
Abstract
An unusual biotype of KPC-2-producing Klebsiella pneumoniae (KPC-Kpn) isolates was detected in Corrientes, Argentina, which, to their isolation date, had been free of KPC-Kpn outbreaks. Our aim was to describe the clinical epidemiology focused on genomic characterization of atypical urease-negative KPC-Kpn clinical isolates belonging to the high-risk hospital-associated clonal lineage ST340/CC258. Thirteen isolates were recovered, all of them from inpatients with KPC-Kpn infection (August 2015 to January 2016). These isolates displayed identical enterobacterial repetitive intergenic consensus-PCR electropherotype belonging to a single clonal sequence type ST340. Whole genome sequencing was performed on two KPC-Kpn and the resistome analyses revealed the following acquired resistance genes: blaKPC-2, blaCTX-M-15, blaOXA-1, blaSHV-11, aac(3)-IId, aph(3′)-Ia, aac(6′)-Ib-cr, sul1, dfrA14, catB3, fosA, and arr-3. Mutations in GyrA (S83I) and ParC (S80I) were also identified. Among the virulence determinants, yersiniabactin was detected in both strains, specifically the ybt9 locus located in ICEKp3. Five plasmid incompatibility groups were observed in this clone and an unusual IncP6 plasmid bearing blaKPC-2 gene (named pKpn3KP) was fully characterized. In this study, we present the first draft genome sequences of two clinical isolates of KPC-2/CTX-M-15-producing K. pneumoniae belonging to the high-risk clonal lineage ST340/CC258 associated with nosocomial outbreaks in Argentina.
Introduction
Multidrug-resistant and extensively drug-resistant Klebsiella pneumoniae are an important cause of hospital-acquired and difficult-to-treat infections. 1
Prevalence of KPC-2-producing K. pneumonia (KPC-Kpn) belonging to international high-risk clones has been driven both by clonal dissemination of KPC-producing K. pneumoniae and the mobility of this gene. 2 Dissemination of hyperepidemic ST258, as well as ST11, ST25, ST392, and ST307 clones have already been reported in Argentina.2–4 IncF, IncN, IncA/C, and IncX were the most common replicon types associated to the blaKPC-2 harboring plasmids described in these STs, although nontypeable replicons have previously been mentioned. 5
An unusual biotype of KPC-Kpn isolates was detected in 2015 in the province of Corrientes, which had been free of KPC-Kpn outbreaks up to that time. This province shares borders with Paraguay and Brazil and is key for the regional transit relationship with these two countries.
The original KPC-producing isolate was obtained from a patient that was referred from Chaco, a neighboring province of Corrientes (Supplementary Fig. S1), in August 2015. The isolate was characterized as Klebsiella (Raultella) terrigena by API 20E (98%) from a urine colonization. Subsequently, the isolate was subjected to the semiautomated Vitek 2 system and matrix assisted laser desorption ionization-time of flight confirmation, which identified it as K. pneumoniae, displaying an atypical negative urease production result. After that, other 12 microorganisms (from 11 different patients) with the same profile were recovered in two nearby hospitals. No similar microorganisms were recovered after January 2016.
Four microorganisms were isolated from rectal swabs, and eight from clinically relevant samples (three from urinary tract infections, two from a respiratory syndrome, one from a hepatic abscess, one from a pancreatic abscess, and one from a mediastinitis case); excluding surveillance cultures and urine samples, four out of five patients died (Supplementary Fig. S2).
In general, antimicrobial susceptibility was performed by disk diffusion, and minimal inhibitory concentration (MIC) for meropenem and colistin was determined by broth microdilution according to CLSI 6 guidelines.
All these isolates showed the same biotype and a multidrug resistance profile displaying resistance to piperacillin/tazobactam, cefotaxime, ceftriaxone, ceftazidime, cefepime, ertapenem, imipenem, meropenem (MIC range = 4–8 μg/mL), ciprofloxacin, trimethoprim/sulfamethoxazole, and gentamicin, being only susceptible to amikacin and colistin (MIC = 0.125 μg/mL). Screening for carbapenemases was always positive by using phenylboronic acid, disks, modified Hodge and blue-Carba tests. 7 The blaKPC-2 gene was confirmed by PCR and sequencing. All of them displayed an indistinguishable repetitive element palindromic/enterobacterial repetitive intergenic consensus-PCR electropherotype 8 (Supplementary Fig. S3) and could be assigned to ST340 when multilocus sequence typing (MLST) was performed by PCR and sequencing using the K. pneumoniae MLST database. For this reason, a further analysis was performed on only two isolates.
