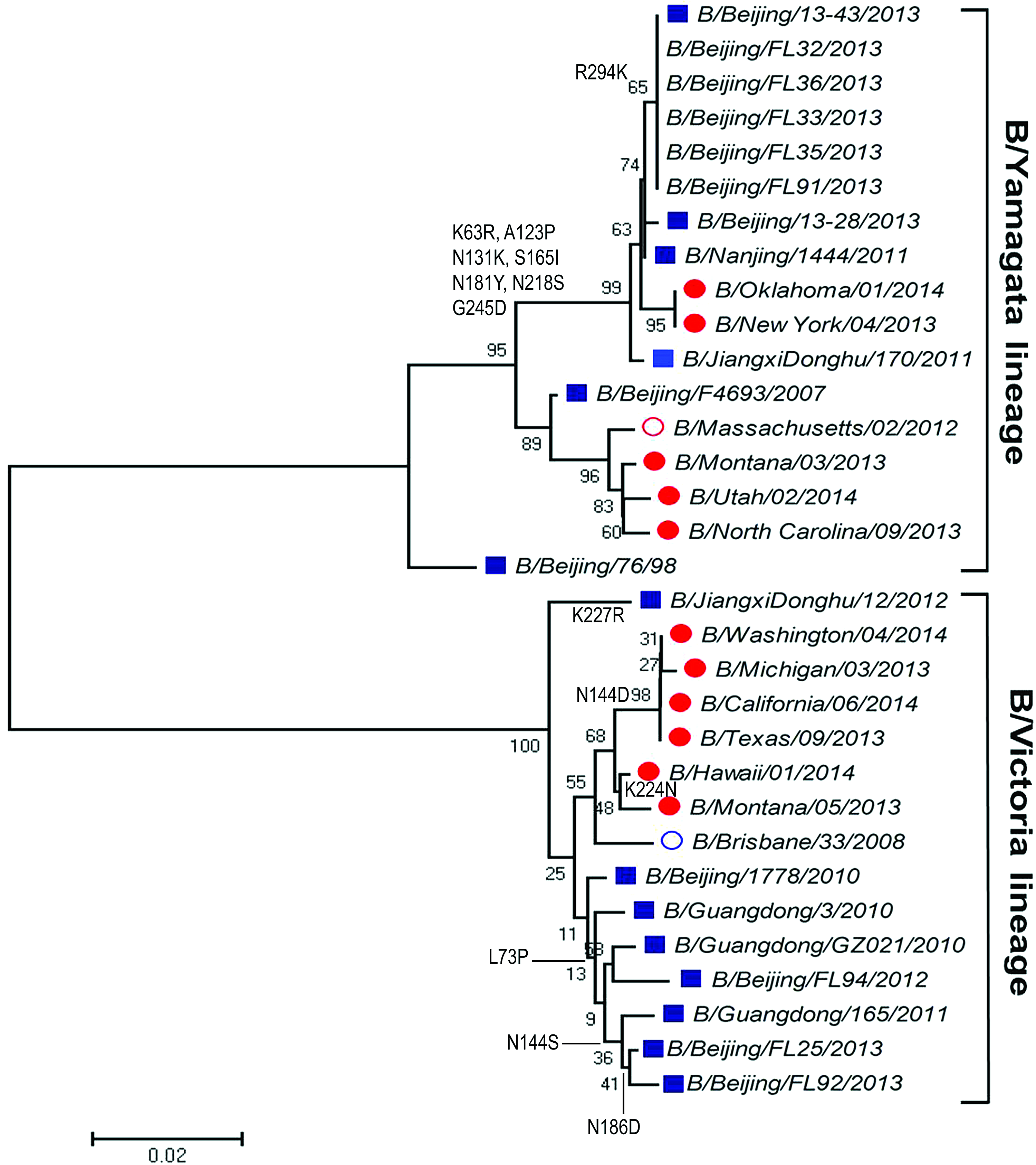
Phylogenetic analysis of hemagglutin in gene of influenza B virus from Beijing during the 2013 and 2014 influenza season. Red circle: WHO-recommended trivalent vaccine candidate strains; blue circle: WHO-recommended quadrivalent vaccine candidate strains; solid red circle: strains circulating out of China; blue square: strains circulating in China; normal character: strains isolated from our laboratory. Color images available online at www.liebertpub.com/mdr
In addition, Table 2 lists the amino acid mutations in the HA sequence of influenza B virus strains compared with the 2013–2014 vaccine strains. There were eight amino acid changes (K63R, A123P, N131k, S165I, N181Y, N218S, G245D, and R294K) that presented in all five Beijing strains compared with the 2013–2014 vaccine strain, B/Massachusetts/02/2012.
