
Abstract
Typing carbapenem-resistant Klebsiella pneumoniae (CR-KPN) is crucial in controlling their dissemination and solving outbreaks. In this context, we searched for an effective, faster, and cheaper alternative technique to type KPN by analyzing the fosAKP sequence. We analyzed the nucleotide sequences of chromosomal fosAKP gene in 350 KPN genomes (70 per sequence type [ST] or clonal complex [CC]). Assembly genomes were randomly downloaded from NCBI and annotated using RAST in PATRIC platform. The isolate STs were verified using multilocus sequence typing 2.0 by the Center for Genomic Epidemiology. Chromosomally encoded fosAKP was confirmed in MLplasmid, and the sequence alignments were performed in Clustal Omega. The amino acid sequences were analyzed using SNAP2 and SMART platforms. Out of the 70 genomes analyzed for each ST/CC, we observed 100% fosA sequence identity for CC258/11, ST15, ST307, and ST101. For ST16, only two fosA sequences were different from each other. We observed differences in amino acid sequences at positions 25 and 79 (ST16) and 86 (ST16, ST101). The C-terminal (amino acid 138, 139, 140) was different for each cluster. None of these polymorphisms is related to the protein active site. Moreover, L25Q (ST16) polymorphism was predicted to probably affect the protein function. We observed that chromosomal fosAKP sequences from KPN are highly conserved in ST15, ST307, ST16, ST101, and CC258/11, suggesting fosAKP sequencing as an alternative, easier, faster, and less expensive technique in identifying epidemiological STs for KPN, and discriminating them from CC258/11.
Introduction
Klebsiella pneumoniae (KPN) is a ubiquitous enterobacterium found in animals, humans, and the environment. Besides being a member of the human microbiota, this species can be pathogenic and cause difficult-to-treat infections. 1 Carbapenem-resistant K. pneumoniae (CR-KPN) has been recognized as a major public health problem because it has been associated with high morbidity and mortality rates, especially due to carbapenemase production. 2 Currently, the multilocus sequence typing (MLST) technique is used to get a comprehensive understanding of the global CR-KPN epidemiology. The global dissemination of K. pneumoniae carbapenemase (KPC) has been attributed to specific clones of KPN such as clonal complex (CC) 258/11, sequence type (ST) 16, ST15, and ST101, with ST258 being the most prevalent worldwide.2,3
In the last years, ST307 has gained importance because it is widely spread around the world and is associated with the emergence of ceftazidime/avibactam resistance. 4 In addition, KPN ST11 clones carrying the blaOXA−48 gene have been frequently reported in Spain, Argentina, and Turkey. In contrast, blaOXA −48 was observed in KPN ST15 and ST101 in the United States, Spain, Czech Republic, Libya, India, Germany, Finland, France, and Japan. 3 Reports from various countries in Asia and Europe showed that KPN ST11 clones also harbored the blaNDM gene. 1 blaNDM was also found in KPN ST340 in Spain, Serbia, and Korea, and in ST437 in Singapore, Pakistan, and Spain.1,2 Although KPC-2-producing KPN CC258/11 was endemic in many Brazilian hospitals before the COVID-19 pandemic, the emergence of “new” STs was also reported.3,5 Furthermore, KPC-2 producing KPN ST16 was associated with a higher virulence and 30-day mortality rate. 3
Although whole-genome sequence (WGS) has been widely applied for outbreak investigations worldwide, the turnaround time for results availability is sometimes long. In addition, this methodology is expensive for most institutions in low- and middle-income countries (LMIC). For this reason, it would be highly desirable to have an easier, faster, and less expensive alternative technique to easily discriminate new emerging STs. In fact, during the COVID-19 pandemic, an alarming increase in New Delhi metallo-β-lactamase (NDM)-producing KPN was detected in many Latin American countries, including Brazil.6,7
FosAKP is a chromosomally encoded glutathione transferase that inactivates fosfomycin in KPN. 8 The fosAKP gene is located downstream of the transcriptional regulator lysR gene, overlapping its end. 9 Furthermore, fosAKP can be mobilized by mobile genetic elements to other species; especially Escherichia coli, leading to fosfomycin resistance. 8 In the present study, we evaluated the sequencing of fosAKP as an alternative for discriminating KPN CC258/11 from other STs.
