
Abstract
Vancomycin-resistant enterococcal (VRE) bacteremia is associated with higher mortality rates and longer hospitalizations than vancomycin-sensitive enterococcal (VSE) bacteremia. A 67-year-old man with a right psoas abscess and pacemaker-associated tricuspid valve endocarditis in September 2020 grew VSE Enterococcus faecium from blood cultures that cleared after administration of intravenous vancomycin and gentamicin. Subsequently, he underwent tricuspid valve repair, pacemaker removal, and partial lead extraction. Valve and postoperative blood cultures grew VRE E. faecium, which cleared after administration of intravenous daptomycin. One VSE and two VRE isolates were collected and sequenced. All isolates belonged to E. faecium multilocus sequence type ST17 and were closely related, having <20 mutations in pairwise genome comparisons. Vancomycin resistance was due to the acquisition of a plasmid-encoded VanA operon. None of the isolates encoded the virulence factors asa1, gelE, cylA, or hyl; all encoded a homologue of efaAfm. VSE E. faecium, but not VRE E. faecium isolates, encoded a glucose transporter gene mutation. Two VRE E. faecium isolates formed more robust biofilms than the VSE E. faecium isolate (p < 0.001). The VRE E. faecium isolates, which generated larger biofilms than the VSE E. faecium isolate, could have remained protected in the heart valve and only caused bacteremia when disrupted during cardiac surgery. This study demonstrates that bacteria detected in the bloodstream of patients with endocarditis may not fully represent the organisms adherent to the cardiac valves or indwelling devices.
Background
Among hospitalized patients, those with vancomycin-resistant Enterococcus (VRE) bacteremia have higher mortality rates and longer lengths of stay than those with vancomycin-susceptible Enterococcus (VSE) bacteremia. 1 Exposure to antibiotics, including vancomycin, can increase the risk of VRE infection. 2 However, the expression of vancomycin resistance is associated with a fitness cost, leading to lower rates of colonization and transmission in the absence of antibiotic pressure. 3 Eight different operons, vanA, vanB, vanD, vanE, vanG, vanL, vanM, and vanN, have been associated with acquired vancomycin resistance in enterococci. 4
VanA-mediated resistance is the most common in the United States, and results in high-level resistance by substituting
Development of a VRE Enterococcus faecium (VREfm) infection in an individual being treated for a VSEfm infection can occur by at least three different mechanisms: if the VSEfm isolate acquires a VanA operon-encoding plasmid, 8 if it carries a disrupted VanA operon that regains functionality, 6 or if the individual becomes subsequently infected with a genetically distinct VREfm isolate. 9
In addition to vancomycin resistance, enterococci possess numerous virulence-enhancing genes that can impact disease, including aggregation substance (AS), gelatinase (Gel), cytolysin (Cyl), enterococcal surface protein (Esp), and hyaluronidase (Hyl). 10 Moreover, studies have demonstrated that gelE and esp from E. faecium are associated with biofilm formation, whereas asa1 and efaAfm improve adherence to endocardial cells. 11
In this report, we used whole genome sequencing (WGS) to characterize the genetic changes underlying the development of vancomycin resistance in an individual with recurrent enterococcal infection, from whom closely related VSE and VRE bacteria were isolated.
In addition, we sought to determine whether the VREfm isolates detected after cardiac surgery possessed other genetic or phenotypic alterations compared with the initial VSE isolate cultured from the patient.
Case Report
A 67-year-old man with a history of a left ureteral stricture requiring nephrostomy tube underwent ureteral reimplantation surgery in January 2020. He developed both VREfm and VSE Enterococcus faecalis (VSEfs) bacteremias between April 2020 and June 2020. In April, his VREfm bacteremia was complicated by cervical 4–5 discitis. In June, his VSEfs bacteremia prompted removal of the left nephrostomy tube. He presented with fever and chills on September 8, 2020. Hospital evaluation revealed left hydronephrosis, a right psoas abscess, and pacemaker-associated tricuspid valve endocarditis.
