
Abstract
MicroRNA (miRNA) sponges allow the selective blockade of a complete family of associated miRNAs, which induce post-transcriptional gene silencing in their target through binding to 3′UTR mRNA. miRNA-365 and miRNA-145 are downregulated in colorectal cancer (CRC) but not in healthy tissues. Based on this, we constructed two vectors by inserting miRNA sponges (one for miRNA-365 and other for miRNA-145), and used enhanced green fluorescent protein (EGFP) as a 3′UTR reporter gene to analyze the ability of each sponge to catch its respective miRNA. Quantitative polymerase chain reaction (qPCR) results corroborated that the expression levels of both miRNAs were lower in CRC cell lines than in normal colon cell lines. Flow cytometry analysis revealed a decrease of the EGFP expression levels in the cell lines transfected with both sponges, being higher on the normal cell line while CRC cell lines presented a minimal decline. Also, this decrease was inversely proportional to the levels of expression of both miRNAs obtained by qPCR. These results were corroborated by fluorescence microscopy, showing a similar decrease in fluorescence. We propose a new vector system to carry in a specific way the expression of genes to CRC cells without affecting healthy cells, preventing damage to healthy tissues.
Introduction
Colorectal cancer (CRC) is the third most commonly occurring cancer in men and the second most commonly occurring cancer in women, and the second in terms of mortality [1]. The risk of metastasis is very high, with this being the main cause of mortality. The 5-year survival in CRC patients diminishes from 90%, if the cancer is localized, to 68% if it has spread to contiguous organs and/or to lymphatic nodes [2]. This aggressiveness and the need for more effective treatments have led to the study of the microRNAs (miRNAs), which are found suppressed or overexpressed in this type of cancer [3].
miRNAs are short noncoding RNAs of ∼22 nucleotides, which induce post-transcriptional gene silencing in its target through binding to 3′UTR mRNA [4]. The miRNAs are involved in diverse biological processes such as cell cycle control [5], cellular differentiation [6], development [7], and metabolism [8]. They are also involved in diseases such as diabetes [9], neurodegenerative diseases [10], and cancer [11,12]. Around 50% of the miRNAs are located in “fragile” areas of the genome, regions of loss of heterozygosity, or breakpoints associated with cancer [13].
The expression profiles of miRNAs are characterized by differences between normal and tumoral tissues, as well as among types of tumors [14]. These characteristic profiles can be used as biomarkers for the determination of prognosis, response to chemotherapy, prediction of the efficiency of the treatment, and susceptibility of a patient to suffer a disease [3] (Table 1).
MicroRNA Involved in Colorectal Cancer
miRNA, microRNA; CRC, colorectal cancer.
The expression of the miRNA-129 induces cell apoptosis through the cell cycle arrest between phases G1 and G2. This miRNA is found deregulated in patients with CRC in stages II and IV [15]. The miRNA-200 family act as tumor suppressors in the epithelial–mesenchymal transition, the mirRNA-200b being related to cell growth, and its downregulation to a worse prognosis and metastasis in CRC [16]. The miRNA-16-1 participates in processes of regulation of apoptosis and cell cycle. Its negative regulation of the KRAS expression entails the inhibition of tumor growth in CRC, thus acting as a great candidate to becoming a therapeutic target [17]. miRNA-126, whose expression seems to be bound to the expression of vascular endothelial growth factor A, could be used as response biomarker to a certain treatment [13]. Also, miRNA plasma panels (mir-532-3p, -331, -195, -17, -142-3p, -15b, -532, and -562) can be used to quickly identify the polyps in plasma or (mir-431, -15b, and -139-3p) the CRC stage IV [18].
This work focuses on the downregulated miRNAs in CRC, miR-145 and miR-365. The miRNA-365 is found situated in the region 13.12 of chromosome 13, and it is present in different stages of regulation according to the type of cancer, being strongly deregulated in the case of glioma, while appearing over-regulated in breast cancer [19] or deregulated, based on the type being treated [20]. miRNA-365 is often downregulated and involved in regulating cell proliferation, differentiation, and apoptosis in numerous cancer cells, such as CRC [21–24]. This regulation of proliferation is due to the fact that a decrease in miRNA-365 positively regulates cyclines such as D1 and cdc25A, which promote proliferation [23]. Bcl-2 and Cyclin D1 are some of the target of miRNA-365. Nie et al. [25] demonstrated that Cyclin D1 and Bcl-2 are correlated with miRNA-365, and its downregulation to prognosis in CRC patients. On the contrary, cluster miRNA-143/miRNA-145 is a tumor suppressor usually downregulated in several tumors, being noticeable in the metastatic tumors of CRC patients, but not expressed in epithelial cells [26–29]. A significant deregulation of this miRNA has been observed in CRC patients after chemotherapy [30], besides a negative fold change of miRNA-145 in stages II, III, and IV CRC [31]. Tanoglu et al. [32] analyzed 16 miRNA expression profiles of 40 patients with recurrent and nonrecurrent CRC, indicating that the miRNA-145 expression was downregulated. These data suggest miRNA-145 as a good biomarker for an early detection of this disease [33]. Some of miRNA-145 targets are apoptosis inhibitor 5, K-RAS, ERK5, and insulin receptor substrate1 [34]. Exogenous miRNA-145 decreases ERK5 and c-Myc expression, cellular growth and tumor volume in mice [35].