Plasmid conjugation was performed by a mating-out assay using Escherichia coli CAG 12177 (tetracycline resistant) and E. coli J53 (sodium azide resistant) as recipients, and Luria Bertani agar plates supplemented with tetracycline (20 mg/L) or sodium azide (150 mg/mL) and imipenem (1 mg/mL) as selective agents. No transfer of blaKPC-2 was obtained by conjugation from K. pneumoniae Kpn 2KP (mediastinitis) and Kpn 3KP (rectal swab), suggesting that this marker was not located on a conjugative plasmid.
Total genomic DNA of strains Kpn 2KP and Kpn 3KP was sequenced using an Illumina NextSeq sequencing platform (Illumina, Inc., San Diego, CA), which produced an average of 1,272,635 paired-end reads (SD 57,813) with a 98 × total coverage. De novo assembly was performed using the SPAdes V3.9 and analyzed by using online bioinformatic tools (Center for Genomic Epidemiology, CARD and Pathogenwatch). The nucleotide sequences (genome assemblies and reads) were uploaded to GenBank under BioProject accession no. PRJNA780904. Data related to G + C content, number of contigs, genome size, and N50 parameter are summarized in Table 1.
Genomic Characteristics of Kpn 2KP and Kpn 3KP Isolates
MLST, multilocus sequence typing.
Automated annotation was done with Prokka v.1.14.6 and manually curated using ISFinder and Uniprot databases, respectively, resulting in 5,189 and 5,121 coding sequences (CDS) for Kpn 2KP and Kpn 3KP strains, respectively. Schematic representation and comparative analysis of plasmids were performed using Proksee.
MLST, antimicrobial resistance genes (ARGs), virulence factors, plasmid replicons, capsule synthesis (K), and lipopolysaccharide (O) loci were identified using bioinformatics software/tools and are summarized in Table 1.
MLST using MLST 2.0 confirmed both isolates as ST340. Resistomes that were analyzed using ResFinder 4.1 revealed identical resistance genes: aac(3)-IId, aph(3′)-Ia, aac(6′)-Ib-cr, blaKPC-2, blaCTX-M-15, blaOXA-1, blaSHV-11, sul1, dfrA14, catB3, mph(A), fosA-like, arr-3, and aac(6′)-Ib-cr, oqxA, oqxB, and substitutions in GyrA (S83I) and ParC (S80I).
Five plasmid incompatibility groups [IncFIB(K), IncFII(K), IncP6, IncR, and IncFIA(HI1)] were identified by PlasmidFinder 2.1. Klebsiella-specific virulence determinants were investigated using Kleborate and only yersiniabactin was detected in both strains, with the ybt9 locus located in ICEKp3 (an integrative conjugative element that mobilizes the ybt locus, which encodes biosynthesis of the siderophore yersiniabactin and its receptor). This ICEkp3 is one of the most widely distributed variants reported in phylogenetically distinct K. pneumoniae lineages. 9
Both Kpn 2KP and Kpn 3KP harbored blaKPC-2 in an IncP6 plasmid, a broad-spectrum plasmid sporadically reported worldwide 10 and never reported before in clinical isolates from Argentina. However, two IncP6 plasmids were recently described in Argentina in nonclinical KPC-2-producing Klebsiella quasipneumoniae and Enterobacter asburiae isolated from wastewater samples in 2018. 11 The full sequence of the blaKPC-2-harboring plasmid (pKpn3KP, GenBank JAJKGK010000036) was 38,573 bp in size and showed that blaKPC-2 was flanked upstream by a ΔISKpn6 and downstream by a truncated blaTEM-1 (ΔblaTEM-1), ISKpn27 and a Tn3 family transposon.
This plasmid did not harbor any further resistance genes and was practically identical (100% coverage with 99% identity) to pKOX3-P5-KPC and pWW14A-KPC2, previously reported in clinical Klebsiella oxytoca in China (GenBank KY913901.1) and in K. quasipneumoniae recovered from wastewater in Argentina (GenBank CP080103.1), respectively. Nevertheless, a 1,702-bp region containing the protecting, antioxidant MsrA/B system genes was absent in pKpn3Kp (Fig. 1). Both plasmids lack a tra module encoding primary pilus, which explains the failure of plasmid conjugation.
