
Abstract
Difference in the development of resistance may be associated with the epidemiological spread and drug resistance of different Salmonella enterica serovar strains. In the present study, three susceptible S. enterica serovars, Typhimurium (ST), Enteritidis (SE), and Indiana (SI) strains, were subjected to stepwise selection with increasing ciprofloxacin concentrations. The results indicated that the mutation frequencies of the SI group were 101–104 higher and developed resistance to ciprofloxacin more rapidly compared with the ST and SE groups. Ciprofloxacin accumulation in the SI strain was also higher than the other two strains in the presence of an efflux pump inhibitor. The development of ciprofloxacin resistance was quite different among the three serovar strains. In SI, increasing AcrAB-TolC efflux pump expression and single or double mutations in gyrA with or without a single parC mutation (T57S) were found in the development of ciprofloxacin resistance. In SE, an increase in the AcrAB-TolC efflux pump regulatory gene ramA gradually decreased as resistant bacteria developed; then resistance resulted from gyrA D87G and gyrB E466D mutations and/or in other active efflux pumps besides AcrAB-TolC. For ST, ramA expression increased rapidly along with gyrA D87 N and/or gyrB S464F mutations. In conclusion, persistent use of ciprofloxacin may aggravate the resistance of different S. enterica serovars and prudent use of the fluoroquinolones is needed. The quicker resistance and higher mutation frequency of the SI isolates present a potential public health threat.
Introduction
S
Quinolone resistance is often associated with chromosomal mutations, and the most common of these are the mutations in the quinolone resistance-determining regions (QRDRs) of the target genes (gyrA; gyrB; parC; and parE). On the other hand, quinolone resistance has also been attributed to a reduced intracellular accumulation of drugs through overexpression of active efflux pumps (mainly AcrAB-TolC). In Salmonella, the reported AcrAB-TolC efflux pump regulators are RamA, SoxS, MarA, Rob, and AcrR, all of which have proved to be associated with quinolone resistance by regulating the expression of AcrAB-TolC efflux pump. 4 RamA, which can be inhibited by RamR, is a global regulator and plays a major role in upregulating AcrAB-TolC, 5 and the previous study also suggested that the expression of RamA can promote the development of ciprofloxacin resistance. 6 Mutations or deletions in the RamR binding site found upstream of RamA can increase the expression of RamA and AcrAB-TolC efflux pump. 5
Serotyping is very useful that allows for trends in Salmonella surveillance data to be followed with the passage of time and over 2,600 serotypes of Salmonella have been identified. Surveys have shown that Salmonella enterica serovar Enteritidis is the most prevalent serotype, followed by S. enterica serovars Typhimurium and Infantis in the USA and Europe.7,8 In China, serotype distribution varies between provinces, but in general, the most prevalent serovars are Typhimurium and Enteritidis with a rising rate of reports of S. enterica serovar Indiana.3,9,10 S. Indiana isolates have been rarely reported previously. There are few reports of large-scale food poisoning cases by S. Indiana isolates in the USA and in Europe.11,12 However, the Indiana serotype is becoming more prevalent in China at veterinary clinics and in animal foods; these numbers have recently significantly increased to 42.8% and 45.7% in Shandong province13,14 and 67.7% in Guangdong province. 15 The characteristics of multidrug-resistant S. Indiana isolates have also been documented.3,14
Despite belonging to the same species, different S. enterica serotypes can vary in their ability to adapt to the environment. This is reflected in disease severity and clinical manifestations of illness as well as antibiotic resistance profiles. Indeed, the prevalence of these different serovar strains can also change over time. Monitoring data shows that the occurrence of quinolone resistance mechanisms is associated with a particular Salmonella serovar. 8 Therefore, a successful epidemiological spread of a drug resistance of Salmonella may be associated to specific serovars.16–18
The development of antibiotic resistance is an evolutionary inevitability which leads to increases in morbidity, mortality, and healthcare costs. Previous study showed that there was temporal interplay between efflux pumps and target mutations in the development of antibiotic resistance in Escherichia coli. 19 However, there are few studies concerning the development of antibiotic resistance in different S. enterica serovars. Whether there is a particular mechanism associated with the rapid evolution of S. enterica resistance is unknown.