The inhibition of miRNA may use different tools such as miRNA sponges, miRNA masking, antisense oligonucleotides, or molecule inhibitors. Anti-miRNA oligonucleotides (AMOs) are being used to suppress dysfunctional miRNAs by steric blocking the target miRNA. AMOs mostly stimulate an RNase-dependent degradation of the target miRNA [36,37]. In contrast to AMOs, miRNA sponges do not degrade the miRNA when they find it, leaving it intact. A miRNA sponge is an mRNA sequence of multiple tandem binding sites (MBS) to target specific miRNAs, allowing to inhibit the expression of diverse miRNAs of the same family [38]. The binding specificity of the miRNA toward the end of 3′ UTR of its mRNA target is determined by its pairing with the “seed” sequence [39]. Said sequence contains certain bases, which do not pair with the mRNA target, giving place to small loop. Likewise, the sponges containing bulged sites that are mispaired opposite to miRNA positions 9–12 show a better and more stable binding efficacy than sponges with complete complementarity [40]. This may be caused by an increase in endonucleolytic cleavage activity of AGO2 in the perfect binding of the miRNA, which leads to the degradation of the sponge [41].
Usually, sponges contain from 4 to 10 MBS separated by a few nucleotides each. It has been demonstrated that a greater number of MBS increase the probability of sponge RNA degradation. The efficacy of the sponge also depends on the concentration of sponge RNAs relative to the concentration of the miRNA; therefore, the use of the strong promoters would maximize sponge expression [42]. The main advantage of these miRNA sponges resides in their expression, stable and easily inducible by means of promoters such as U6 or cytomegalovirus (CMV) [43,44]. The possibility of gene vector-encoded sponges offers new means of controlling miRNAs for therapeutic regulation of gene expression [45,46]. Usually, these miRNA sponges are generally designed to connect to reporter genes to be well tracked and sorted [40]. Reporter genes are usually placed directly upstream of the MBS, such that the protein expression directly represents sponge RNA expression [42].
We constructed a vector by inserting seven tandem-arrayed microRNA binding sites to downstream of the CMV promoter, and used enhanced green fluorescent protein (EGFP) as a 3′ UTR reporter gene to analyze the ability of the miRNA sponges to catch the miRNAs 365 and 145. miRNA sponges must bind to the endogenous cellular miRNA, so the expression of the EGFP must be inhibited. The EGFP expression levels are therefore proportional to the number of molecules of the endogenous cellular miRNA.
The aim of our work was to use a new gene-specific tumor tissue driving system in which the target gene will carry at its 3′ UTR end a miRNA sponge for miR-145 or miR-365. These miRNAs are upregulated in healthy colon tissues but downregulated in CRC. Therefore, in healthy cells, the target gene will be silence post-transcriptional by the binding of miRNA (miR-145 and miR-365, respectively) to the sponge, whereas in CRC cells, where miR-145 and miR-365 levels are low, this gene will be expressed being able to perform its function. This new system could restore CRC downregulated gene expression levels or specifically target the expression of killer genes against CRC cell without harming healthy cells.
Materials and Methods
Cell culture
The human colon cancer cells, RKO, T-84, SW480, HCT-15, CACO2, HT-29, and HCT-116, and CCD-18Co human colon normal cells were provided by the Instrumentation Service Center of the University of Granada, Granada, Spain. All cell types were grown in Dulbecco's modified Eagle's medium (Sigma, St. Louis, MO), supplemented with 10% fetal bovine serum and 1% streptomycin–penicillin (Sigma), under air containing 5% CO2 and in an incubator at 37°C.