Materials and Methods
Bacterial isolates and genome analysis
A total of 23 KPC-2-producing KPN strains belonging to 6 different STs were previously characterized. 3 During that analysis, it was noticed that KPN not belonging to the CC258/11 had distinct fosA sequences. To evaluate the correlation between fosA sequence and specific STs, we included other 327 KPN genomes obtained from the Pathosystems Resource Integration Center (PATRIC) and National Center for Biotechnology Information (NCBI) data banks in our analysis, summing-up to 350 KPN genomes (70 KPN strains each from CC258/11, ST15, ST16, ST101, and ST307).
The 280 KPN genomes were annotated using RAST in PATRIC platform, while the sequence of fosAKP gene was obtained in fasta format and the sequences were then compared by nucleotide sequence alignment using Clustal Omega and MView. Furthermore, the ST of each strain was confirmed using MLST 2.0 by the Center for Genomic Epidemiology. Supplementary Table S1 shows the accession numbers of the strains included in this study. The chromosomal location of fosAKP was confirmed by MLplasmid® software. The SNAP2 software was used to estimate the functional effects caused by amino acid changes. In addition, the protein domain was searched by SMART platform.
Results
fosA gene nucleotide sequence analysis
After performing the nucleotide and amino acid sequence alignment of fosA and FosA, respectively, for the first 23 KPN strains, 4 clusters were identified, and all the strains belonging to a specific cluster shared identical fosA nucleotide and FosA amino acid sequences. Interestingly, each cluster corresponded to an ST or CC, being divided into CC258/11, ST16, ST15, and ST101.
By enlarging the number of KPN representatives, more genomes were downloaded, annotated, and analyzed, amounting to 70 strains in each cluster including ST307, an emergent ST around the world. The alignment among the fosAKP sequences of each ST/CC is shown in Fig. 1. The percentage of fosAKP identity among the STs obtained by fosAKP sequence analysis of each ST and the distance between each cluster are depicted in Fig. 2. For CC258/11 represented by isolates of ST258, ST11, ST340, and ST437, the 70 fosAKP gene and FosA protein sequences shared 100% identity, and the same was observed for ST15, ST101, and ST307. Only 2 (2.8%) out of the 70 KPN ST16 strains showed different fosAKP sequences, sharing 95.0% nucleotide and 96.4% amino acid identities.

Alignment between the STs. The nucleotides in the boxes are those presenting mutations in at least one of the sequences. ST, sequence type.
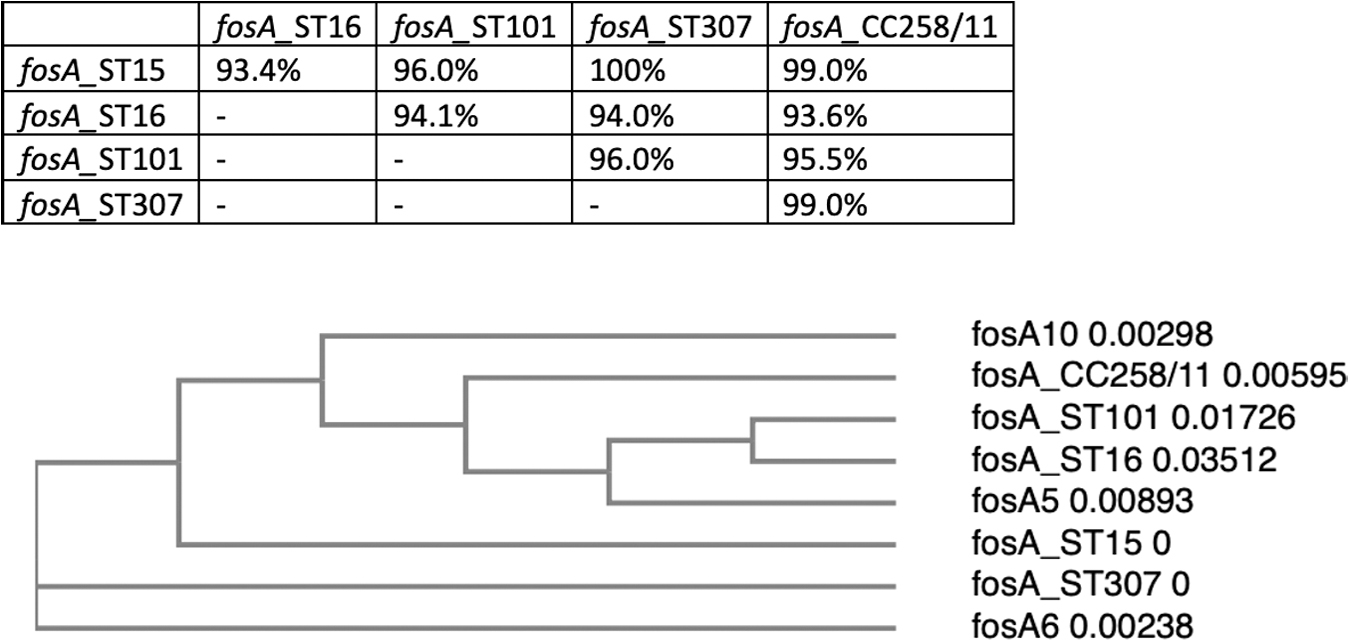
Phylogram comparing the fosA gene from the different clusters and fosA5, fosA6, fosA10 and the identity percentages among STs/CCs. CCs, clonal complexes.
