
Abstract
Integrons are prokaryotic genetic elements known to carry and exchange antibiotic resistance gene cassettes through a site-specific recombinase called integrase. In this work, 107 Aeromonas isolates from environmental origin, including fish, water, and sediments, were investigated for the presence of integrons. Using specific primers for Class 1, 2 and 3 integrases, only Class 1 and Class 2 integrons were detected. Detection of Class 2 integrases and their associated variable regions required two rounds of polymerase chain reaction (PCR). Sequencing of the intI2 amplicons confirmed them as integrase-derived products. Class 1 integrons were detected in 26 out of 107 isolates. PCR amplification of the variable regions associated to these integrons revealed an outstanding homogeneity, 25 of them having variable regions with an identical dfrA12-orfF-aadA2 cassette array and one integron carrying only the dfrA16 cassette. To assess clone diversity, chromosomal DNA from isolates was subjected to enterobacterial repetitive intergenic consensus-PCR (ERIC-PCR), which discarded clonality in all instances. Class 2 integrons were surprisingly more prevalent than Class1 integrons, being detected in 60 out of 107 isolates. Forty-six of them showed a unique ERIC profile, while the remaining 14 strains displayed profiles that could be grouped in five different patterns. Cassette arrangements of all Class 2 variable regions were those described as the most prevalent (dfrA1-sat2-aadA1). A rather startling result of this work is the sensitivity to trimethoprim, streptomycin, and streptothricin of most strains, despite the presence of the cognate resistance genes. To know the integron distribution in environmental Aeromonas species, a phylogenetic reconstruction was done using rpoD/gyrB or rpoD/gyrA gene sequences. Isolates bearing these elements corresponded to Aeromonas hydrophila, Aeromonas veronii, Aeromonas salmonicida, Aeromonas dhakensis, Aeromonas sanarellii, Aeromonas taiwanensis, Aeromonas media, Aeromonas caviae, Aeromonas jandaei, and Aeromonas sp. This work revealed an unusual high incidence of Class 2 integrons and a low variability of cassette arrangements in environmental Aeromonas species.
Introduction
Integrons are genetic elements able to incorporate, express, and exchange gene cassettes, frequently but not exclusively, encoding antibiotic resistances. Cassette exchanges are mediated by a recombinase able to excise and reinsert them through site-specific recombination between flanking att sequences. Cassette genes lack, for the most part, a promoter and their excision and reinsertion at the attI site, proximal to a Pc promoter, renders them transcriptionally active. 1 Cassette exchange thus represents a way of saving metabolic resources in bacteria facing antibiotic threats, by keeping distal cassettes as a genetic reservoir. 2
Integrons are classified as Class 1 to Class 4 on the basis of the sequences of their associated integrases. Class 1 integrons are described as the most prevalent and the most frequently linked to antibiotic resistance in Gram-negative bacteria. 3 Class 2 integrons are second in abundance, while Class 3 integrons have the lowest prevalence of all. 4 On the other hand, Class 4 integrons, also called superintegrons, are more complex structures having a large number of cassettes, normally not associated to antimicrobial resistance. Superintegrons, located on the chromosome, are regarded as nonmobile integrons. These structures have been described mainly in Vibrio. 5
The presence of Class 1 integrons in Aeromonas has been well documented and the prevalence of these elements is around 21%, while reports on the presence of Class 2 integrons in Aeromonas are very scarce.6–9 Class 2 integrons display a limited variability, probably resulting from the presence of a TAA stop codon at position 179 of the integrase gene, which leads to an incomplete and therefore nonfunctional protein. Only a few functional intI2 genes encoding the 385 amino acid residues protein have been described. Class 2 integrons are commonly associated to Tn7 transposon. 10 A single report on the presence of Class 3 integrons in Aeromonas exists. 11 This work aims to establish the relative frequency of Class 1, 2, and 3 integrons in environmental Aeromonas and their variability of associated resistance cassettes.