RNA isolation and real-time-polymerase chain reaction
Total RNA from cells was extracted using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. A total of 1 μg RNA was used as template for cDNA synthesis. Polymerase (cloned) (Invitrogen) catalyzed the addition of adenosine to the 3′ end of RNA in a sequence-independent manner. The parameters used were as follows: 37°C for 60 min and 70°C for 5 min. Reverse transcription reactions were performed using SuperScript™ II Reverse Transcriptase (Invitrogen) with universal reverse transcription primer. The reverse transcription thermocycling parameters were as follows: 42°C for 80 min and 85°C for 5 min. Quantitative polymerase chain reaction (qPCR) was performed using SYBR green supermix (iTaq Universal SYBR Green Supermix [Bio-Rad Laboratories, Hercules, CA]). The thermocycling parameters were as follows: 95°C for 30 s, 95°C for 15 s, and 60°C for 60 s. All quantitave real time-polymerase chain reaction (RT-PCR) assays were performed on an ABI 7900 system (ABI), and 2−ΔΔCt method was applied for calculation of relative levels of expression. Small nucleolar RNA 44 (SNORD44) was used as the internal reference. Sequences of the miRNA sponge oligonucleotides were obtained from miRNABase (miRbase; www.mirbase.org), and the primers were synthesized according to them (Table 2).
Primers Designed for the Amplification by Quantitative Polymerase Chain Reaction
SNORD44, small nucleolar RNA 44.
Sponge design and construction
The hsa-miRNA-145-5P and has-miRNA-365-3p sequences were provided by miRBase database. miRNA sponge sequences were chosen based on optimizations previously described by several authors. The features adopted here included the following: 7 microRNA binding sites (MBS) [42,47,48]; a central bulge with 4 nucleotide mismatches at positions 10–13 of the MBS [48,49]; 4 nucleotide spacer between each MBS [47–49]. For directional cloning, specific restriction sites for EcoRI and XbaI, and SpeI and SacII enzymes were added to the 5′ and 3′-end, respectively. Both sponges were synthesized (IDT, Coralville, IO) and subcloned into pEGFP-C1vector (Clontech), which expresses the EGFP, that was used as reporter gene. Subcloned bacteria for both constructions, miR-145 sponge vector (pEGFP-145) and miR-365 sponge vector (pEGFP-365), were tested by PCR (Table 3), and those that were positive were verified by DNA sequencing.
Primers Designed for the Amplification by Polymerase Chain Reaction
Transfection of cell lines
One day before transfection, CCD-18Co, T-84, HTC116, and MC38 cell lines were seeded into 24-well plates (5 × 104 cells per well). In brief, a transfection mixture was prepared by adding 500 ng vector and 1 μL Lipofectamine 2000 (Invitrogen). Experiments were performed in four groups: cell lines transfected with pEGFP-145; cell lines transfected with pEGFP-365; cell lines transfected with pEGFP and not transfected cell lines, which were used as controls.
Detection of sponge expression by RT-PCR in vitro
RNA was extracted from RKO, T-84, HTC-116, and CCD-18Co cells with the Rneasy Mini kit (Qiagen). cDNA was generated by means of the Promega Reverse Transcription System (Promega, Madrid, Spain) using total cellular RNA (1 μg). PCR amplification of the sponges, pEGFP-145 and pEGFP-365, was performed to determine correct gene expression as described above (Fig. 4).
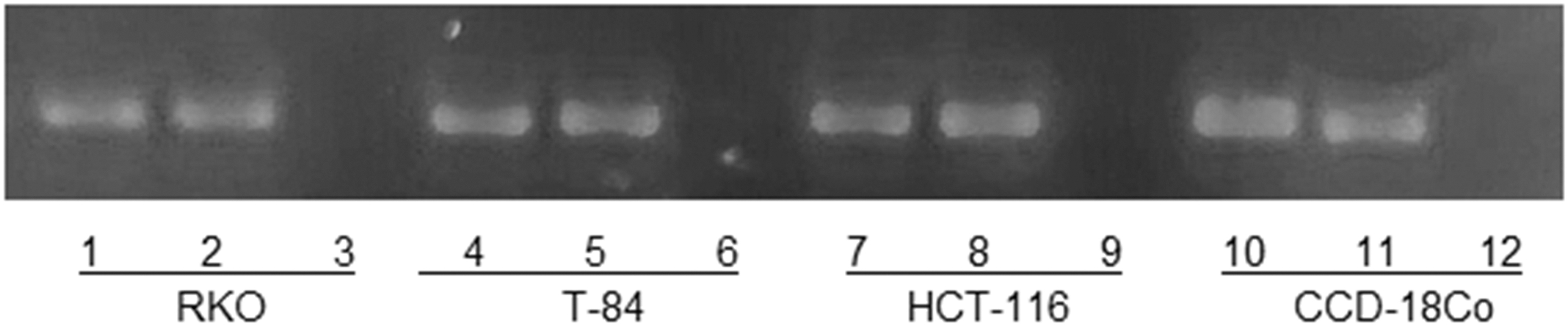
Correct expression of sponges in vitro. PCR of pEGFP-145 and pEGFP-365: RKO: lane 1, pEGFP-145; lane 2, pEGFP-365; lane 3, pEGFP; T-84: lane 4, pEGFP-145; lane 5, pEGFP-365; lane 6, pEGFP -549; HCT-116: lane 7, pEGFP-145; lane 8, pEGFP-365; lane 9, pEGFP; CCD-18Co: lane 10, pEGFP-145; lane 11, pEGFP-365; lane 12, pEGFP.