These two sequences shared 100% identity with fosAKP genes from KPN ST17, a closely related ST. Furthermore, we observed that fosAKP identity between ST15 and ST307 was 100% (Fig. 2).
FosA protein amino acid sequence analysis
After performing the FosA alignment from the different STs and the previously reported FosA variants recognized to have originated from KPN (FosA5, FosA6, and FosA10),10–12 we observed the presence of 139 amino acids in FosA from ST15, ST101, ST307, and CC258/11, while FosA from ST16 presented 140 amino acids. Furthermore, we found amino acid polymorphisms as L25Q and S79R changes in ST16, and A86V change in ST16 and ST101. At position 91, the CC258/11, ST16, and ST101 showed the amino acid valine (V), while ST15 and ST307 had an isoleucine (I) such as FosA6 and FosA10. It is important to highlight that none of these polymorphisms was within the protein active site, 13 but in the glyoxalase protein domain, according to Pfam 34.0. 14 Also, the final portion of the amino acid sequences (residues 138, 139, and 140) varied (Fig. 3).

Alignment among the FosA protein of each cluster and FosA5, FosA6, and FosA10. The arrows indicate the residues of the protein active site. In gray indicates the glyoxalase domain. The (*) indicates the conserved amino acids among the different FosA sequences.
Moreover, a single polymorphism was predicted by SNAP2 to probably affect the protein function: the FosA from ST16 presented the amino acid glutamine instead of leucine (L25Q), as presented by ST15, ST101, CC258/11, ST307, FosA5, FosA6, and FosA10.
Discussion
Recent studies have shown increasing rates of CR-KPN worldwide. 15 Although CR-KPN CC258/11 has been responsible for the worldwide spread of KPC, emerging STs have been reported to cause novel outbreaks.3,16 In addition, other CR-KPN STs carrying blaNDM have been observed.5,17 Furthermore, there is evidence that different STs or CCs vary across distinct geographical regions, raising the need for better CR-KPN characterization.15,18,19 In this manner, it is important to rapidly and precisely identify CR-KPN to discriminate outbreaks from those caused by CR-KPN CC258/11, as well as to identify clinical outbreaks, prevent their spread, and evaluate possible differences in clinical outcomes.1,2
Despite recent advances in WGS, MLST has been a technique commonly used for microbiological characterization of KPN isolates. It involves sequencing of seven different housekeeping genes: gapA, infB, mdh, pgi, phoE, rpoB, and tonB. In addition to being laborious, MLST can cost between $25 and $60 per isolate in Brazil, which is expensive for most clinical settings, especially in LMIC. In this manner, alternative typing methods to identify CR-KPN clones have been searched. Pena et al recently reported the use of matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) as an alternative way to type ST11 and ST101 CR-KPN, thus reducing the costs and turnaround time required for typing results. 20
In the present work, we analyzed fosAKP sequence, a chromosomally encoded gene in K. pneumoniae, as an alternative typing method to distinguish CC258/11 from other STs. We observed that the fosAKP sequence was highly conserved in ST15, ST16, ST101, ST307, and CC258/11, indicating that fosAKP could be a surrogate marker to identify distinct STs associated with outbreaks. In contrast, it was not possible to differentiate between ST15 and ST307 because they share the same fosAKP sequence. However, the specific fosAKP sequence found in ST15 and ST307 could early warn about the possible emergence of these two important STs. ST307 is an emergent and important K. pneumoniae ST related to the emergence of ceftazidime/avibactam resistance, 4 and ST 15 carries a recently identified new adhesion system, Kpi. This adhesion system favors the strain persistence in the nosocomial environment. 21
By analyzing the FosA sequences from ST15, ST16, ST101, ST307, and CC258/11, we observed that the polymorphisms in the amino acid sequences of FosA were not associated with residues of protein active site, possibly indicating no changes in the interactions between FosA and its ligands. Klontz et al studied the crystallized FosAKP structure and amino acid sequence, and reported residues H7, H67, E98, and E113 as important to the interaction with the divalent cation; T9, K93, S97, S101, Y103, and R122 as coordinating the fosfomycin binding; and Y65 connecting the active site and the dimer loop. 13 These residues are not related to the polymorphisms observed in the present study.