A summary of culture data and antibiotic courses is outlined in Fig. 1. In September 2020, the patient's VSEfm bacteremia was treated with vancomycin and gentamicin, with subsequent blood cultures demonstrating no growth. On October 5, 2020, the patient underwent tricuspid valve repair, pacemaker removal, and partial lead extraction. The valve and subsequent blood cultures grew VREfm, and antibiotic therapy was changed from vancomycin to daptomycin. The patient completed his course of daptomycin on November 24, 2020. Repeat abdominal CT on December 31, 2020, demonstrated resolution of the psoas abscess. Subsequent bacteremia was not detected through May 2021.
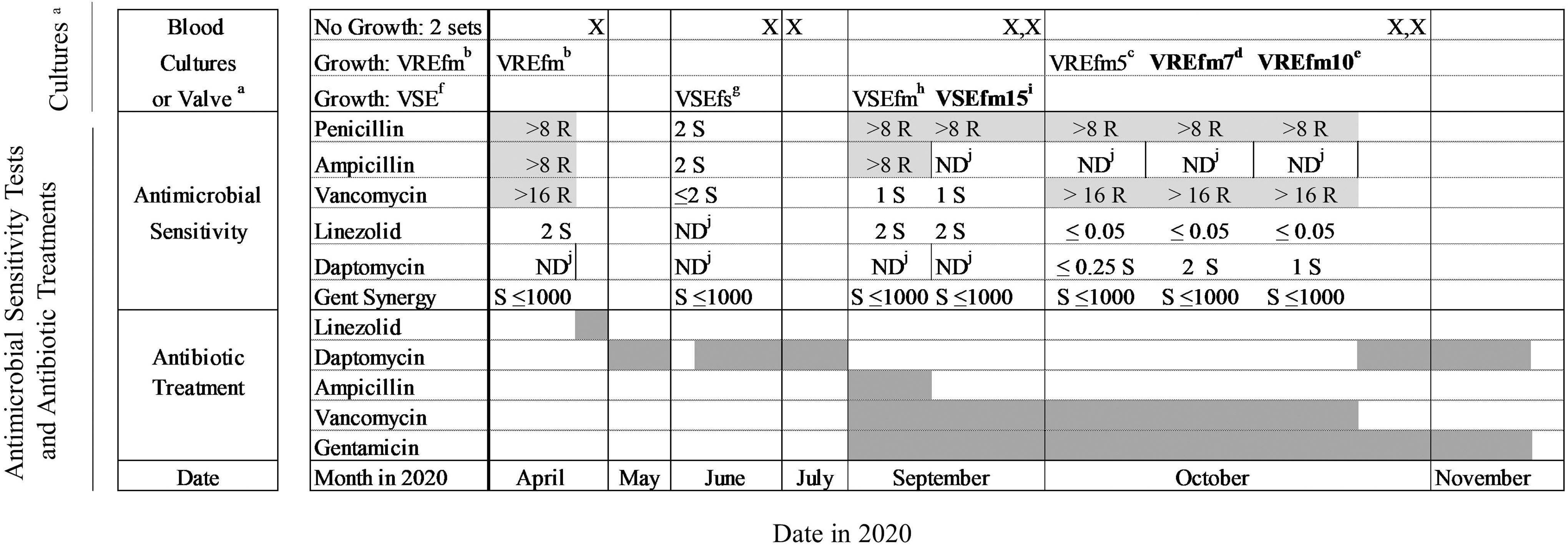
Timeline of bacteremias, antimicrobial sensitivity tests, and antibiotic treatments. aOctober 5 isolate from tricuspid valve; all other isolates are from blood cultures. bVREfm—vancomycin-resistant Enterococcus faecium (fm). cVREfm5—vancomycin-resistant E. faecium (fm)—October 5 tricuspid valve. dVREfm7—vancomycin-resistant E. faecium (fm)—October 7 bloodstream. eVREfm10—vancomycin-resistant E. faecium (fm)—October 10 bloodstream. fVSE—vancomycin-sensitive Enterococcus. gVSEfs—vancomycin-sensitive Enterococcus faecalis (fs). hVSEfm—vancomycin-sensitive E. faecium (fm). iVSEfm15—vancomycin-sensitive E. faecium (fm)—September 15 bloodstream. jND—not done. Bold: sequenced isolates—VSEfm15, VREfm7, VREfm10; light gray, resistant; dark gray, treatment course. VRE, vancomycin-resistant enterococcal.