Materials and Methods
Isolation and presumptive identification of Aeromonas strains
Strains were isolated from water, sediment, and wild fish collected from different regions of Mexico. Samples (25 g or 25 mL) were homogenized in 225 mL of Trypticase Soy Broth (TSB) and incubated at 37°C for 18 hours with shaking at 200 rpm. Dilutions of these cultures were spread onto Bile Salts Brilliant Green Starch (BBGS) agar plates and Starch Ampicillin Agar (SAA). Plates were incubated at 37°C during 18 hours; then, Lugol's solution was added and amylase-positive isolates with a clear zone surrounding the colony were presumptively identified as Aeromonas. 12 Isolates were subjected to genus confirmation by biochemical tests. 13 Strains identified as Aeromonas were preserved in cryotubes containing TSB supplemented with 20% glycerol and stored at −70°C.
Genus confirmation by mass spectrometry
Isolates were identified to the genus level using the MALDI-TOF VITEK® MS (Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight Mass Spectrometry) system. Bacterial strains were streaked onto Luria plates and incubated at 37°C for 18 hours. Isolated colonies were spread on a glass slide and covered with a α-cyano-4-hydroxycinamic acid matrix solution. Slides were introduced into the VITEK MS PLUS system (bioMérieux, Marcy l'Etoile, France) to obtain the spectrum of each sample. Ribosomal protein profiles were compared with those contained in the VITEK MS In Vitro Diagnostic (IVD) version 3.0.0 database.14,15
rpoD, gyrB, and gyrA amplification and sequencing for phylogenetic analysis and species identification
Chromosomal DNA was obtained by standard protocols. 16 For the polymerase chain reaction (PCR) amplification of the rpoD, gyrB, and gyrA genes, primers reported by Martinez-Murcia et al. 17 were used (Table 1). Amplified products were sequenced by the Sanger method using the ABI3730XL system. Sequences were edited manually using FinchTV v1.4.0 (The Geospiza Research Team, 2004–2006). The resulting dataset together with the 36 sequences of the same Aeromonas genes available at the GenBank was assembled using Seaview v4.6. Oceanimonas spp. was used as an out-group. 18 Gene sequences were used for the phylogenetic analysis, which was performed using the maximum likelihood method, as implemented in the program LRT-PHYML. 19 The evolutionary model of nucleotide substitution was obtained by using the Smart Model Selection. 20
Primers Used in This Work
ERIC-PCR, enterobacterial repetitive intergenic consensus polymerase chain reaction; VarI, variable region 1; VarII, variable region 2.
Detection of integron associated genes
The presence of Class 1, 2, and 3 integrons and their variable regions was assessed by PCR amplification of the integrase coding sequences using the primers described previously (Table 1).21–23 Amplification reactions included 12.5 μL PCR Master Mix (Thermo Scientific), 200–300 ng template DNA, and 10 pmol/μL of each primer in a final volume of 25 μL. Amplifications involved DNA denaturation at 94°C for 3 minutes followed by 30 PCR cycles (DNA denaturation at 94°C for 30 seconds, primer annealing at 52°C [intI1 gene] or 61°C [intI2 and intI3 genes] for 30 seconds, and extension at 72°C for 30 seconds) and a final extension step at 72°C for 5 minutes. Amplifications of all variable regions were done as before, except PCR Master Mix was substituted for Phusion Flash High-Fidelity PCR Master Mix (Thermo Scientific), annealing temperature was 60°C for the variable region 1 (VarI) and 65°C for the variable region 2 (VarII) and the extension time was increased to 2 minutes. Escherichia coli C3297 bearing Class 1 and Class 2 integrons was used as positive control. 24 Gene amplifications from Class 2 integrons required two consecutive PCR rounds to improve amplicon detection and purification.