Among the polymorphisms observed, only the L25Q in FosAKP from ST16 is supposed to affect the protein function. Even though this change is not related to the protein active site, the replacement of a charged for an uncharged amino acid could alter the protein function and consequently its efficiency. 22
In this study, we demonstrated that the fosAKP sequencing is a promising surrogate marker to discriminate CR-KPN CC258/11 from other clones. By sequencing only fosAKP, the amount of work could be significantly reduced as well as the amount spent for this purpose: from $25–$60 to $3–6 per isolate. In addition, the time spent to achieve the isolate's ST would be shorter, resulting in faster introduction of controlling measures to limit the CR-KPN dissemination.
It is important to highlight that even fosA is a fosfomycin resistance gene and typing bacteria through resistance genes is not usual, a previous study has already been successful using resistance genes to type bacteria. 23
However, our study has limitations. We included in our analysis the most frequent STs reported in high-risk KPN clones, but the number of STs evaluated was low and could be further expanded in future studies. Furthermore, although chromosomal fosAKP gene capture from KPN by plasmids is possible, to the best of our knowledge, no report of plasmid-encoded fosAKP (or its variants fosA5/fosA6/fosA10) in this species has been documented so far. Furthermore, when located in plasmids, the fosA regulator, lysR gene, is usually truncated by an insertion sequence, whereas in the chromosome, the lysR is intact.
Thus, we could solve this doubt; designing primers targeting the lysR-fosA sequence and based on the size of the amplicon suggests the fosA location (plasmid or chromosome). Also, it is important to highlight that the usually found plasmid-enconded fosA in KPN is the fosA3 variant, which can be easily discriminated from fosAKP by polymerase chain reaction 9 or sequencing, the technique chosen in the present study.
Finally, chromosomally encoded fosA has also been reported in other bacterial species. Ito et al identified high frequencies of chromosomal fosA gene in Providencia stuartii (100%), Serratia marcescens (99.7%), Pseudomonas aeruginosa (98.8%), Klebsiella aerogenes (98.4%), Klebsiella oxytoca (96.6%), and Morganella morganii (90.5%). 8 However, more investigation is needed to fully understand and confirm the chromosomal location of fosA genes in these species and to verify if the fosA sequence could also be applied for typing.
Therefore, new studies focusing on the analysis of fosA gene sequences are needed to confirm our findings and the roles of fosA as an alternative typing tool.
Conclusions
Our study showed that fosAKP is a highly conserved gene in K. pneumoniae and its nucleotide sequence could be used as an alternative technique to discriminate CR-KPN CC258/11 clones from other clones, as well as identifying clones from ST16, ST15, ST307, and ST101. Thus, by becoming available, this alternative technique would be easier, faster, and less expensive, especially for LMIC, for typing high-risk K. pneumoniae clones.
Footnotes
Acknowledgments
We would like to thank the researchers: Dr. Willames M.B.S. Martins and Dr. Diego O. Andrey for providing the genome sequencing data. Also, we are grateful to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for providing scholarships to Á.C.d.S.R. (PDS), F.F.S. (PNPD), and I.B.M. (Young Talent).
Authors' Contributions
Á.C.d.S.R.: conceptualization, data curation, formal analysis, investigation, methodology, visualization, and writing—original draft.
F.F.d.S.: conceptualization, investigation, methodology, and writing—original draft.
I.B.M.: writing—review and editing.
L.A.d.R.M.: writing—review and editing.
A.C.G.: writing—review and editing, supervision, resources, and project administration.
Disclosure Statement
No competing financial interests exist.
Funding Information
Scholarships funding by CAPES.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.