Detection by flow cytometry
We used EGFP as gene reporter to study the ability of miRNA-145 and miRNA-365 to catch their respective miRNA targets. Forty-eight hours after transfection, the cell lines were collected, and the changes in the EGFP expression levels were analyzed using the Cellfit program with a FACScan flow cytometer (Becton Dickinson, San Jose, CA).
Microscopic analysis
EGFP was used as reporter gene in the used constructions as described above. One hundred nanomolar 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen) was used for nuclear staining. EGFP was excited at 488 nm and DAPI nuclear stain at 364 nm. Fluorescence microscopy analysis was carried out with a Leica DMI6000 microscope (Heidelberg, Germany).
Results
Real-time PCR confirmation of miRNA differential expression
We worked normalizing with SNORD44 instead of 5S, as the first one shows a constitutive expression closer to the miRNAs of interest and a more reliable normalization. The expression of 5S was too high (data not shown). Real-time qPCR results revealed that the expression levels of all miRNAs were lower in all of CRC cell lines than in normal colon cell lines. Also, the expression levels of miR-145 and miR-365 were similar, being on the HCT-116 cell line where minor expression was observed (13.27 ± 0.64 for miR145 and 11.16 ± 0.35 for miR-365) (Fig. 1).
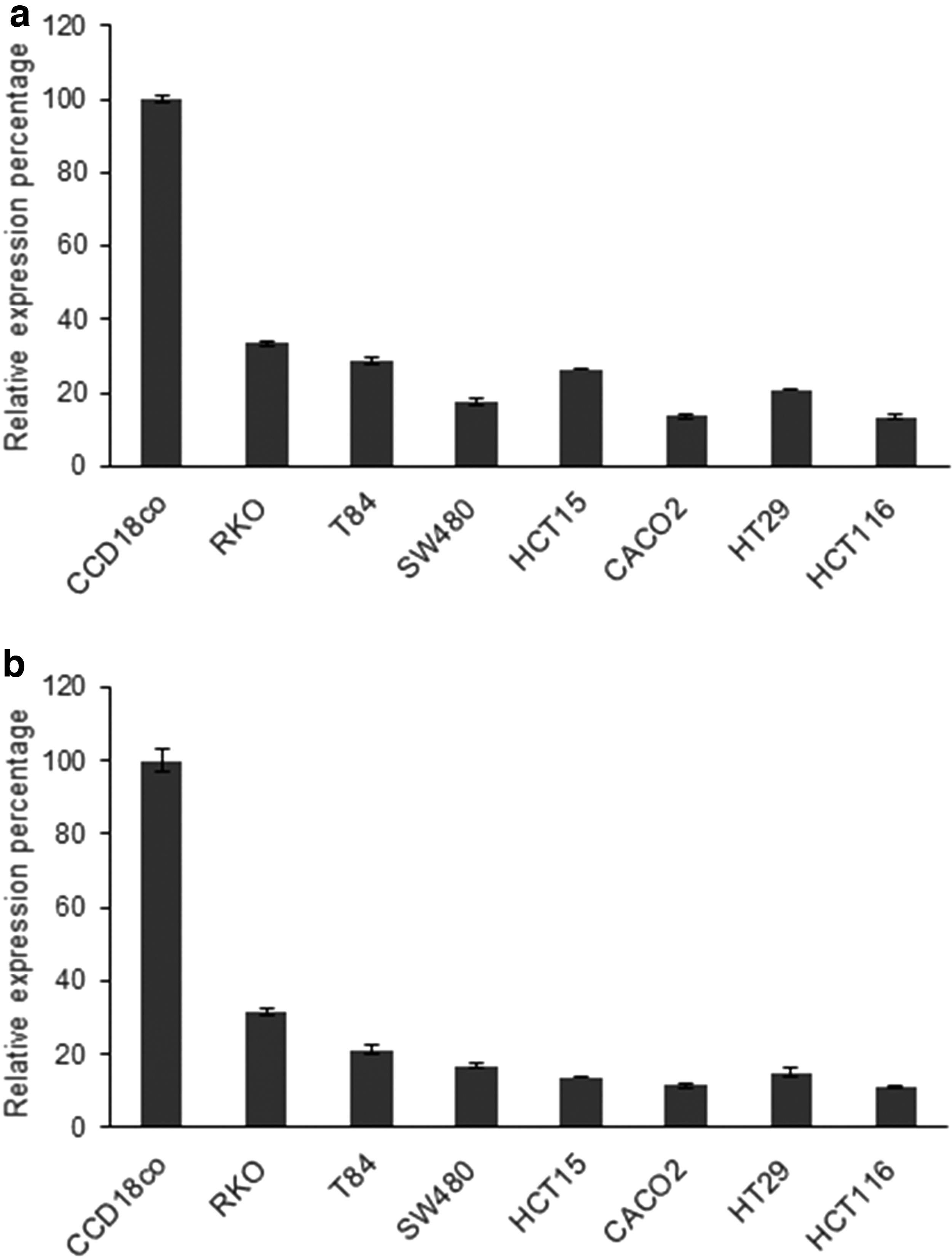
Expression level of miRNA in cell lines.
Development of a simple and effective miRNA sponge expression system
The hsa-miRNA-145-5p and has-miRNA-365-3p sequences were provided by miRbase database (Fig. 2A). The hsa-miRNA-145-5p and has-miRNA-365-3p sponges were designed to the section “Materials and Methods” described above (Fig. 2B), synthesized (IDT) and subcloned into pEGFP-C1 vector (Clontech) (pEGFP-145; pEGFP-365) (Fig. 2C).
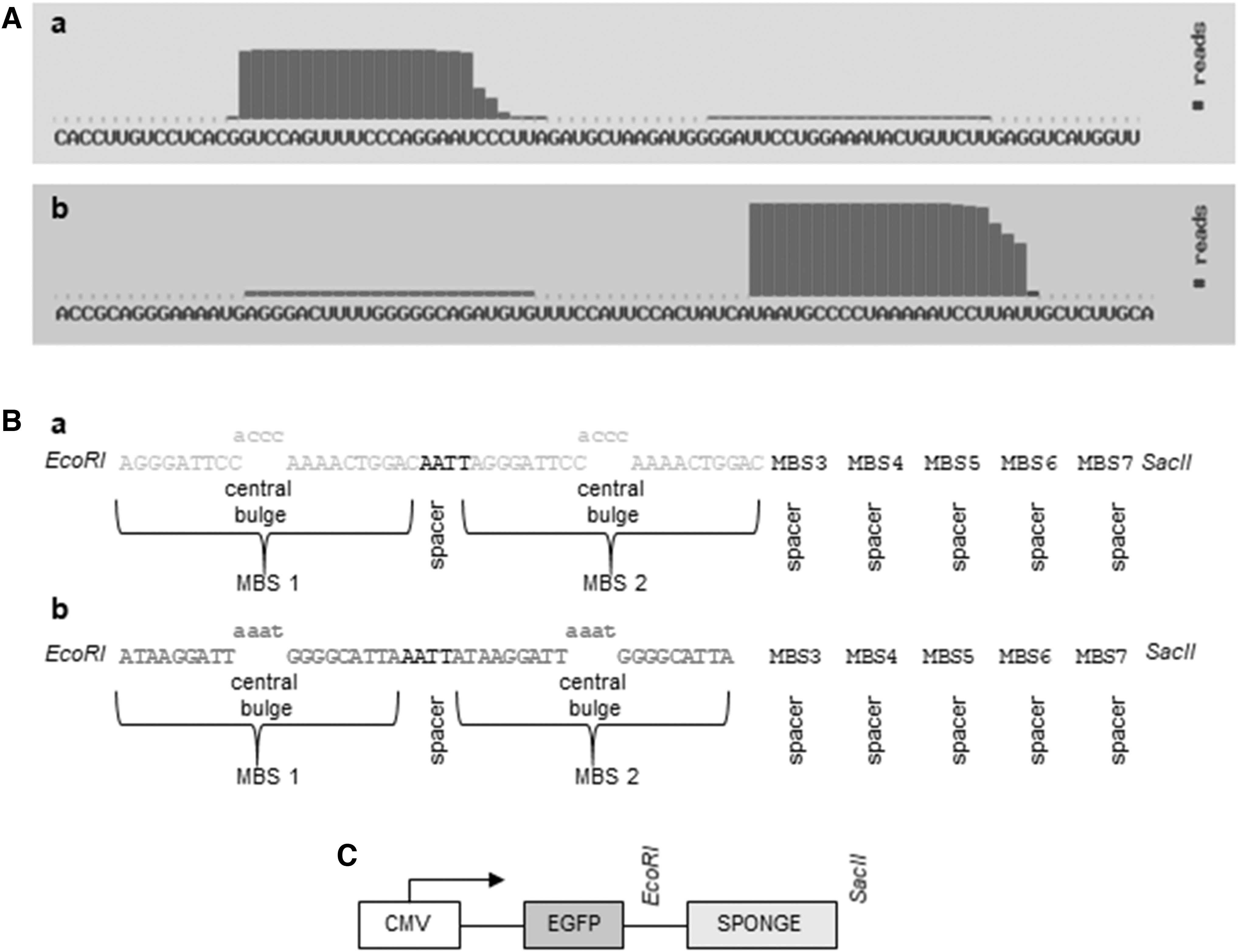
miRNA sponge expression system.