Methods
The VSEfm isolate detected in the blood culture collected on September 15, 2020 (VSEfm15), was compared with the VREfm isolates detected in the blood cultures collected on October 7 (VREfm7) and October 10 (VREfm10) 2020 (Fig. 1). No rectal swabs for VREfm were collected during the study period. Genomic DNA was extracted from pure overnight cultures inoculated from single bacterial colonies using a Qiagen DNAeasy Blood and Tissue Kit according to manufacturer's instructions (Qiagen, Germantown, MD).
Library construction and sequencing were conducted at the Microbial Genomics Sequencing Center at the University of Pittsburgh using an Illumina Nextera DNA Sample Prep Kit with 150 bp paired-end read length, and libraries were sequenced on the NextSeq WGS platform (Illumina, San Diego, CA). Sequence type (ST) was determined for each genome by comparing seven housekeeping genes. 12 Antimicrobial resistance genes, plasmid rep genes, and virulence factors were assessed using online tools from the Center for Genomic Epidemiology (www.genomicepidemiology.org) and by manual BLAST searching. All enterococcal isolates were screened for the presence and differences in the sequence of glcU, asa1, gelE, cylA, esp, hyl, and efaAfm.
In vitro biofilms were generated with standard sterile 96-well polystyrene plates, as previously described. 13 In brief, an overnight culture of each isolate was diluted 100-fold into brain heart infusion broth supplemented with 0.25% glucose. In total, 200 μL of this dilution was plated into eight replicate wells of a 96-well untreated polystyrene microtiter plate, and plates were incubated for 24 hr at 37°C under static conditions. Planktonic cells were discarded and plates were washed three times with 250 μL 1 × PBS, after which adherent bacteria in each well were stained with 200 μL 0.1% crystal violet (CV) prepared in water.
After incubation for 30 min at 4°C, wells were washed twice with 250 μL 1 × PBS to remove excess stain. Plates were dried and then 250 μL of 4:1 ethanol–acetone was added to each well to solubilize the CV-stained biofilms. After incubation for 45 min at room temperature, the absorbance in each well was measured at 550 nm using a Synergy H1 microplate reader (Biotek, Winooski, VT). The biofilms were compared with the two-tailed t test. Approval by IRB was not required given that this study was performed as an operational activity related to quality improvement.
Results
A total of four different episodes of enterococcal bacteremia were detected between April 2020 and November 2020 (Fig. 1). Between September and October 2020, bacteremias due to both VSEfm and VREfm were detected. Three enterococcal isolates collected from blood cultures were further characterized: VSEfm15 was detected on September 15, VREfm7 was detected on October 7, and VREfm10 was detected on October 10. All three isolates belonged to E. faecium multilocus sequence type ST17. Both the VREfm isolates were found to have acquired the VanA operon along with rep14a, rep17, and repUS15 plasmid replicon-encoding genes, indicating plasmid-encoded vancomycin resistance acquisition.
None of the isolates encoded asa1, gelE, cylA, or hyl. The VSEfm15 isolate and the VREfm7 isolate encoded esp, whereas the esp gene sequence in the VREfm10 isolate was disrupted. All three encoded a homologue of efaAfm. The VREfm genomes each encoded 15 or 16 single nucleotide polymorphisms (SNPs) when compared with the VSEfm genome, confirming that the three genomes were closely related to one another. The two VREfm genomes differed from each other by one SNP. Analysis of these SNPs revealed that the VSE isolate, but not the VRE isolates, encoded a mutation in the glucose importer glcU.