This study was designed to gain an understanding of the drug resistance development process of three different S. enterica serovar strains (Typhimurium, Enteritidis, and Indiana) from food animals. We wanted to determine whether different fluoroquinolone resistance mechanisms are involved by artificially generating mutants from susceptible strains in the presence of increasing ciprofloxacin concentrations in vitro. We monitored mutation frequencies and AcrAB-TolC efflux pumps expression and mutation in target genes and in RamR-RamA as well as following cellular ciprofloxacin accumulation.
Materials and Methods
Bacterial strains and antimicrobial susceptibility
Three different ciprofloxacin (minimum inhibitory concentration [MIC] 0.03–0.125 μg/ml) and nalidixic acid (MIC 8 or 16 μg/ml)-susceptible serotypes of S. enterica strains, Indiana (SI) (isolated from pig), Enteritidis (SE) (isolated from chicken), and Typhimurium (ST) (isolated from chicken), were used for this study. These three strains were analyzed for mutations and none was found in the QRDRs of gyrA, gyrB, and parE, but a mutation in parC (T57S) was identified in SE and ST isolates. Growth curves of the three isolates showed no significant difference compared with that of S. enterica Typhimurium ATCC14028.
Susceptibility testing was determined for these three parental isolates using the standard agar dilution method on Mueller-Hinton (MH) (Huankai Microbial, Guangzhou, China) agar according to the Clinical and Laboratory Standards Institute (CLSI) guidelines. 20 The following antimicrobials were tested: ciprofloxacin (CIP), nalidixic acid (NAL), enrofloxacin (ENR), levofloxacin (LEV), ampicillin (AMP), tetracycline (TET), chloramphenicol (CHL), and kanamycin (KAN) (National Institutes for Food and Drug Control, Beijing, China). E. coli strain ATCC 25922 was used as the control (maintained in our laboratory).
Selection of ciprofloxacin-resistant mutants in vitro
Serial mutants of SI, SE, and ST were obtained in a multistep selection process with increasing ciprofloxacin pressure as previously described with some modifications. 21 Briefly, the three parental strains were grown in MH broth at 37°C overnight. One hundred microliters of the overnight culture was spread on MH plates supplemented with half of ciprofloxacin MIC concentration of each parental strain and incubated for 24 h at 37°C. Single colonies were selected and passaged 5 times in MH broth without ciprofloxacin, and then MIC value of ciprofloxacin was determined. One of the selected single colonies was retained for the next step and was incubated in MH broth containing the same concentration of ciprofloxacin as that in the plate for 24 h at 37°C. One hundred microliters of the overnight culture was spread onto MH plates supplemented with an increased concentration of ciprofloxacin. This procedure was then repeated until mutants exhibiting high MICs for ciprofloxacin were identified. Resistant strains that grew in the presence of ciprofloxacin with the stable phenotype (not adaptive growth) after being passaged 5 times were defined as serial mutants. All SI, SE, and ST mutants were grouped according to MICs as follows: decreased susceptible (MIC 0.5–2 μg/ml), resistant (MIC 4–8 μg/ml), and high-level resistant (MIC ≥16 μg/ml), according to CLSI guidelines. Serial mutants were designated as M1, M2, M3, and M4 and were further studied. MIC values of NAL, ENR, LEV, AMP, TET, CHL, and KAN were also examined as above and the selected mutants were stored at −80°C until used.
Mutation frequencies
The mutation frequencies were calculated as the number of detectable colonies in MH agar with ciprofloxacin divided by the number of the ones in MH agar without ciprofloxacin. 22
DNA sequence analysis of gyrA, gyrB, parC, parE, and ramR-ramA
Mutations in the QRDRs of gyrA, gyrB, parC, and parE in mutants and ramR-ramA intersection in both the parent strains and mutants were detected using the primers described in previous studies. All PCR products were directly sequenced to confirm the mutations as previously described.5,23
Complementation testing was carried out using the parent strains if ramR-ramA mutations were detected. Wild-type ramR was amplified as previously described 5 and the PCR fragment was cloned into pGEM-T Easy Vector (Promega, Madison). The pGEM-T-ramR plasmid was then introduced into parent strains which had ramR mutations. The MICs of these strains was examined as above.