Several colonies were analyzed by PCR, choosing those that showed the size corresponding to the miRNA-145 and miRNA-365 base pairs, respectively (Fig. 3), for subsequent sequencing (Supplementary Data S1 and S2).
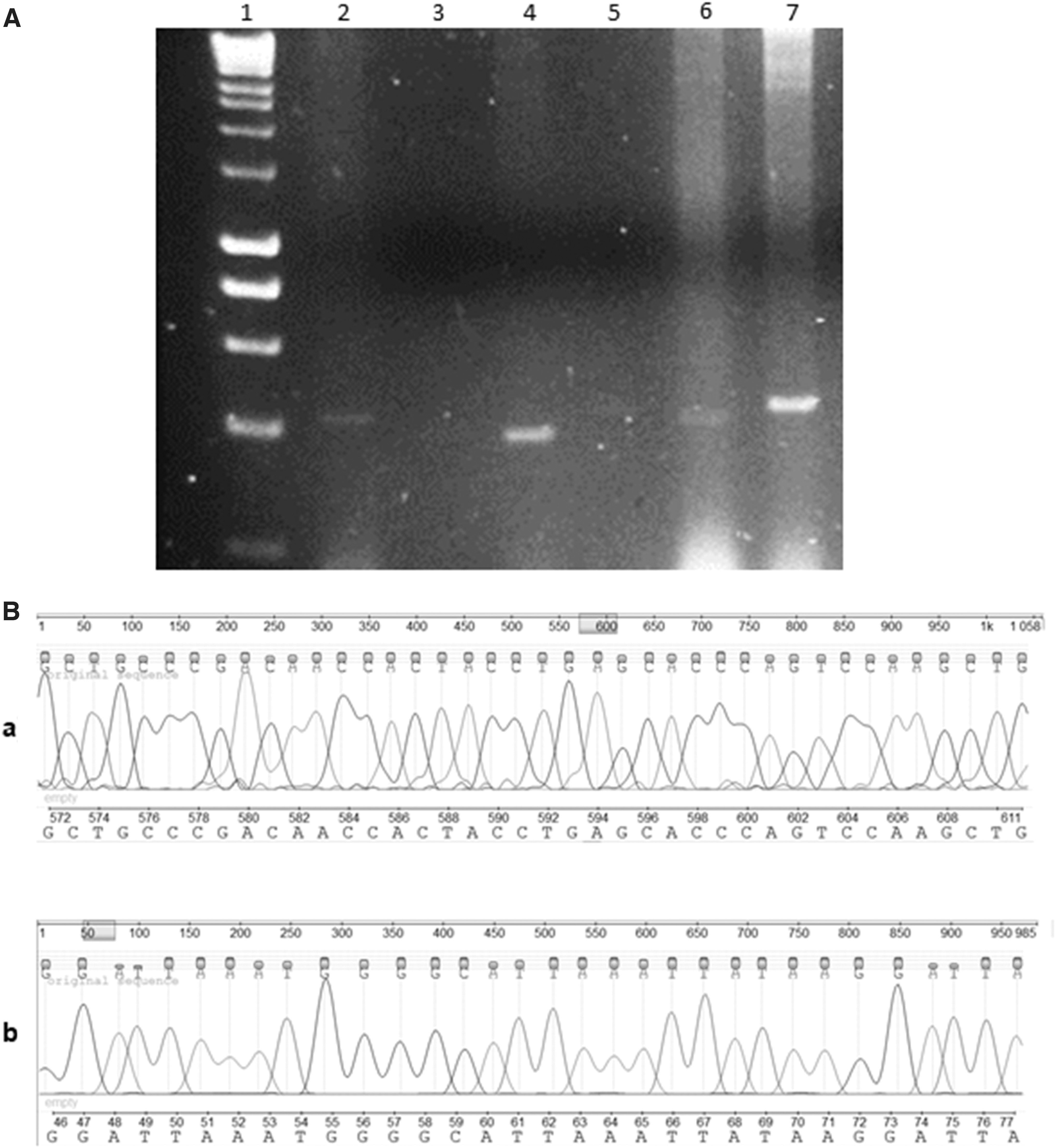
Determination of the correct subcloning of miRNA-154 and miRNA365 into pEGFP-C1 vector.