A nonsynonymous SNP at amino acid position 43 was detected that caused a Leucine residue to mutate to a stop codon. When we measured the biofilm formation of each isolate (Fig. 2), we found that the median OD550 of VSEfm15 was 0.12 (interquartile range [IQR], 0.11–0.13); in contrast the median OD550 of either of the VRE isolates was significantly greater: VREfm7 0.21 (IQR, 0.19–0.22) and VREfm10 0.20 (IQR, 0.18–0.22); ANOVA p < 0.0001; t-test p < 0.0001). The median OD550 of the two VREfm biofilms did not differ (p = 0.66).
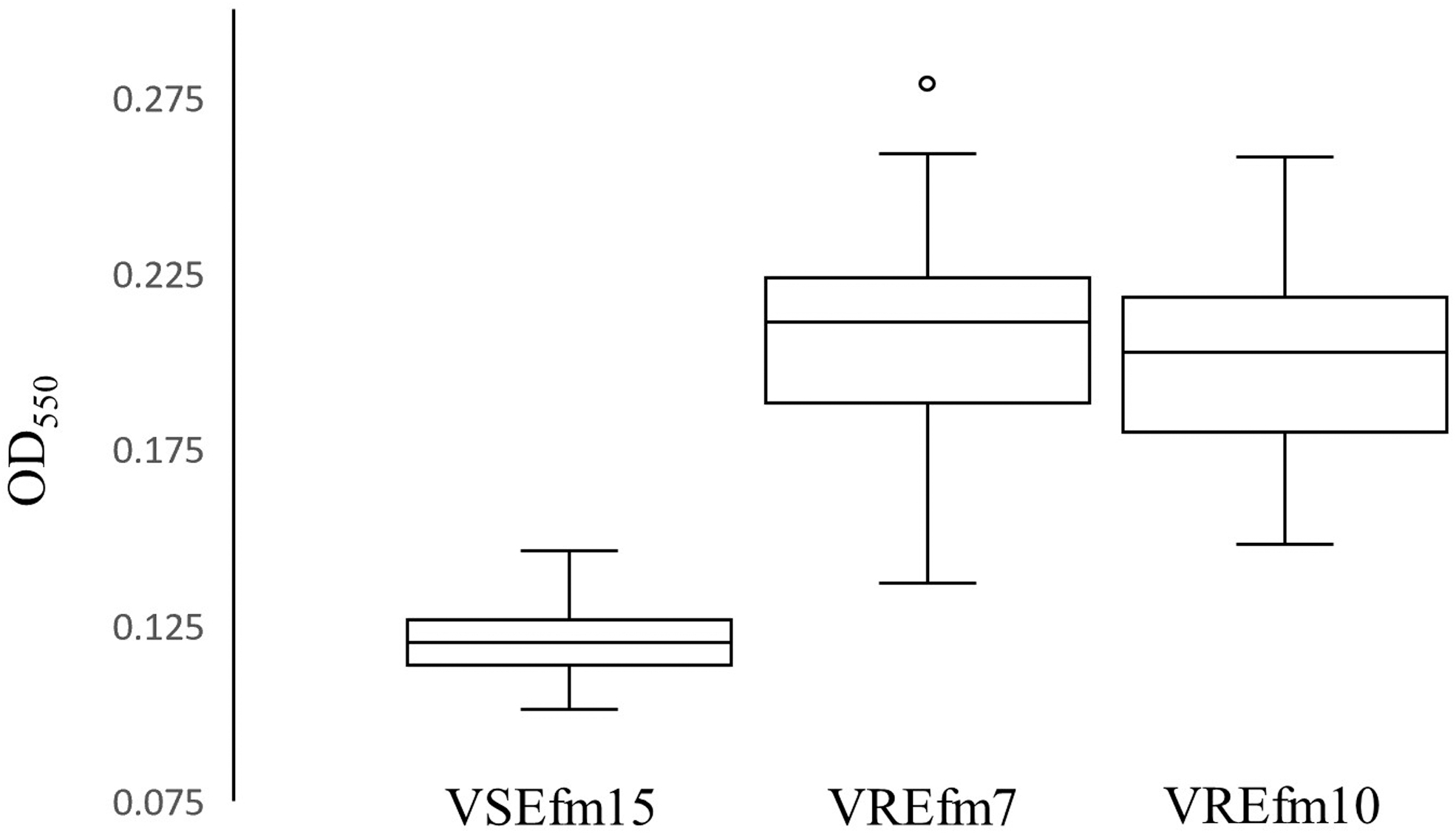
Biofilm formation of VSEfm15, VREfm7, and VREfm10 isolates. VREfm10 biofilm with one data point not shown at 0.34. VSEfm15 versus VREfm7, p < 0.0001. VSEfm15 versus VREfm10, p < 0.0001. VREfm7 versus VREfm10, p = 0.66. OD550—biofilm density550.