Characterization of the resistance cassettes and promoter regions
Products of the variable region were sequenced and sequences were compared with the GenBank nucleotide sequence database using the BLAST server at NCBI. Sequences with significant similarity were aligned using the Multiple Alignment using Fast Fourier Transform (MAFFT) Alignment tool from the EBI web page. Variable region cassette arrangements were confirmed by restriction with EcoRI and HindIII (VarI region) or SseI (VarII region; Thermo Scientific). Restriction fragments were separated on 1.5% agarose gels in TAE buffer at 6 V/cm and stained with ethidium bromide. Association of Class 1 and Class 2 integrons with the 3′ qacEΔ-sul1 or Tn7 was established using primers in-F/in-B2 and attI2-F/23CS, respectively. 25 PCR conditions were those described for the Var regions. Promoter regions from Class 1 and 2 integrons were amplified using primers intPcFW/dfr12aminoQ 26 and intI2.1FW/dfrA1RV (Table 1).
Enterobacterial repetitive intergenic consensus-PCR analysis
Clonality of strains within a species was established by means of an enterobacterial repetitive intergenic consensus (ERIC)-PCR analysis (Table 1).27,28 PCR products were separated by 10% polyacrylamide (Sigma) gel electrophoresis on TAE buffer at 100 volts, during 90 minutes. DNA bands were stained with ethidium bromide (0.5 μg/mL) and visualized under ultraviolet light. The resulting band pattern was used to create an absence-presence matrix. Similarity among clones was estimated using the Dice similarity index. The resulting similarity matrix was used to perform a cluster analysis using the unweighted pair group method with arithmetic mean. To evaluate the resulting tree, the cophenetic correlation coefficient was calculated with the program Past v3.0. 29
Antimicrobial susceptibility testing
The minimum inhibitory concentration for trimethoprim, streptomycin, and streptothricin (Sigma) was established by the broth microdilution method using 96-well microtiter plates, following the protocol of the Clinical and Laboratory Standards Institute (CLSI). 30 E. coli ATCC 25922 and Aeromonas hydrophila ATCC 7966 were used as sensitive control strains. Pseudomonas aeruginosa ATCC 27853, A. hydrophila 6479, and E. coli C3297 were used as resistant controls.26,31
Results
Phenotypic and genotypic identification of Aeromonas strains
Biochemical tests presumably identified 109 environmental isolates as Aeromonas. Genus was confirmed in 107 isolates by the MALDI-TOF VITEK-MS system. Species assignation was done only in strains bearing at least one integron (n = 77). This was achieved by sequencing of the rpoD and gyrB genes. We resorted to amplifying and sequencing the gyrA gene whenever amplification of rpoD or gyrB was not effective. The species found were as follows: 39 A. hydrophila (50.65%), 9 Aeromonas veronii (11.69%), 8 Aeromonas salmonicida (10.39%), 7 Aeromonas dhakensis (9.09%), 4 Aeromonas sanarellii (5.19%), 4 Aeromonas taiwanensis (5.19%), 2 Aeromonas media (2.6%), 2 Aeromonas sp. (2.6%), 1 Aeromonas caviae (1.3%), and 1 Aeromonas jandaei (1.3%) (Supplementary Table S1). Sequences of housekeeping genes were deposited at the Genbank (gyrB: MN659202-MN659262, rpoD: MN659263-MN659328, and gyrA: MN659186-MN65920).
Integron detection and characterization
The 107 confirmed Aeromonas strains were examined for Class 1, 2, and 3 integrons by PCR amplification of the integrase genes. This search resulted in 26 isolates (24%) carrying Class 1 and 60 (56%) with Class 2 integrons. Nine strains contained both types of integrons, while no single isolate with a Class 3 integron was found. PCR amplification of the VarI region of 25 out of 26 strains yielded the same amplicon. Analysis of the amplicons showed that all strains shared the dfrA12-orfF-aadA2 array coding for trimethoprim and streptomycin resistance, with a single strain carrying the dfrA16 cassette. All Class 1 integrons carried a qacEΔ-sulI region at their 3′ end, as confirmed by the production of a slightly larger amplicon when a second reverse primer (in-B2) was used (results not shown). Fifty-seven out of sixty Class 2 integrons displayed the dfrA1-sat2-aadA1 array, while no variable region could be detected in the three remaining templates. The sat2 cassette in this array codes for streptothricin resistance. All Class 2 integrons with a detectable variable region were found to be associated with Tn7 transposon (Fig. 1).