Relative expression levels of multidrug resistance efflux pumps
Total RNA was harvested using the RNAiso Plus Kit and the synthesis of cDNA was performed using the PrimeScript™ RT Reagent Kit with gDNA Erase (TaKaRa, Japan) according to the manufacturer's instructions. Real-time PCR was performed using the iQ5 multicolor real-time PCR system (Bio-Rad, Hercules) with specific primers (Table 1), cDNA template, and iQ SYBR Green Supermix (TaKaRa, Japan). The16S rRNA gene was used as an internal reference control. Each sample was run three times independently. Relative expression level of each gene was calculated using the 2−ΔΔCT method. 24
F, forward; R, reverse
Ciprofloxacin accumulation
Ciprofloxacin uptake was assayed by the method previously described with some modifications. 25 The fluorescence of the supernatant was measured with a Welch Ultimate Polar-RP Ci8 at excitation and emission wavelengths of 280 and 450 nm, respectively. The concentration of ciprofloxacin in the supernatant was calculated by comparison with a standard curve for ciprofloxacin (0.02–2.5 μg/ml) in 0.1 M glycine hydrochloride (pH 3.0). The results were expressed as nanograms of ciprofloxacin incorporated per milligram (dry weight) of bacteria. The experiments were performed three times to ensure reproducibility.
Results
Resistant phenotype and selection of ciprofloxacin-resistant mutants
The three parental strains SI, SE, and ST were susceptible to all tested antimicrobials (CIP, NAL, ENR, LEV, AMP, TET, CHL, and KAN). We then subjected these strains to ciprofloxacin pressure, a series of mutants were obtained from each parental strain and was identified based on MICs. The CIP concentrations used for M1, M2, M3, and M4 selection are given in Table 2. Four of each serotype mutants were used for further study. Overall, the mutants showed reduced susceptibility or resistance to other quinolones besides CIP, including ENR, LEV, and NAL. These mutants also showed reduced susceptibility or resistance to the unrelated antimicrobials such as KAN, CHL, TET, and AMP. Mutants were all resistant to AMP and TET, and 11/12 of these selected mutants were also CHL resistant (Table 2).
The results of MICs and chromosomal mutations of parental strains are shown in bold type.
M1, M2, M3, M4, serial selected mutants; — Wild-type allele (no mutation).
CIP, ciprofloxacin; NAL, nalidixic acid; ENR, enrofloxacin; LEV, levofloxacin; CHL, chloramphenicol; AMP, ampicillin; KAN, kanamycin; TET, tetracycline; MIC, minimum inhibitory concentration.
Interestingly, the processes of changing susceptibility among the three serotype strains were quite different; higher ciprofloxacin pressures were needed for SE (8–533 × parental MIC) and ST (8–2,133 × parental MIC) to increase the resistance level than that in SI (2–64 × parental MIC). When the MIC for a mutant was 0.5 μg/ml, the CIP selective pressure needed in SI was 0.25 μg/ml or twice the parental MIC. In SE and ST, selective pressure needed were 0.5 and 0.25 μg/ml, and these values were 8× the parental MIC values. When the parental strains were subjected to 64× MIC (CIP selective pressure were 8, 4, and 2 μg/ml in SI, SE, and ST, respectively), the mutant MIC values were 32, 4, and 2 μg/ml for SI, SE, and ST, respectively. These values were 256-, 64-, and 64-fold higher than MIC of corresponding parental strain. A similar case was seen in the other mutants (Table 2).
The frequencies of mutants also varied in SI, SE, and ST. SI developed resistance to ciprofloxacin at higher frequencies than that in SE and ST (Table 2). When the MIC for the mutant was 0.5 μg/ml, the mutation frequency of SI was 100-fold higher compared with SE and ST (10−7vs. 10−9). When high-level resistance was reached (MIC ≥16 μg/ml), SI had a 10–10,000 higher frequency and developed mutants more readily. It is worth noting that the frequencies of mutants reached as high as 4.28 × 10−3 for the SI-M4. A longer time was needed to select resistant mutants of SE and ST. These also grew as small colonies with changed morphologies (data not shown).
