
Abstract
Antisense oligonucleotides (AOs) can facilitate the expression of internally deleted dystrophin in dystrophin-deficient Duchenne muscular dystrophy (DMD) by correcting the reading frame of the pre-mRNA with AO-mediated exon skipping. An antisense 18-mer 2′-O-methyl RNA/ethylene-bridged nucleic acid chimera AO targeting exon 45 of the dystrophin gene, AO85, can induce exon 45 skipping efficiently in cultured cells. AO85 is expected to facilitate dystrophin expression in 8%–9% of all DMD patients. Here, we examined the kinetics of AO85-mediated exon 45 skipping in a cell-free splicing system. In vitro transcribed pre-mRNAs containing dystrophin exon 45 and part of its flanking introns within a hybrid minigene were incubated with HeLa cell nuclear extract, and the resultant mRNAs were amplified by semiquantitative reverse transcriptase–polymerase chain reaction. Time-course analysis revealed that the splicing process fitted well to first order kinetics. Addition of AO85 produced an extra spliced product, deleting exon 45 (Δexon 45), indicating AO85-mediated exon 45 skipping. Production of Δexon 45 increased linearly with increasing concentrations of AO85, reaching a maximum of nearly 80% of the transcripts. The half-maximal effective concentration (EC50) of AO85 was 58.0 nM. The percentage of Δexon 45 among the transcripts decreased inversely with the pre-mRNA concentration; Lineweaver-Burk plotting revealed a competitive fashion of AO85 action. The low EC50 indicates high potential of AO85 for clinical application.
Introduction
Duchenne muscular dystrophy (DMD) is a rapidly progressive muscle wasting disease that usually results in death during the third decade. DMD is characterized by a dystrophin deficiency that usually arises from out-of-frame deletion mutations in the dystrophin gene that create a premature stop codon in the dystrophin mRNA. AO-mediated exon skipping therapy has been proposed for the treatment of DMD, producing in-frame dystrophin mRNA from the out-of-frame mRNA by inducing exon skipping. The newly generated in-frame dystrophin mRNA is expected to produce semifunctional, internally deleted dystrophin protein (Takeshima et al., 1995). The production of internally deleted dystrophin has been clinically demonstrated in some patients who showed natural skipping of an exon encoding a nonsense mutation (Shiga et al., 1997; Flanigan et al., 2011). Compared with gene replacement therapy, the benefits of exon skipping with AOs include the ability to use the endogenous gene and easy chemical synthesis of AOs. Currently, induction of exon skipping with AOs is considered one of the most plausible treatments for DMD (AARTSMA-RUS, 2010; Lu et al., 2011). Remarkably, systemic administration of 2′-O-methyl phosphorothioate AO (PRO051) showed a promising result of a modest improvement in the 6 minutes walk test after 12 weeks of the treatment (Goemans et al., 2011).
AOs with phosphorothioate backbones directed against the splicing enhancer sequence within human dystrophin exon 19 provided the first demonstration of exon skipping in human cells (Pramono et al., 1996). Since then, intravenous infusion of AOs against exon 19 has been reported to induce exon 19 skipping and the expression of internally deleted dystrophin in an exon 20-deleted DMD patient (Takeshima et al., 2006). Several chemical modifications of the first-generation phosphorothioate AOs have been introduced over the past decade to improve the efficiency of antisense therapeutics. Morpholino AOs have been shown to induce dystrophin exon 51 skipping and promote dystrophin expression in DMD muscle cells (PARTRIDGE, 2010).