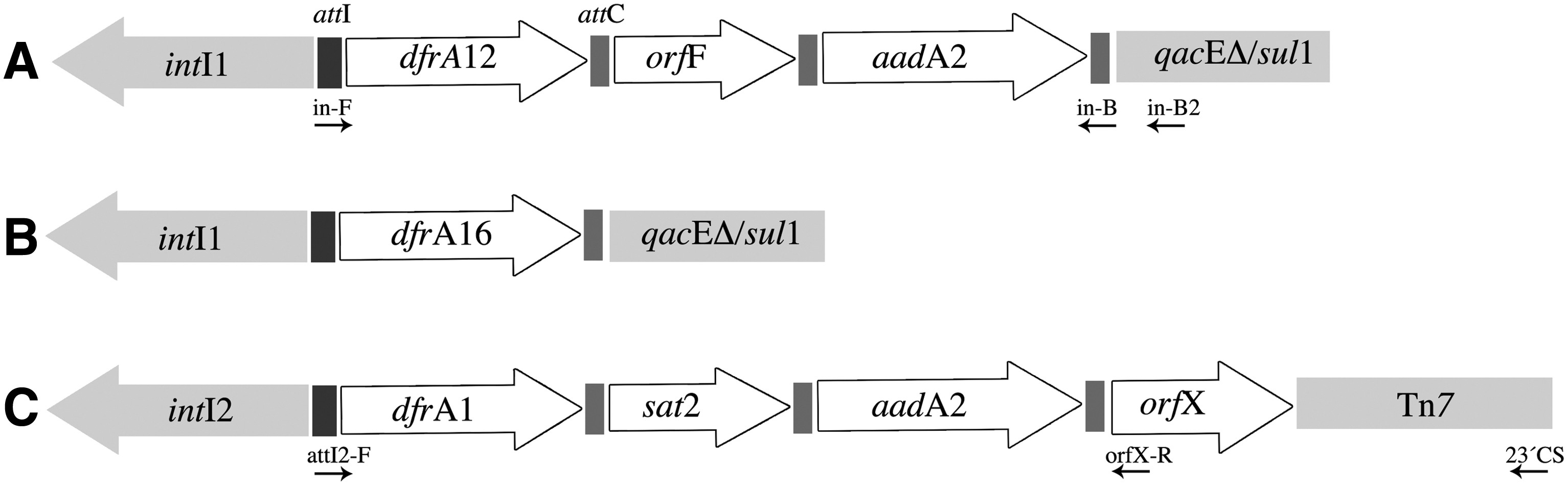
Variable regions of Class 1 and 2 integrons from environmental Aeromonas strains.
Promoters from the Class1 integrases and their cassettes were identical in all strains. The same was true for all Class 2 promoters (Fig. 2).

Promoter regions of Class 1 and Class 2 integrons.
Clonality analysis
Since some isolates originated from the same geographic region, Class 1 strains belonging to the same species were subjected to ERIC-PCR to discard clonality. ERIC profiles were unique for all Class 1 integron-positive strains of the same species. As with Class 1 positive isolates, 46 Class 2 positive strains displayed unique ERIC profiles, while the remaining 14 strains shared ERIC profiles with at least another clone forming 5 clonal groups (Supplementary Fig. S1). Some strains belonging to the same group were isolated from different geographic areas and in a different year.
Resistance profiles
Owing to the ubiquitous presence of gene cassettes encoding resistance to trimethoprim and streptomycin in the two types of integrons found in environmental Aeromonas, we decided to determine the minimal inhibitory concentration of these antibiotics. Strikingly, most strains, even those bearing the two types of integrons, were sensitive to any of the antibiotics. Only three strains, two with a Class 1 and one with a Class 2 integron, were resistant to both antibiotics. Three and four Class 2 strains were resistant only to trimethoprim and streptomycin, respectively. Only one strain was resistant to streptothricin (Fig. 3 and Supplementary Table S1).