Identification of chromosomal mutations
The SI, SE, and ST parental strains had no mutations in gyrA, gyrB, and parE, but a mutation in parC (T57S) was detected in the SE and ST as mentioned above. After in vitro resistance development, these three serovars displayed quite different chromosomal mutations. For the SI group, a single gyrA mutation (S83F) was first detected in SI-M1 (MIC = 0.5 μg/ml). To develop high-level resistance (SI-M3) with MIC values 16 μg/ml, an additional mutation in gyrA (D87G) was detected. A parC T57S mutation in SI-M4 was also detected (Table 2). For the SE group, mutation in gyrA (D87G) was first detected in SE-M1 (MIC = 0.5 μg/ml) and an additional mutation in gyrB (E466D) was first present in further resistance strain SE-M2 (MIC = 2 μg/ml). The ST group was similar to the SE group with a few interesting exceptions. A single gyrA mutation (D87N) was first detected in ST-M1 (MIC = 0.5 μg/ml). High-level resistance with an MIC >16 μg/ml required an additional mutation in gyrB (S464F) (Table 2). However, the type of amino acid substitution in the gyrA Asp87 codon varied with the serovars. Mutations in the Asp87 codon resulted in substitutions to asparagine residues (D87N) in ST, while this encoded a glycine residue (D87G) in SI and SE. All mutants of SE and ST also retained the parC (T57S) mutations, but no parE mutation was observed in any of the mutants (Table 2).
Detection of mutations in ramR-ramA intergenic region showed that SI group had a T18P mutation and SE group had an M83T substitution; no ramR mutation was detected in the ST group. Following complementation of a ramR mutant with wild-type ramR in SI and SE, there was no change in either the MIC values or AcrAB-TolC efflux pump expression levels. This indicated that these were nonsense mutations (Table 2).
Relative expression analysis of acrA, acrB, tolC, and ramA
One mechanism for antibiotic resistance is an increased expression of efflux pumps. We did not find any overexpression of AcrAB-TolC efflux pump in three parental strains compared with the S. enterica Typhimurium SL1344 (Table 3). Expression of ramA in the mutants increased from ∼13 to 111-fold compared with that in SL1344. In particular, expression levels of ramA in SI-M1, SI-M3, SI-M4, SE-M1, and ST-M4 mutants increased ≥100-fold (Table 3).
Relative expression levels are compared with those in S. enterica Typhimurium SL1344, data in the brackets which are compared with respect parental strains.
The increase in ramA expression was quite different between SI, SE, and ST. In SI, ramA expression was biphasic: a 100-fold increased expression level in mutants SI-M1, SI-M3, and SI-M4, whereas about 50-fold increase in SI-M2. In SE and ST, the increases of ramA expression levels were linear. The range of increase was gradually reduced in SE mutants (∼99- to 56-fold) and increased in ST mutants (∼13- to 103-fold) (Table 3).
Expression levels of acrA, acrB, and tolC were generally only slightly increased in most mutants, but this increase did not correlate with ramA levels. In SI-M4, a slightly decreased expression of acrA was found (0.97-fold). In ST-M3 and ST-M4, acrA and/or acrB expression increased over 10-fold compared with that of SL1344 control strain (Table 3). When compared with their respective parental strains, decreased expression levels of acrA, acrB, and tolC were found in five mutants (SI-M2, SI-M4, SE-M2, SE-M4, and ST-M2) (Table 3, data in brackets).
Accumulation of ciprofloxacin
Ciprofloxacin accumulation in three parental strains was low in the absence of efflux pump inhibitors. SI had lower ciprofloxacin accumulated than the other two strains at the same time point. Five minutes later after addition of ciprofloxacin, there was almost no accumulation of ciprofloxacin in SI. Addition of carbonyl cyanide m-chlorophenylhydrazone (CCCP), a proton motive force uncoupler, increased ciprofloxacin accumulation in all three parental strains. CCCP induced a very rapid and significant increase in cell-associated ciprofloxacin in SI; there was almost no ciprofloxacin accumulation in SE. In ST, a weak accumulation of ciprofloxacin was found only after a prolonged time (about 25 min later) of ciprofloxacin pressure (Fig. 1).