A modified nucleic acid, 2′-O,4′-C-ethylene-bridged nucleic acid (ENA), has high binding affinity for the complementary RNA strand and more nuclease resistance than unmodified nucleic acid (Morita et al., 2001; Veedu and Wengel, 2010). One antisense 2′-O-methyl RNA/ENA (RNA/ENA) chimera was shown to be 40 times more effective than the conventional phosphorothioate backbone oligonucleotides in inducing exon 19 skipping (Yagi et al., 2004). Furthermore, an RNA/ENA chimera against dystrophin exon 41 encoding a nonsense mutation has been shown to induce efficient skipping of a mutated exon 41 (Surono et al., 2004). Considering that skipping of exon 45 is expected to express internally deleted dystrophin in 8%–9% of DMD patients (Aartsma-Rus et al., 2009a; Takeshima et al., 2010), AOs that induce exon 45 skipping have been proposed as one of the most important AOs for DMD treatment. In our previous studies, we demonstrated that an 18-mer RNA/ENA chimera, AO85, strongly induced exon 45 skipping in cultured myocytes (Takagi et al., 2004a; Takeshima et al., 2011). However, its kinetics, including the half-maximal effective concentration (EC50), are still unknown, as for other AOs designed for DMD treatment (AARTSMA-RUS, 2010).
To facilitate the clinical application of AO85, here we examined AO-mediated exon 45 skipping in a cell-free splicing system and determined the splicing parameters of AO85, revealing a low EC50.
Materials and Methods
AO85
An 18-mer RNA/ENA chimera (5′-CgCTgcCCaaTgCCatCC-3′; upper and lower case letters represent ENA and 2′-O-methyl RNA, respectively), AO85, complementary to the 5′ region of dystrophin exon 45 (Takeshima et al., 2011), was synthesized by KNC Laboratories Co. (Kobe, Japan) and dissolved in water.
In vitro transcription
The region encompassing dystrophin exon 45 and part of its flanking introns was amplified from human genomic DNA by polymerase chain reaction (PCR) using a set of primers containing NheI and BamHI restriction enzyme recognition sites (forward primer, Int44FNheI 5′-
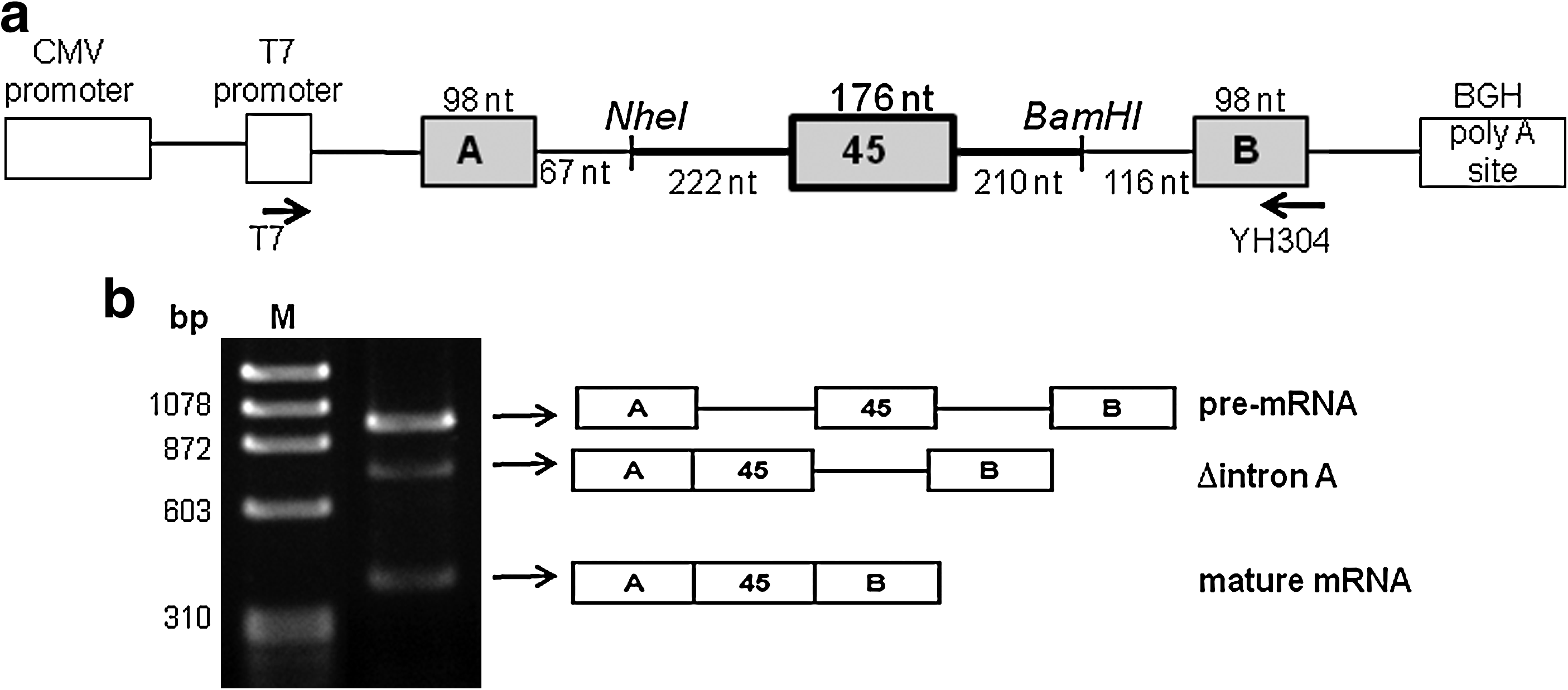
Cell-free splicing reaction.