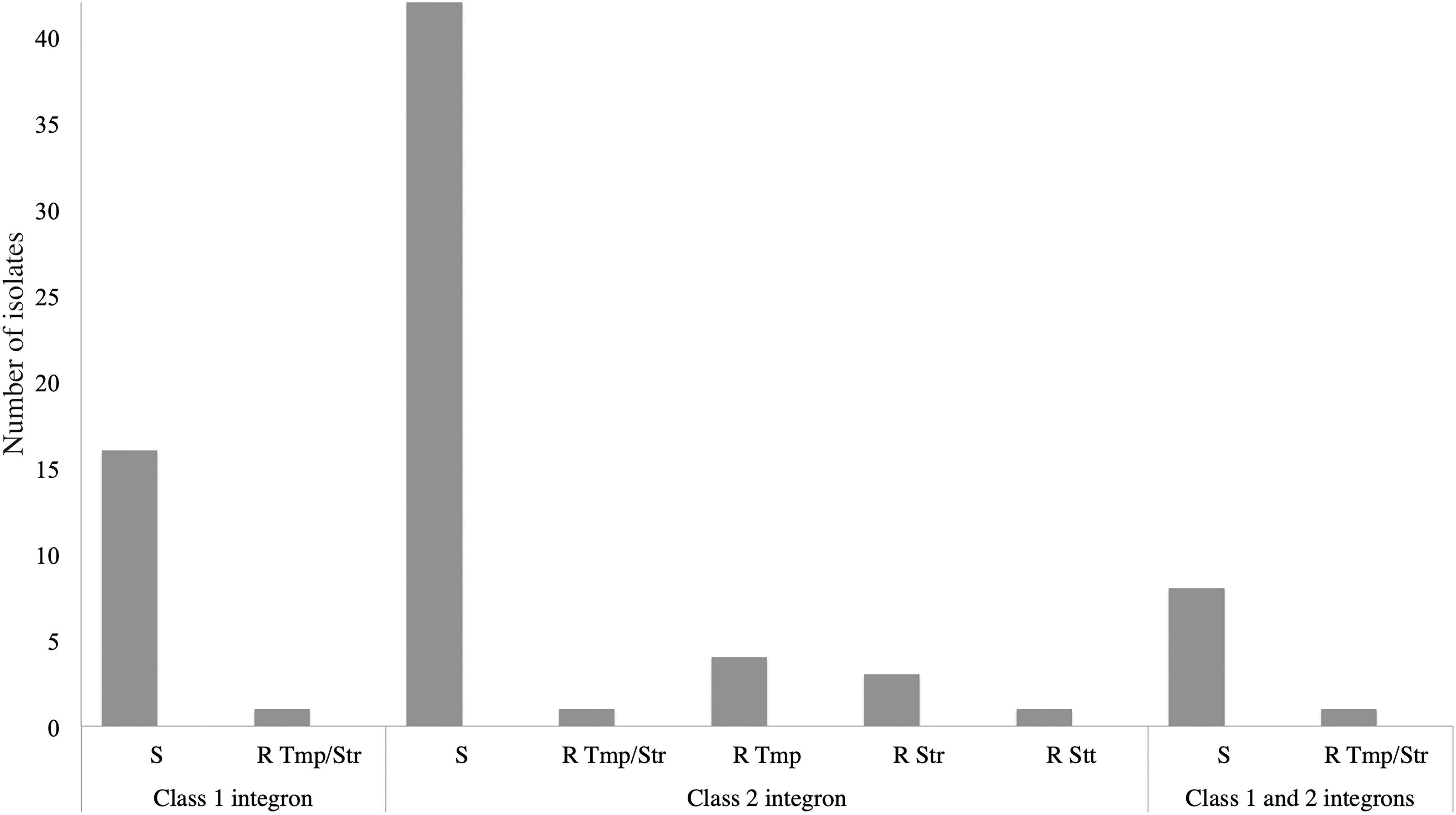
Trimethoprim, streptomycin, and streptothricin resistance frequencies. Seventy-seven strains harboring integrons were tested for antimicrobial resistance (17 Class 1, 51 Class 2, and 9 Class 1 and 2). R, resistant; S, sensitive; Str, streptomycin; Stt, streptothricin; Tmp, trimethoprim.