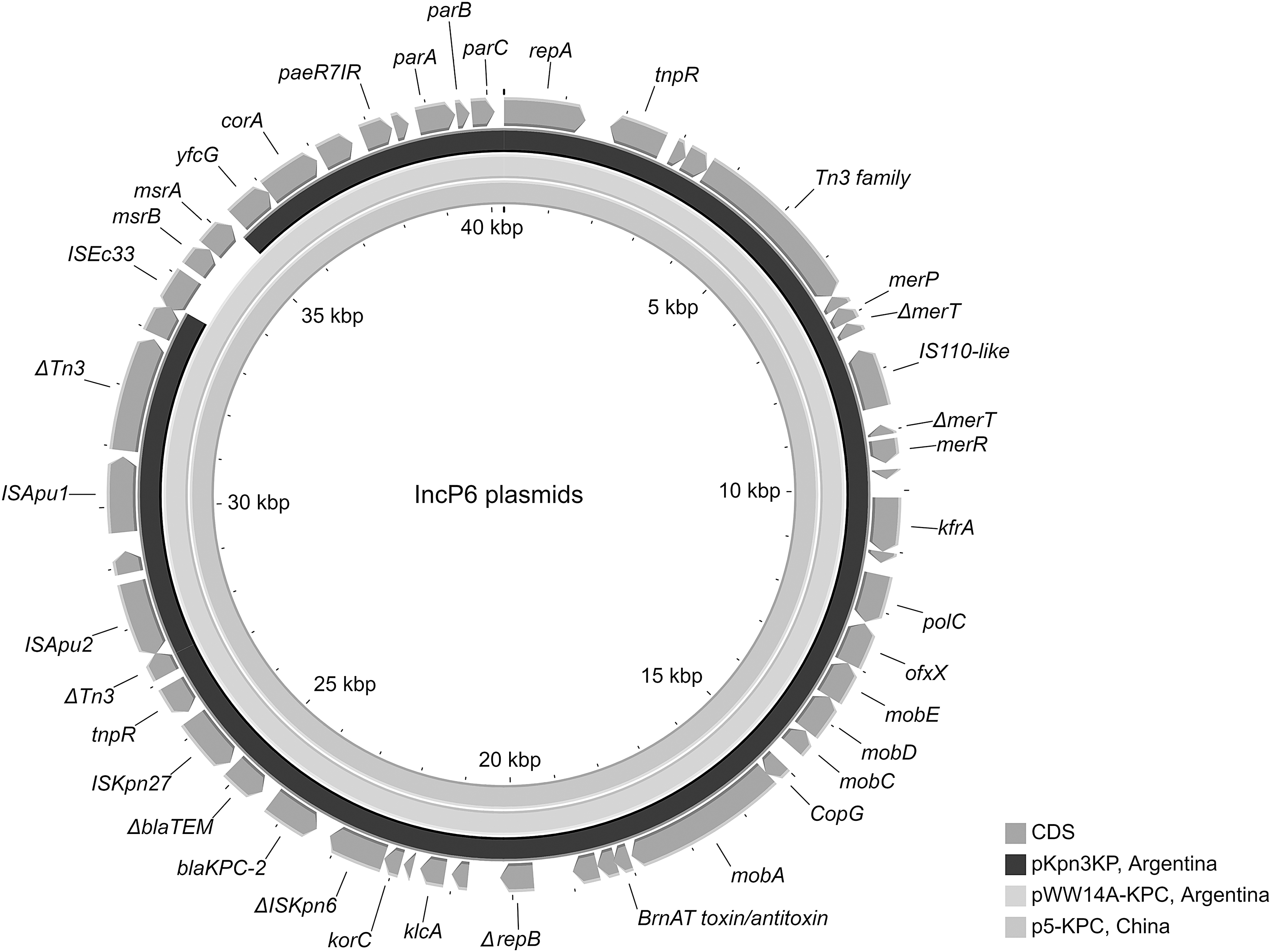
Comparison of pKpn3KP with IncP6 plasmids available in public databases. The analysis showed that the pKpn3KP structure was highly similar to blaKPC-2-containing plasmids of both environmental and clinical origin deposited in GenBank. Both pWW14A-KPC2 from Klebsiella quasipneumoniae recovered from wastewater (GenBank accession no. CP080103.1; Argentina) and p5-KPC_PKOX3 from clinical Klebsiella oxytoca (GenBank accession no. KY913901.1; China) displayed 100% query coverage and 99% nucleotide identity.