In vitro splicing reaction
The in vitro splicing reaction was carried out at 30°C for 2 hours in a volume of 20 μL containing 30 ng pre-mRNA, 50% (v/v) HeLa cell nuclear extract (Lot No. 4168HNE; Computer Cell Culture Center, Mons, Belgium), 1.6 mM MgCl2, 0.5 mM ATP, 20 mM creatine phosphate, and 20 U of RNaseOUT (Invitrogen, Carlsbad, CA) (Habara et al., 2008). In some experiments, the indicated amounts of AO85 were added directly to the reaction mixture. The incubation temperature or time was varied to study the temperature or time dependency of the in vitro splicing.
After the designated incubation time, reaction mixtures were treated with Proteinase K (Sigma-Aldrich, St. Louis, MO) and the resulting RNAs were purified by phenol–chloroform extraction and ethanol precipitation as previously described (Habara et al., 2008). Purified total RNA (750 ng) was reverse transcribed using random hexamer primers with M-MLV reverse transcriptase (Invitrogen) according to the manufacturer's protocol and then PCR amplified using a forward primer from exon A (YH307, 5′-ATTACTCGCTCAGAAGCTGTGTTGC-3′) and a reverse primer from exon B (YH308, 5′-AAGTCTCTCACTTAGCAACTGGCAG-3′) (Habara et al., 2008). PCRs were performed in a volume of 20 μL containing 4 μL of cDNA, 1× Ex Taq Buffer, 250 nM dNTPs (Takara Bio, Inc., Kyoto, Japan), 10 pmol of each primer, and 1 U of Ex Taq Polymerase (Takara Bio, Inc.). The PCR cycling conditions were as follows: initial denaturation at 94°C for 2 minutes followed by 16 cycles of denaturation at 94°C for 1 minute, annealing at 58°C for 1 minute, extension at 72°C for 2 minutes, and a final extension at 72°C for 5 minutes. PCR products were analyzed by agarose gel electrophoresis. Each amplified product was semiquantified by measuring the peak areas of capillary electrophoresis using an Agilent 2100 Bioanalyzer with a DNA1000 kit (Agilent Technologies). The sequences of all the detected bands were confirmed by subcloning and sequencing as previously described (Tran et al., 2006). As an internal standard, the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene was also amplified by reverse transcriptase (RT)-PCR from cDNA, using primers spanning exons 3–6 of the GAPDH cDNA (Zhu et al., 2007).
The percentage of spliced products among the total transcripts was calculated by the following formula: splicing efficiency (%)=[spliced/(spliced+unspliced pre-mRNAs)]×100 (Zheng and Baker, 2000). The percentage of the skipped exon among the total transcripts was defined as [Δexon45/(normally spliced product+Δexon45)]×100. The ratio of normally spliced mRNA to the internal standard GAPDH was calculated and its relationship with the pre-mRNA was analyzed.