Discussion
Bacteria belonging to the genus Aeromonas maybe resistant to antibiotics from several families.32–35 Resistance to these antibiotics is sometimes mediated by integrons. 3 These elements have been previously reported in Aeromonas strains isolated from water, fish, and clinical samples.8,13,36 Class 1 integrons have been so far informed as the most prevalent in this genus, with values ranging between 6.9% and 64.6%.37,38 The incidence found in this work was 24% (26/107), which is within the observed range. Literature describes multiple cassette arrays in Class 1 integrons, although the cassette diversity is rather limited. Some particular arrangements are widespread in nature, exhibiting a pandemic distribution, 39 with aadA and dfrA cassettes among the most abundant.40,41 Class 1 integrons of environmental Aeromonas strains studied here contained almost exclusively the dfrA12-orfF-aadA2 arrangement. Although a high incidence of this particular arrangement has also been described in Aeromonas spp. and Klebsiella spp.,42,43 this work reports the highest incidence found so far. It is important to notice that this limited variability in Aeromonas is not necessarily a rule. Cassette variability could be associated to selective pressure caused by the indiscriminate use of antibiotics, since Aeromonas strains isolated from clinical samples in Mexico and Taiwan displayed a wider variety of cassette arrangements.13,44 Sequences of the Class 1 integrases studied in this work were identical to the one found in integrases from two clinical isolates proven to be completely functional. 26 This corroborates that the arrangement conservation observed in environmental integrons is not due to a defective integrase.
The high incidence of Class 2 integrons was an important finding in this work, since the few reports of these elements in Aeromonas show a very low incidence,8,45 and in some instances, the variable region was not characterized.46,47 First attempts to amplify the Class 2 integrase genes in this work produced faint bands of the right size, in contrast with the positive control, which generated a robust amplicon. Changes in amplification parameters, including the use of special buffers to amplify GC-rich templates like the Aeromonas DNA, did not improve the amplicon yields. Since some amplification problems are solved with a reamplification, 48 we decided to perform an additional PCR round in all samples. Amplicons derived from the second PCR were isolated and their identity confirmed by DNA sequencing. This sequencing showed that primer binding sites were totally complementary to the primer sequences, which indicates that the low amplicon yields in the first PCR round do not stem from a lack of primer complementarity.
Most reports indicate that Class 1 integrons are more abundant than Class 2 integrons.8,45 In this study, Class 2 integrons were two times more frequent than Class 1 integrons, even considering the occurrence of several clones among those bearing Class 2 integrons. This result is analogous to those obtained with Shigella spp., Acinetobacter baumanii, Enterobacter cloacae, and bacteria from the Proteae family.49,50 Mendes Moreira et al. 51 suggested that the Proteae family, with 31% Class 2 integrons, could represent a reservoir of these elements. Following this line of reasoning, Aeromonas could be considered a reservoir for these integrons too. Our findings suggest that the low efficiency of amplification of Class 2 associated genes may lead to an underestimation of these integrons. Analysis of the VarII regions identified in this work indicates that the vast majority of Class 2 integrons share the dfrA1-sat2-aadA1 cassette arrangement commonly found in this type of integrons. As seen with other Class 2 integrases, 10 IntI2 coding sequences of all strains analyzed in this work were interrupted by a stop codon. This explains, in part, the arrangement homogeneity observed.
It is really interesting to observe that integrons found in this work contain only the dfrA and aadA cassettes. Multiple dfrA gene variants exist, most of them located in integrons as cassette genes.52–55 The wide distribution of the dfr genes reveals a different behavior compared to many other cassettes. A retrospective study in Sweden revealed that after a 2-year reduction in the use of trimethoprim, the frequency of resistant bacteria did not change significantly. 56 This work shows that trimethoprim resistance cannot be easily reverted. Another study performed in E. coli evidenced that the dfr gene is prone to suffer mutations after exposition to subinhibitory doses of the compound. 57 This could explain the rising of multiple variants of the dfr genes, although it does not clarify why this gene is commonly associated to integrons.