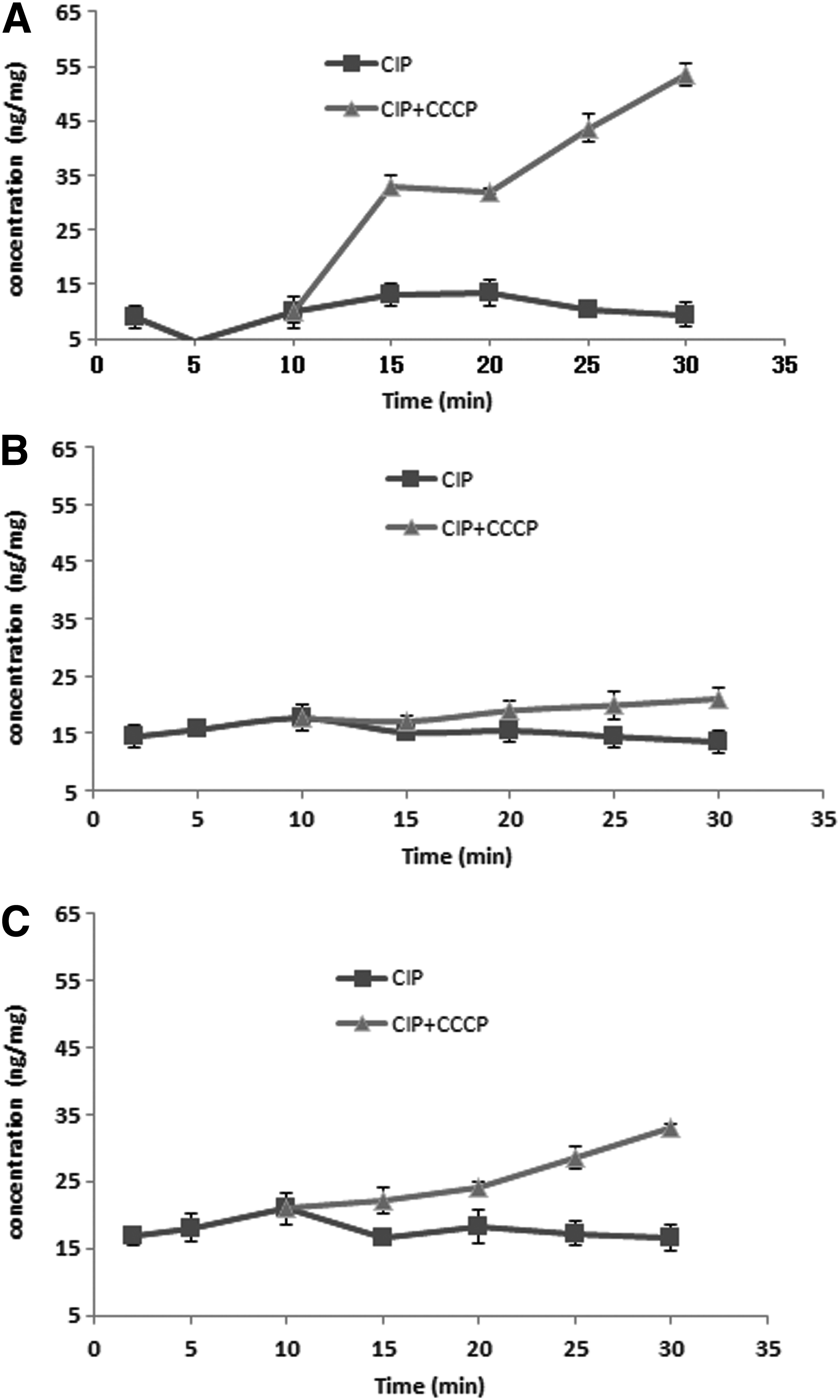
Accumulations of ciprofloxacin in three different serotypes Salmonella with or without CCCP.