Statistical analysis
All analyses were performed using GraphPad Prism 5 (GraphPad Software, Inc., San Diego, CA). The mean±standard deviation was derived from 2 or more independent experiments. Both nonlinear and linear regressions were used to determine the mechanism of AO85-induced exon 45 skipping.
Results
Cell-free splicing of pre-mRNA
Synthesized pre-mRNA was incubated with HeLa cell nuclear extract and the resulting products were analyzed by RT-PCR of total RNA extracted from the reaction mixture. Amplification of a fragment extending from exons A to B revealed 3 products on agarose gel electrophoresis (Fig. 1b). The largest product corresponded to the substrate pre-mRNA that maintained the 3 exons (exons A, 45, and B) and 2 introns (introns A and 45). The middle-sized product lacked the upstream intron A from the pre-mRNA (Δintron A), indicating a splicing intermediate. The smallest band lacked both introns (introns A and 45), corresponding to the mature mRNA. All 3 products showed the expected exon/exon junction sequences and there were no unexpected splicing products caused by cryptic splice site activation. Mature mRNA was obtained most abundantly at 30°C compared with at 37°C or 40°C (data not shown), as reported previously (Habara et al., 2009).
We then monitored the time-course of the splicing reaction by semiquantitative RT-PCR (Fig. 2a). With an incubation time of zero, only the pre-mRNA was detected. At 15 mins, Δintron A was also detected. This indicates that the 5′ intron, intron A, was spliced first in this cell-free splicing system. At 30 minutes' incubation, the amount of Δintron A increased and the mature mRNA was detected. Trace amounts of Δexon 45 were also detected (Fig. 2a).