Discussion
Influenza virus infection is a serious public health issue worldwide. Antiviral drugs are an important supplement to vaccination for reducing the public health impact of influenza virus infections. To select the optimal NAIs for clinical use, it is important to monitor the resistance of different influenza virus strains. 18 Adamantanes and NAIs represent two classes of drugs that target the viral matrix protein 2 and NA, respectively. The widespread use of adamantanes and NAIs against influenza has led to the emergence of resistant viral strains. 19 In this study, resistance monitoring showed that all strains with influenza A(H1N1)pdm09 and A(H3N2) subtype were resistant to adamantanes during 2013–2014 in Beijing, consistent with the results of the Chinese National Influenza Center. 20 In fact, by 2012, all circulating A(H1N1)pdm09 and A(H3N2) viruses were naturally resistant to the M2 ion-channel blockers, so priority should be given to testing for NAI susceptibility. 21 The only amino acid substitution in the NA protein with resistance to NAIs in clinical cases, which has been demonstrated in large population studies, is the H275Y substitution.22,23 In the 2013–2014 influenza season, a community cluster of oseltamivir and peramivir cross-resistance viruses occurred in Japan 24 and significant numbers of these resistant viruses were detected in China 19 and the United States 25 In this study, no H275Y substitution was detected, which indicates that NAIs can still serve as the first-line antiviral drugs in China. However, almost all A(H1N1)pdm09 subtype strains were detected with V241I substitution, which were identified to be compensatory mutation for virus fitness in an in vitro experiment. 16 Therefore, intense surveillance of NAI resistance mutations is still needed.
Beijing as a quickly expanding metropolis has a great number of immigrants coming from other regions of China and other countries around the world. This may complicate the evolutionary traits of influenza virus in this region, because influenza viruses undergo mutations when they spread from place to place and therefore introduce gradual changes in the HA and/or NA gene. Therefore, it is critical to get more knowledge about the molecular evolution of these viruses for producing effective vaccines. For influenza A virus, A(H1N1)pdm09 was the predominant subtype in the 2013–2014 influenza season, whereas for influenza B virus, the B-Yamagata lineage was the predominant lineage and the B-Victoria lineage accounting for a small proportion of cases in the world. Our results showed that A(H1N1)pdm09 and A(H3N2) influenza viruses in the Beijing area have same clades with relevant vaccine strains A/Christchurch/16/2010 [A(H1N1)pdm09] and A/Texas/50/2012 (H3N2) on the evolutionary branches, respectively. Notably, the influenza B virus based on the phylogenetic tree of NA and HA classification was inconsistent. All of the five influenza B virus genomes based on NA genotyping belonged to B-Victoria lineage, but HA genomes belonged to B-Yamagata lineage. The reassortment should be taken into serious consideration for monitoring future influenza outbreaks in the Beijing region.
There are four major antigenic epitopes presenting on the membrane distal domain of HA of influenza B virus that lie in the 120-loop (amino acid [aa 116–137]), 150-loop (aa 141–150), and 160-loop (aa 162–167), and in the 190-helix (aa 194–202). 26 On comparison with the vaccine strain B/Massachusetts/02/2012, A123P and N131K substitutions were identified in the 120 loop of the HA1 region in all the Beijing strains; S165, which is located in the 160 loop, was replaced by I and was detected in all the Beijing strains in our study. This might have an impact on the antigenicity of HA protein and suggested that regular influenza vaccination might not confer all-round protection.
In summary, NAI resistance mutation in influenza viruses was relative low during this period of 2013–2014 in the Beijing area. The reassortant strain, which combined NA gene of B-Victoria lineage and HA gene of B-Yamagata lineage together, may reduce the efficacy of influenza vaccine based on the WHO-recommended vaccine strain. Therefore, the quadrivalent vaccine, which can provide protection against both the B-Victoria and B-Yamagata lineage, is strongly recommended for future vaccine production in China.
Footnotes
Acknowledgments
This work was supported by the PLA 12th Five-Year Grand Project for Key Techniques and Devices in Management of Infectious Diseases (BWS11J048), and the Science and Technology Project of Beijing, China (No. D131100005313016).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.