Discussion
The development process of flouroquinolone resistance in three most prevalent Salmonella enterica serovars (Typhimurium, Enteritidis, and Indiana) isolates under ciprofloxacin selective pressure were described in this study. Differences in the resistance mechanisms during resistance development process were found among the serovars. In particular, SI developed resistance more easily than SE and ST. S. enterica serovar Indiana isolates were first identified in 2005 in foods of animal origin in China 26 and showed resistance to multiple antimicrobial agents. Subsequently, the S. enterica serovar Indiana increased rapidly and showed a higher rate of antimicrobial resistance compared with other serotypes in China.9,10,13–15 Our previous study on the Salmonella isolates recovered from food-producing animals also suggested high prevalence and multidrug resistance of Indiana isolates. 27 The more rapid resistance and higher mutation frequencies of serovar Indiana isolates from food animals during the resistance development process should be a cause for concern, which may be a potential threat to public health.
Mutation frequency shows a cross section of the bacterial population at a given time and reflects not only the mutation rate, but also the history of the population before selection is applied. 22 In this study, we compared the effects of ciprofloxacin pressure on the development of resistance in SI, SE, and ST, to ensure that the drug-resistant strains selected were stable rather than the result of adaptive growth; all strains in every procedure were cultured five generations in MH broth without antibiotic. The result demonstrated that SI presented a mutation frequency of up to 10−3 and quinolone resistance developed more rapidly and to a greater extent than SE and ST after ciprofloxacin exposure. This may explain why the Salmonella serotype Indiana strains always had multidrug-resistant phenotypes and became more and more prevalent in some provinces of China.3,13,14,26,27
It has been suggested that accumulation of multiple mutations in several genes correlates with the evolution of fluoroquinolone resistance. A high mutation frequency may play an important role in the development of fluoroquinolone resistance by increasing the rate of accumulation of new mutations. 28 Increasing mutation rates can also drive the development of fluoroquinolone resistance. 29 In the present study, increased mutation frequencies were found as the mutation numbers increased. Especially in SI, the additional gyrA D87G and parC T57S mutations made the mutation frequency increase two orders of magnitude and mutation frequency increased to 10−3. Under the same selective pressure, in SE and ST, the gyrB E466D/S464F also increased the mutation frequency, but the level was still only at 10−6/10−7. These data suggest that existence of the gyrA87 mutation may be crucial for increasing mutation frequencies under the quinolone selective pressure. Considering that the signal gyrA83 mutation cooperated with gyrA87 and/or another target such as parC mutations could contribute to high levels of resistance to fluroquinolones.30,31 Thus, existence of the gyrA87 mutation probably facilitates the development of fluoroquinone resistance.
Previous researches have suggested a correlation between a particular Salmonella serotype and a particular mutation. For example, gyrA mutations at Ser83 and Asp87 may not be equally distributed among different serovars. 21 There is also a highly significant correlation between resistance and mutations at gyrA Ser83 in S. enterica serovar Hadar 32 ; and the positions of the gyrA substitutions varied with the serovars. 33 We detected double gyrA mutations in high-level resistance of SI mutants and only a single gyrA 87 position mutation in SE and ST mutants. In the Enterobacteriaceae, in general, high-level fluoroquinolone resistance from gyrA 83 and 87 mutations often requires an additional single and/or double mutation in parC (80 and 84 positions). When we examined parC mutations, the 80/84 position mutations were not present, although the parC T57S substitution was observed in mutants of all three Salmonella serovar strains. This parC T57S mutation was first identified in S. enterica serovar Enteritidis strains. 34 Previous reports suggested the absence of the parC Thr57Ser mutation in S. enterica serovar Typhimurium isolates,16,33,34 but our study found this mutation in ST, which was also consistent with a recent report. 35 It will be interesting to investigate the relationship between Salmonella serotypes and the mutation(s) carried out using larger sample sizes.