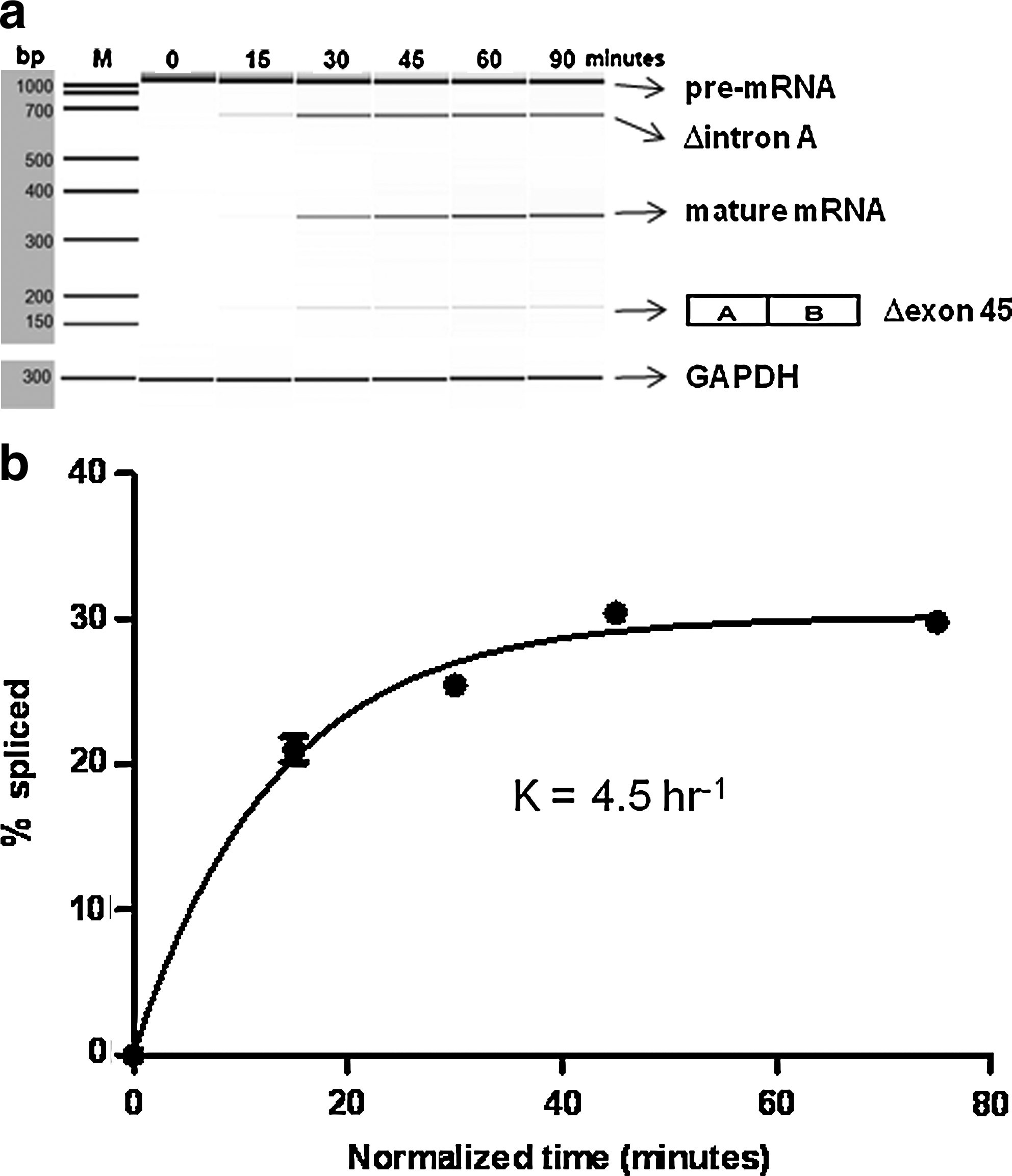
Time-course of the cell-free splicing reaction.
We then analyzed the percentage of spliced product. Because the Δintron A splicing product appeared at 15 minutes (Fig. 2a), the first 15 minutes was considered an initial lag period (Hicks et al., 2005). When the 15-min lag period was taken into consideration, the appearance of spliced products followed a profile characteristic of a first order reaction (Fig. 2b). The product appearance data fitted the pseudo-first order rate description Y=C×(1–e–kt), where Y is the fraction spliced, C is the fraction spliced at the end-point of the reaction, k is the apparent rate constant, and t is the time (Hicks et al., 2005). The observed rate constant was 4.5±0.42 hour–1, fitting well to the 1-phase association model with R2=0.9905 (Fig. 2b) and confirming that this cell-free splicing reaction is time dependent. In the following experiments, the incubation time was shortened to 30 minutes.
Induction of exon 45 skipping with AO85
To examine whether AO85 was able to induce exon 45 skipping in the cell-free splicing system, 0–160 nM AO85 was added to the incubation mixture, and the resulting RNAs were analyzed by RT-PCR. In the absence of AO85, 3 products, corresponding to pre-mRNA, Δintron A, and mature mRNA, were clearly observed (Fig. 3a). At 20 nM AO85, the percentage of Δexon 45 increased, whereas both Δintron A and the mature mRNA decreased. At 160 nM AO85, almost 80% of the total product was Δexon 45. When the percentage of Δexon 45 was plotted against AO85 concentration, the percentage increased linearly (Fig. 3b). The data fitted well to a dose–response model, with R2=0.9990. From this, the EC50 was calculated as 58.0 nM.