Even after different assembly methods were attempted, the blaCTX-M-15 contig (1,425 bp in length) was not long enough to predict its location. After annotation, the contig was found to carry only blaCTX-M-15 and the wbuC gene encoding a cupin fold metalloprotein, commonly found flanking blaCTX-M-15 located on both chromosomal and plasmid sequences.
Comparative analyses were performed to understand the urease-negative phenotype in these isolates. A complete urtABCDE operon was found in both sequences, whereas the single-component urea transporter, Yut, was absent. Of note, in the urease (EC 3.5.1.5) operon (ureABCDEFG) ureC, which encodes the alpha subunit, a 1327-bp fused IS5075 (an IS110 family) was found. These findings may explain the experimental evidence on the lack of ureolytic activity, and, as far as we know, no explanation for this phenotype in K. pneumoniae has been previously given.
The genetic relationship based on single nucleotide polymorphisms (SNPs) was assessed using genomic sequences of K. pneumoniae ST340 available at Pathogenwatch (July 2021). SNPs were identified and extracted using SNP-sites v.2.5.1 and a maximum-likelihood clustering was inferred by IQ-TREE Phylogenomic v.1.5.5.3 (1,000 bootstraps). The tree was visualized using Microreact. Clustering based on core genome SNPs has shown the evolutionary relatedness among 117 K. pneumoniae ST340 isolates worldwide (including strains Kpn 2KP and Kpn 3KP), and the accompanying ARGs were also highlighted. The same substitutions were detected in all ST340 isolates in the quinolone resistance-determining region (GyrA S83I and ParC S80I), being the most frequent alterations in clinical isolates. 12
The phylogeny of the ST340 genomes showed that KPC-producing isolates (32.5%, 38/117) were found in clades arising from two main deep branches of the tree (Supplementary Fig. S4a).
All KPC-3- (n = 22) and KPC-4- (n = 5) producing strains were recovered from the United States (yellow marks), and clustered into the same clade I, whereas most of KPC-2 producers (10/11), including the isolates in this study, had geographically distinct origins [Argentina (pink marks), Brazil (red marks), and non-American countries (blue marks)] and were grouped into Clade II. Noteworthy, most of the European and other non-American ST340 genomes not associated to blaKPC (blue marks) also belonged to the same Clade II, most of which harbored blaCTX-M-15.
K. pneumoniae Kpn 2KP and Kpn 3KP were closely grouped with six other KPC-2- and/or CTX-M-15-producing strains ST340 within Clade II, two of which were collected in Brazil from human hosts (Supplementary Fig. S4b).
K. pneumoniae Kpn 2KP and Kpn 3KP showed 158 SNPs of difference. Interestingly, none of the 115 K. pneumoniae ST340 isolates compared in this study carried an IncP6 plasmid.
These KPC-2-producing isolates with an atypical urease reaction were associated with nosocomial outbreaks. The initial misidentification of these microorganisms by commercial panels may pose a potential risk due to the lack of an active outbreak policy.
These are the first draft genome sequences of clinical isolates of KPC-2/CTX-M-15-producing K. pneumoniae belonging to the high-risk clonal lineage ST340/CC258 associated with nosocomial outbreaks in Argentina, providing an early warning of the emergence of an unusual clone; this is particularly relevant considering that CTX-M-15- (and/or KPC-2-) producing K. pneumoniae ST340 have already been reported in clinical isolates,13,14 companion animals, 15 and environmental samples 16 from Brazil. Excluding surveillance and urine samples, four of the five patients died within the study period.
Last but not least, it is important to note that an unusual IncP6 plasmid bearing the blaKPC-2 gene was detected in this clone, which is already circulating in isolates from different niches in Argentina. 11
Footnotes
Acknowledgments
J.D.C. and G.G. are members of CONICET.
Authors' Contributions
J.D.C. and G.G. conceived and designed the experiments, analyzed the results and wrote the article. M.E.B., Y.C., and L.P. supplied the clinical isolate and performed the phenotypical characterization. N.L. and H.F. performed WGS and analyzed the results obtained. All authors reviewed and edited the article.
Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This study was supported by Agencia Nacional de Promoción Científica y Tecnológica PICT 2019-01879 to JDC, PICT 2015-1925 and UBACyT 2018- 20020170100473BA to GG and Conselho Nacional de Desenvolvimento Científico e Tecnológico (grants AMR 443819/2018-1, 433128/2018-6, and 312249/2017-9).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.