In our study, none of three S. enterica serovar isolates contained a parE mutation. Mutations in gyrB were found in both SE (E466D) and ST (S464F), but not in SI. In addition, both the gyrB E466D and gyrB S464F mutations were confirmed to contribute to fluoroquinolone resistance.36,37 The gyrB E466D mutation was first detected in an S. enterica serovar Enteritidis strain 36 and was also found only in SE mutants (SE-M2, M3 and M4). In ST, the gyrB S464F change was only associated with high-level resistance (ST-M4) and suggests that the gyrB S464F must have an important role in the development of high-level ciprofloxacin resistance of ST mutants.
Measurement of ciprofloxacin accumulation showed great variation between three parental isolates. In particular, SI had a rapid and significant increase in cell-associated ciprofloxacin compared with SE and ST, which also suggested higher efflux pump activity in SI. A weak accumulation of ciprofloxacin in ST after a prolonged time of ciprofloxacin pressure indicated the possible role of efflux pump in the later stage of resistance development.
There are at least nine multidrug efflux pumps in Salmonella and the AcrAB-TolC efflux pump is the best characterized multidrug resistance (MDR) system. All mutants showed reduced susceptibility or resistance not only to quinolones, but also to other antimicrobial classes in this study. This suggests a role for efflux pumps with broad substrate specificities, the AcrAB-TolC pump can efflux many different classes of antibiotics, including quinolones, β-lactams, tetracycline, and chloramphenicol. 38 Our results may indicate that the role of efflux pumps in the development of resistance is underappreciated. Inhibition of efflux pumps could have great effect on the resistance development which also provides opportunity for potential intervention. 19
It has previously been demonstrated in Salmonella that the AcrAB-TolC efflux pump is regulated by the global regulator RamRA. 5 In the present study, the expression level of RamA in all derived mutants was higher than that of their parental strains, which suggested the crucial role of AcrAB-TolC efflux pump in the development of fluoroquinolone resistance in different S. enterica isolates. The increase in ramA expression levels in SI serial mutants was not linear indicating that AcrAB-TolC efflux pump played different roles in different resistance development periods. This also suggests a role in the early reduction of susceptibility and the final high-level resistance in SI. In SE mutants, the fold increase of ramA expression decreased rapidly in the development of resistance. Conversely, ramA expression rise quickly as higher levels of resistance developed in ST. This difference in change of ramA expression in the mutants of different serotypes may relate to other FQ cooperative resistance mechanisms, such as target alterations and expression of other efflux pumps.
Our results indicated that (I) the AcrAB-TolC efflux pump plays an important part in the first mutational stage and facilitates high-level resistance development along with target-site mutations in SI. This may also assist SI to gain a higher mutation frequency and to develop resistance more easily. (II) In SE, target-site mutations in gyrA (D87G) and gyrB (E466D) and/or other efflux pumps besides the AcrAB-TolC efflux pump are important in the development of FQ resistance. (III) In ST, the AcrAB-TolC efflux pump had a crucial impact on the whole resistance development process. Target-site mutations in gyrA (D87N) could not fully mediate the gradually increasing ciprofloxacin resistance until the mutation in gyrB (S464F) developed. Our study also suggests that clinical strains with reduced susceptibilities to fluoroquinolones might be crucial in the development of FQ resistance. These strains can become highly resistant by the accumulation of target gene mutations together with the active efflux pumps under the ciprofloxacin selective pressure.
Conclusion
In summary, high-level FQ resistance in three S. enterica serovar strains were all linked to target gene mutations and the AcrAB-TolC efflux pump as well as other efflux pumps. The two main quinolone resistance mechanisms operate in a highly coordinated manner in the development of resistance. The role of these mechanisms in the development of resistance varies among the three S. enterica serovar strains. However, all three developed high-level FQ resistance and other antibiotics under persistent ciprofloxacin pressure. The SI strains developed resistance easier and faster, which may lead to future problems. Therefore, fluoroquinolones should be used prudently in both humans and food animals.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31272602) and the Program for Changjiang Scholars and Innovative Research Team in the University of Ministry of Education of China (Grant No. IRT13063).
Author Disclosure Statement
No competing financial interests exist.