Correct expression of sponges in vitro
pEGFP-145 and pEGFP-365 expression in cell lines were assessed by RT-PCR. As shown in Fig. 4, an amplification 232 bp was found in all cell lines transfected on both constructions, respectively, but no transfected cells did not show amplification.
Potential miRNA sponges suppress reporter gene activity
Flow cytometry analysis revealed that all the cell lines transfected with the pEGFP-145; pEGFP-365 showed decrease of the EGFP expression levels. However, this decline was only significant on the CCD-18Co cell lines (41.9% and 46.1% for miR-145 and miR-365, respectively), while CRC cell lines represented a minimal decline: 21.8% RKO, 13.7% T-84, and 9.3% HTC-116, for miR-145, and 12.4% RKO, 4.3% T-84, and 4.6% HTC-116, for miR-145 (Fig. 5A). These results were corroborated with the decrease in fluorescence observed by fluorescence microscopy (Fig. 5B).

Analyses of the ability of the sponges to catch their respective miRNAs.
Discussion and Conclusion
CRC is one of the most common cancers, with a high risk of metastasis, the main cause of mortality. For this reason, it is necessary to find more effective treatments. Specific patterns of deregulated miRNAs have been associated with diagnosis, prognosis, and therapeutic response of tumors, so that they could be used as biomarkers and as a new therapeutic strategy [3,14]. miRNA-365 and miRNA-145 express lower levels in CRC tumor tissues than in healthy tissues. miRNA-365 acts as tumor suppressor, and that is why its downregulation in CRC leads to a faster tumor proliferation and of worse prognosis, ending in metastasis [50]. miRNA-145 is related to the inhibition of metastasis and tumor invasion, and for that reason, its downregulation in CRC decontrols these processes [34].
We studied the expression level of these miRNAs in the human CRC cells, RKO, T-84, SW480, HCT-15, CACO2, HT-29, and HCT-116, and CCD-18co human colon normal cells. At the beginning of the study, we analyzed two housekeeping genes, SNORD44 and 5S. The levels of expression of 5S were higher than those of miRNAs of interest, whereas those of SNORD44 were closer to miR-16, miR-143, miR-145, and miR-365. This fact, together with the frequent use of SNORD44 in commercial kits, such as PerfeCTaR microRNA Assays of Quanta, and that Appaiah et al. [51] demonstrated the misuse of 5S for normalization as it showed much variability in several samples, led us to our decision of using SNORD44 to normalize data in the process of quantification of miRNAs. Real-time qPCR results revealed that the expression levels of both miRNAs were lower in all of CRC cell lines than in normal colon cell lines. Similar results were obtained by Nie et al. [25], whose expression analysis of miR-365 by qPCR was also decreased in the CRC cell lines, HT29, LoVo, SW480, and SW620, compared with normal colon mucosa tissues. Wan et al. [52] also corroborated by PCR that the expression levels of miR-145 were significantly suppressed in the CRC cell lines, HT-29 and HCT-116 cells. To make the assays with sponges, we chose three CRC cell lines: the two lines with higher expression of both miRNAs (RKO and T-84) and the line with lower expression (HCT-116). As control we used the nontumor line CCD-18Co.