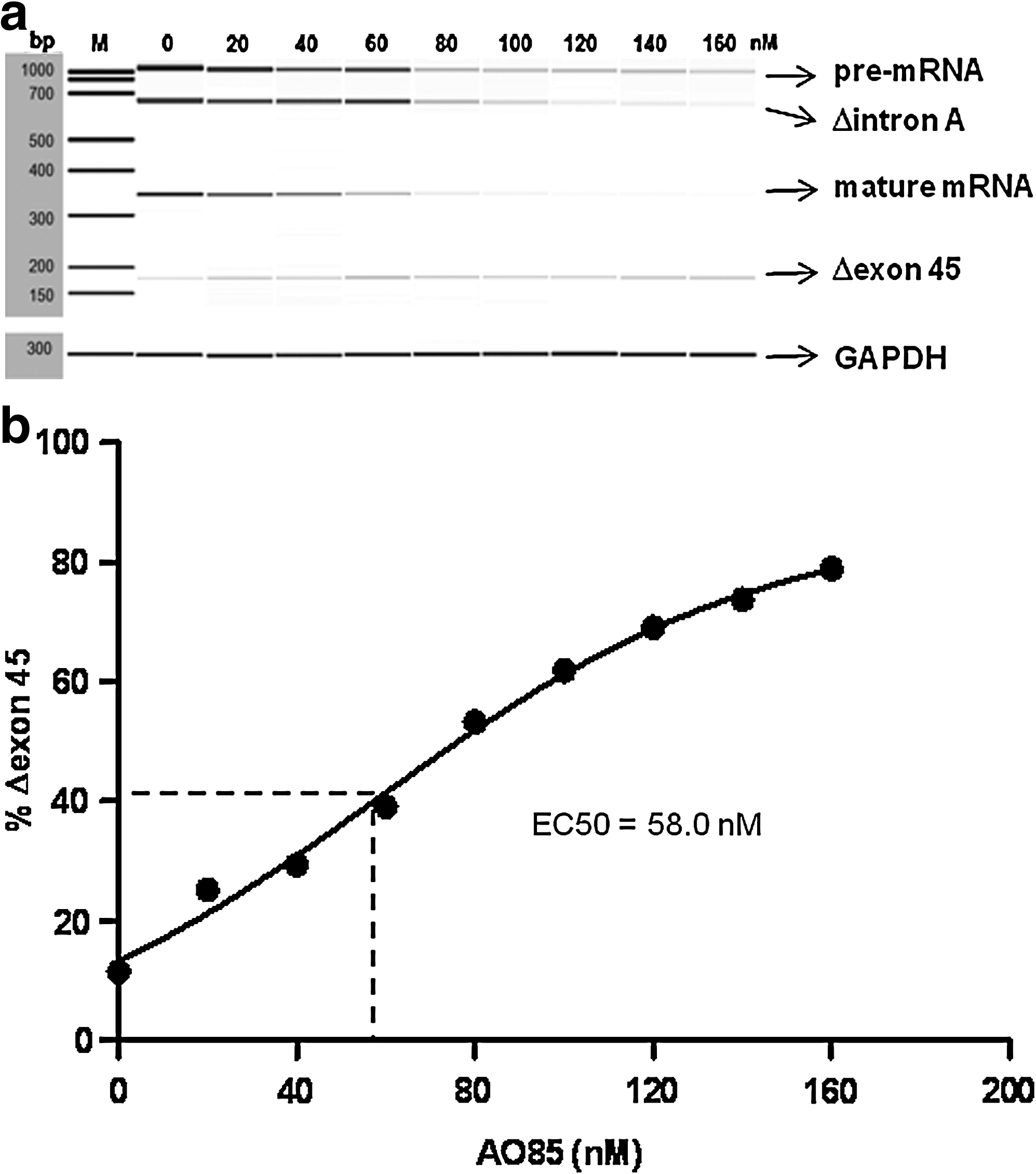
Dose dependency of exon 45 skipping.
Competitive fashion of AO85 action
We next analyzed action patterns of AO85 in a total splicing reaction by measuring the production of mature mRNA with different concentrations of AO85 and pre-mRNA. The amount of pre-mRNA ranged from 0 to 10.5 nM with 3 different AO85 concentrations (0, 20, and 40 nM). The amount of mature mRNA was normalized to that of GAPDH. On a Lineweaver-Burk plot (Fig. 4a), the data showed that AO85 inhibited the production of mature mRNA in a dose-dependent manner and that 3 lines linking spots met together at the Y axis, indicating a competitive fashion of AO85 action. The Ki of the inhibition was found 10.1 nM (Fig. 4b).