Discussion
In this case report, we describe a patient with four episodes of enterococcal bacteremia spanning a 6-month time period. In September 2020, the patient developed a VSEfm bacteremia associated with endocarditis and psoas abscess formation. After clearance of the VSEfm bacteremia with intravenous vancomycin and gentamicin, cardiac surgery revealed pacemaker lead vegetations and tricuspid valve endocarditis due to VREfm.
Genetic analyses using WGS revealed that the VREfm sampled in October was closely related to the VSEfm isolate collected in September, with the VREfm isolates acquiring vancomycin resistance through a plasmid. A deeper understanding of the different plasmids present in the VREfm strains could have been achieved through nanopore sequencing and hybrid assembly, however, we are unable to make such comparisons with Illumina short read sequence data alone. 14
The exact origin of this VanA operon-encoding plasmid is unknown; although no additional isolates were available for analysis, we suspect that it was likely acquired from a VREfm residing on the patient's skin or in the gastrointestinal flora, as the patient had enterococcal bacteremia due to a VREfm isolate in April 2020. It is less likely that the VSEfm isolate was a VVE strain, as no partial Van operon was detected in the VSEfm isolate genome, and the bacteremia cleared on vancomycin. Unfortunately, the isolate detected on the heart valve was not saved for further analyses, and thus we were unable to investigate this further.
With one exception, the VREfm and VSEfm isolates did not markedly differ in the presence of virulence genes or genes often associated with biofilm formation. In contrast, the VSEfm isolate, but not the VRE isolates, had an altered glucose transporter gene, which may have caused it to grow poorly in vitro. Thus, variation in biofilm formation may be due to alterations in this gene in addition to other unknown factors.
We hypothesize that VREfm did not cause the initial bacteremia given the fitness cost associated with vancomycin resistance. 3 Rather, the VREfm strain may have remained protected in the valve and pacemaker lead as a biofilm instead; the disruption caused by the cardiac surgery subsequently led to the VREfm bacteremia. This theory is supported by the greater biofilm formation of the VREfm isolates compared with the VSEfm isolate.
This case has implications for the diagnostic evaluation of patients with enterococcal endocarditis. Enterococci readily exchange genetic material through plasmid acquisition and the transfer of other mobile genetic elements. Thus, bacteria detected in the bloodstream may not accurately reflect organisms that have adhered to cardiac valves or in-dwelling devices, such as a pacemaker lead. In an effort to prevent treatment failure due to enterococci protected within biofilms, all tissue removed during cardiac surgery should be cultured; moreover, obtaining postoperative surveillance blood cultures should also be considered. In this way, patients can be assured that their antibiotic regimen is active against all bacterial strains causing infection.
Footnotes
Acknowledgment
We gratefully thank the Microbial Genomic Sequence (MiGS) Center for assistance with whole genome sequence data collection.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by funding from the University of Pittsburgh Department of Medicine (DVT).