Streptomycin is a secondary metabolite formed by Streptomyces. Resistance to this antibiotic is given by the aadA genes coding for an aminoglucoside 3"-adenylyltransferase. Just like for the dfrA genes, several variants of the aad gene exist.40,52 The wide variability of the aad genes could be related to the early presence of Streptomyces on earth. In this regard, Kurakov et al. 58 described the presence of the aadA27 gene in a non-cassette manner in an elder lineage plasmid isolated from permafrost samples.
Despite the presence of two antibiotic resistance genes in both classes of integrons, most strains were sensitive to trimethoprim and streptomycin. We would expect that all strains were resistant at least to trimethoprim, considering that the dfrA cassette is located in the first position of the arrangements. Pc promoters with different strength have been described.10,59,60 PcH1 followed by P261,62 promoters was found in all Class 1 integrons. This promoter combination leads to intermediate gene expression levels. This finding shows that the sole presence of a gene cassette does not guarantee the resistance to the antibiotic in environmental strains and contrasts with a previous report where integrons from two clinical isolates with DNA sequences identical to those described in this study make these isolates resistant to both antibiotics. 26 Previous work in our laboratory showed that the cloned integron of one of these strains render E. coli resistant to both antibiotics. All Class 2 integrons from the strains analyzed in this work contain promoters Pc2A and Pc2B, which are also considered intermediate strength promoters. 10 As with Class 1 integrons, most strains were sensitive to trimethoprim, streptomycin, and streptothricin. These results suggest the participation of other cell factors in the expression the gene cassettes.
Sequencing of the rpoD/gyrB or rpoD/gyrA genes allowed the assignation of isolates to the following species: A. hydrophila, A. veronii, A. salmonicida, A. dhakensis, A. taiwanensis, A. sanarellii, A. media, A. caviae, and A. jandaei. Two isolates could not be identified this way. The most frequently isolated species was A. hydrophila. This species is recognized as one of the most prevalent worldwide. 63 Strains classified as A. salmonicida were able to grow at 37°C and did not produce pigment. This atypical behavior has been previously documented. 64 A. hydrophila, A. caviae, and A. veronii, the most frequently reported strains in México, did not show the same prevalence in this work. A. dhakensis, A. sanarellii, A. taiwanensis, and A. jandaei have rarely or never been reported in Mexico. This is probably due to the improvement of the identification methods.65–67 Our isolation strategy included the usual ampicillin-containing SAA medium and an isolation medium without ampicillin (BBGS). This was used to allow the growth of ampicillin-sensitive strains. Not a single ampicillin-sensitive isolate was found, discarding the presence of Aeromonas trota in the analyzed samples. 68 The number of class 2 integrons in the different species is directly related to the frequency of isolation. Interestingly, all A. salmonicida isolates carried only class 1 integrons.
Results reported in this work show a low variability of gene cassettes in integrons Classes 1 and 2 and a remarkably high incidence of Class 2 integrons in environmental Aeromonas strains. An unexpected result of this work is the lack of antibiotic resistance to cassette-encoded antibiotics in the strains examined.
Footnotes
Acknowledgments
We are grateful to Dr. Rosa del Carmen Rocha Gracia, Centro de Investigaciones en Ciencias Microbiológicas, Instituto de Ciencias, and Benemérita Universidad Autónoma de Puebla, Puebla, Mexico, for the generous gift of E. coli C3297 strain.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Secretaria de Investigación y Posgrado del Instituto Politécnico Nacional grants 20171019, 20181641, and 20196132. J.E.O.-O. was a Consejo Nacional de Ciencia y Tecnología (CONACYT) and Estímulo Institucional de Formación de Investigadores (BEIFI) fellow. E.C.-Q. is a COFAA (Comisión de operación y foment de actividades academicas), EDI (Estimulo al desempeño de investigadores), and SNI (Sistema Nacional de Investigadores) fellow. A.P.-V. is an EDI and SNI fellow. M.G.A.-A. is a COFAA, EDI, and SNI fellow.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.