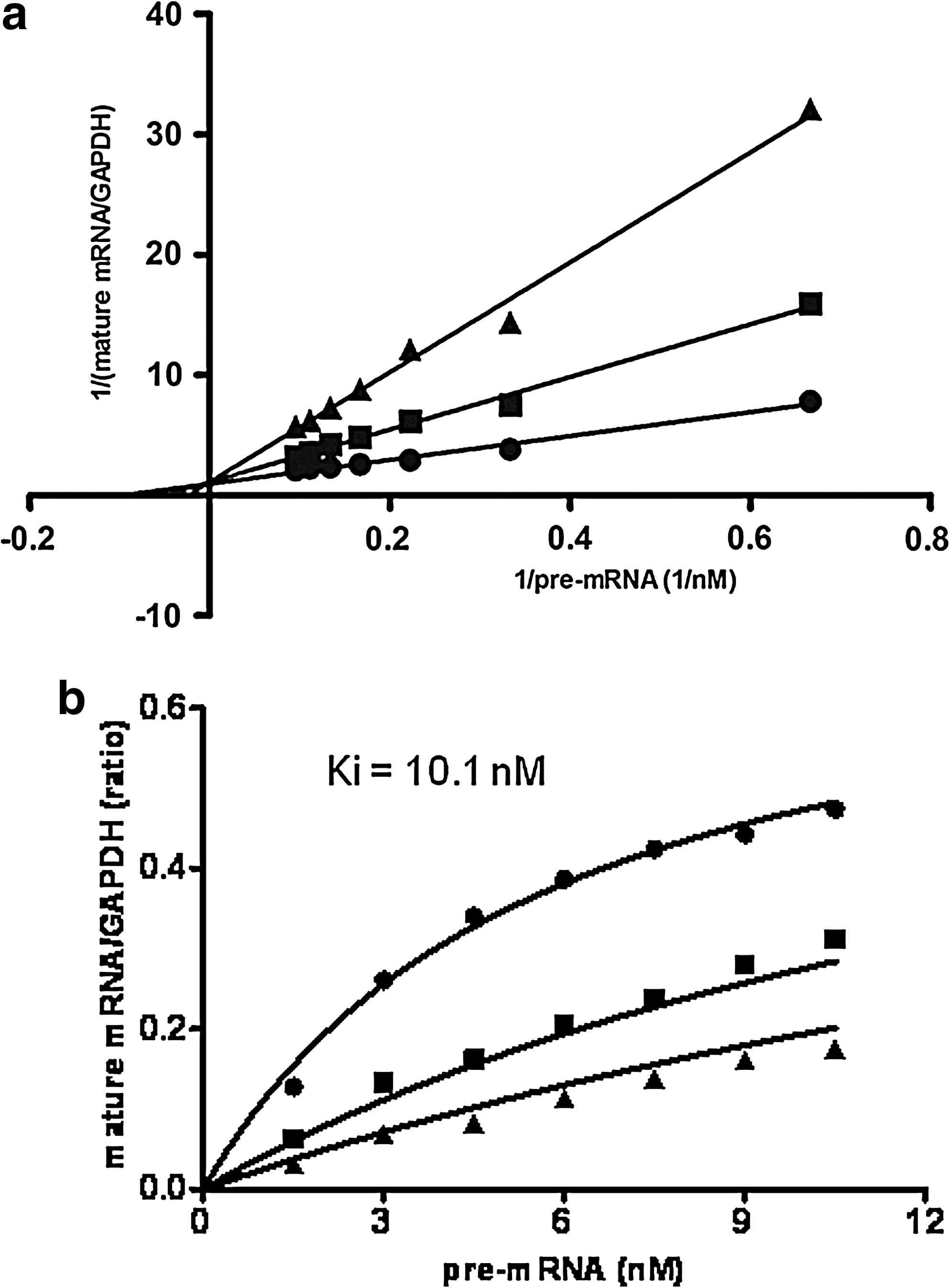
Competitive fashion of of AO85 action.
Discussion
We showed that AO85 induced exon 45 skipping in a cell-free splicing system that contained substrate pre-mRNA and HeLa cell nuclear extract, indicating that exon skipping does not require intact cells. Regardless of the shortened introns (Fig. 1) the pre-mRNA was subjected to exon skipping. The in vitro splicing assay has been shown useful to examine antisense-mediated exon skipping, although there are limitations in applying the results in understanding in vivo splicing (Spitali et al., 2009). One of the most important limitations is that in vitro splicing requires the use of relatively short transcripts (approximately 1,000–2,000 bp) that usually lack the complexity and length of natural pre-mRNA, and thus may not identify the rate limiting steps for expression in vivo (Nasim et al., 2002). In our in vitro splicing system total amount of RNA decreased at the high concentration of AO85 (Fig. 3a). This may be due to tight connection of AO85 to the target sequence, hampering PCR amplification or unknown RNAse activation.