Sponges are mRNA molecules able to abduct miRNA molecules thanks to their sequence of multiple tandem binding sites (MBS) [38], allowing to inhibit the expression of diverse RNAs of the same family [53]. The use of some sponges has shown an improvement in various diseases: against miR-23b, reduces tumor malignancy in glioblastoma [54]; against miR-181a, restores chemosensitivity to imatinib mesylate in leukemia [55]; and against miR-221, attenuates neointimal hyperplasia and improves blood flows in vein grafts [56]. The sponge against knockdown of long noncoding RNA called FBXL19-AS1, which is upregulated in metastatic CRC, inhibits migration, proliferation, tumor growth, invasion, and metastasis in vivo [57]. The TUSC7, another long noncoding RNA potential tumor suppressor, could inhibit CRC cell proliferation by completely sponging miR-211-3p [58].
Based on the capability of sponges to abduct specific miRNAs and induce post-transcriptional gene silencing in its target through binding to 3′UTR mRNA [4], and also that the expression profiles of miRNAs are characterized by differences between normal and tumoral tissues [14], we propose to use them to express in a specific way a certain endogenous gene in CRC cell lines, which have a downregulated target miRNA, without affecting the healthy cells, with higher levels of said miRNA.
In this work, two sponges have been designed: one for miRNA-365 and another for miRNA-145, each one of them formed by seven specific multiple tandem binding sites (MBS). Although the sponge activity is proportional to the number of MBS, sponges are usually designed to include 4–12 MBS, because saturation could occur and increase the risk of RNA degradation and genetic recombination [40,59]. Kluiver et al. [48] designed several sponges against miR19 with a different number of MBS, verifying that the one with 6 MBS reaches the maximum inhibition of miR-19, without showing an increase of this inhibition efficiency with a greater number of MBS, indicating that the sponge with 12 MBS was less effective. In this research, we did not analyze the correlation between MBS and miRNA inhibition effectiveness, but we showed that seven MBS were sufficient to strongly inhibit miR-145 and miR-365 in CCD-18Co cells. The MBS contain four nucleotide mismatches in the middle positions, giving place to small loop. Ebert et al. [40] found that sponges with small loop had better and more stable binding efficacy than sponges with complete complementarity. Also, sponges that are perfectly base paired are vulnerable to Ago2-mediated endonucleolytic cleavage, more easily than sponges with imperfect binding [41]. Kluiver et al. [49] subcloned sponges against miRNA-19, one with MBS with perfect complementarity and another with MBS with imperfect complementarity, in the 3′UTR of the GFP. Both sponges were transfected in WEHI-231 cells, murine B-cell lymphoma cells with high expression levels of miRNA-19. In both cases, a decrease in the GFP expression was observed by flow cytometer, this decrease being higher in the sponges forming protuberances than in those that had a perfect complementarity. Besides, MBS were separated by a “spacer,” a short four nucleotide sequence modification, to optimize the binding of miRNA to every possible MBS and reduce the chance of formation of RNA secondary structure [59].
To determine the capability of inhibiting the expression of endogenous genes, the EGFP gene was chosen as gene reporter, expressed downstream of the CMV promoter in the pEGFP-C1 vector (Clontech). Both sponges were subcloned in this plasmid in the 3′UTR of the EGFP (pEGFP-145 and pEGFP-365) and transfected in the cell lines. Flow cytometry analysis revealed a decrease of the EGFP expression levels in the cell lines transfected with both sponges with respect to the lines transfected with the pEGFP. This decline was higher on the CCD1–8Co cell lines, while CRC cell lines presented a minimal decline. In addition to this, this decrease in the EGFP expression was in concordance with the values obtained from the study of qPCR, being inversely proportional to the levels of expression of miRNA-365 and miRNA-145. CCD-18Co cells exposed the highest levels of expression for both miRNAs, and exposed the highest inhibition to the EGFP expression when transfected by pEGFP-145 and pEGFP-365. Regarding the CRC cell lines, lines RKO and T-84, whose levels of expression for both miRNAs were the highest, showed a bigger decrease of the expression of EGFP than HCT-116. HCT-116, with the lowest expression of both miRNAs, showed a lower decrease of the expression of EGFP. These results were corroborated by fluorescent microscopy, displaying a decrease in fluorescence in the lines transfected with pEGFP-145 and pEGFP-365, versus the ones transfected with pEGFP, and being that decrease higher in the nontumor line than in the CRC cell lines.
Our results show the proper functioning of the vector system, pEGFP-145 and pEGFP-365, to carry in a specific way the expression of genes to CRC cells without affecting healthy cells.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Fondo Europeo de Desarrollo Regional (FEDER), Plan Nacional de Investigación Científica, Desarrollo e Innovación Tecnológica (I+D+I), Instituto de Salud Carlos III-Fondo de Investigaciones Sanitarias (FIS) through project PI19/01478. This research was developed within the Doctoral Program in Biomedicine of University of Granada.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.