The cell-free splicing reaction had an initial lag period of up to 15 minutes (Fig. 2). Regulatory elements located within introns and exons guide the splicing complex, the spliceosome, and auxiliary RNA-binding proteins to the correct sites for intron removal and exon joining (Pandya-Jones and Black, 2009; Ward and Cooper, 2010). Presumably, exogenously added pre-mRNAs are initially coated with heteronuclear ribonucleoproteins (hnRNPs) that are abundant in nuclear extracts, and the competing association and dissociation between hnRNPs and components of the spliceosome requires time (Hicks et al., 2005). We assume that the observed lag period corresponds to the spliceosome association. After this initial phase, the splicing reaction proceeded linearly.
AO85, which was identified through trial-and-error, is considered to bind to an exon splicing enhancer (Takeshima et al., 2011). Nuclear proteins bound to the enhancer sequence can promote exon definition by directly recruiting other splicing factors and/or by antagonizing the action of nearby silencer elements. Thus, failure of nuclear proteins to bind to the enhancer sequence would prohibit recognition of the exon by the splicing machinery and cause exon skipping (Takeshima et al., 1995; Pramono et al., 1996; Freier and Altmann, 1997; Stahel and Zangemeister-Wittke, 2003; Aartsma-Rus et al., 2009b; Popplewell et al., 2009). AO85 is expected to bind to a region targeted by 2 SR proteins, SRp30c and SRp40 (Mathews et al., 1999), suggesting that AO85 competes with these 2 SR proteins for binding the pre-mRNA. The binding site for SRp30c is partially located where the pre-mRNA adopts an open secondary structure, whereas the predicted binding site of SRp40 is located within a closed structure (Mathews et al., 1999; ZUKER, 2003). AO85 probably competes with SRp30c, but this needs further examination. Our results showing a competitive fashion of AO85 action (Fig. 4a) support the idea of competition with nuclear proteins.
Our study of the kinetics of AO85-mediated exon skipping in in vitro splicing revealed that (1) AO85 induced exon 45 skipping by inhibiting the production of mature mRNA, with a Ki of 10.1 nM. This relatively low Ki value indicates that AO85 is a potent competitor in the recognition of exon 45, and (2) the EC50 of AO85 was 58 nM. The IC50 of AOs in gene knockdown has been determined to be between 70 and 220 nM (Grunweller et al., 2003), indicating that AO85 works efficiently in our in vitro splicing system. At 160 nM AO85, exon skipping was induced in approximately 80% of transcripts. In comparison with this, at most 70% of exon skipping has been induced by 500 nM 2′-O-methyl phosphorothioate AOs in another cell-free splicing reaction (Spitali et al., 2009). The recent efforts to improve exon skipping efficiency by optimizing the AO sequence, backbone chemistry, and additional modifications (Kurreck et al., 2002) are worthwhile. Considering that AO85 has low EC50 (58.0 nM) (Fig. 3b), high exon skipping efficiency (80%) (Fig. 3b), and ability to induce exon 45 skipping in cultured human muscle cells (Takagi et al., 2004b), we believe that AO85 has high potential for clinical use. However, further studies are necessary before clinical use.
Footnotes
Acknowledgments
We would like to thank Ms. Kanako Yokoyama for her administrative assistance. This work was supported by a grant from the New Energy and Industrial Technology Development Organization, a Grant-in-Aid for Scientific Research (B) and a Grant-in-Aid for Exploratory Research from the Japan Society for the Promotion of Science, a Health and Labour Sciences Research Grant for Research on Psychiatric and Neurological Diseases and Mental Health, and a research grant for Nervous and Mental Disorders from the Ministry of Health, Labour, and Welfare, Japan.
Author Disclosure Statement
No competing financial interests exist.