
Abstract
Preventing the harm caused by nerve degeneration is a major challenge in neurodegenerative diseases and in various forms of trauma to the nervous system. The aim of the current work was to investigate the effects of systemic administration of 2,4-dinitrophenol (DNP), a compound with newly recognized neuroprotective properties, on sciatic-nerve degeneration following a crush injury. Sciatic-nerve injury was induced by unilateral application of an aneurism clip. Four groups of mice were used: uninjured, injured treated with vehicle (PBS), injured treated with two intraperitoneal doses of DNP (0.06 mg DNP/kg every 24 h), and injured treated with four doses of DNP (every 12 h). Animals were sacrificed 48 h post injury and both injured and uninjured (contralateral) sciatic nerves were processed for light and electron microscopy. Morphometric, ultrastructural, and immunohistochemical analysis of injured nerves established that DNP prevented axonal degeneration, blocked cytoskeletal disintegration, and preserved the immunoreactivity of amyloid precursor protein (APP) and Neuregulin 1 (Nrg1), proteins implicated in neuronal survival and myelination. Functional tests revealed preservation of limb function following injury in DNP-treated animals. Results indicate that DNP prevents nerve degeneration and suggest that it may be a useful small-molecule adjuvant in the development of novel therapeutic approaches in nerve injury.
Introduction
A
Although PNS neurons have an intrinsic regenerative capacity, regeneration is often compromised or limited by factors such as the severity of the lesion and the distance between the site of injury and the target organ. Blocking axonal degeneration should be an integral part of the development of novel approaches to preserve nerve structure and to optimize conditions for regeneration following trauma. In fact, functional impairment of target muscles can result if nerve regeneration takes too long to complete. Axonal preservation may be a crucial antidegenerative strategy than can not only rescue axons from axoplasmic disintegration but also, as a consequence, protect neuron cell bodies from death and accelerate regeneration. Moreover, blocking degeneration potentially opens a time window in which proregenerative therapeutics can be introduced. Surprisingly, however, few studies so far have effectively explored antidegenerative approaches in the development of treatments for nerve trauma. Strategies have been proposed to limit the progression of WD (Araújo Couto et al., 2004; Camins et al., 2006; Zhai et al., 2003), to support axon regrowth after complete transection (Battiston et al., 2005; Hattori et al., 2008; Pereira Lopes et al., 2006), to bridge disconnected neural stumps (Meek and Coert, 2002), to improve cell survival (e.g., through the use of N-acetylcysteine as a safe pharmaceutical agent that protects cells from oxidative damage; Moschou et al., 2008; Reid et al., 2009; West et al., 2007), and to promote recovery of limb function (Bregman et al., 1997, 2002; A.C. Lee et al., 2005). However, effective strategies to block axonal degeneration are still lacking.
2,4-Dinitrophenol (DNP) is a classical uncoupler of mitochondrial oxidative phosphorylation (Parascandola, 1974) with newly recognized neuroprotective actions (for reviews, see De Felice and Ferreira, 2006 and De Felice et al., 2007). We recently demonstrated that DNP stimulates neuronal differentiation and neurite outgrowth in primary cortical and hippocampal neuronal cultures, as well as in a neuroblastoma cell line, and that this effect is at least in part related to increased neuronal levels of cAMP (Wasilewska-Sampaio et al., 2005). Significantly, modulation of cAMP levels has been proposed as a possible strategy to promote regeneration and functional recovery in spinal cord injury (Hanson et al., 1998; Pearse et al., 2004; Qiu et al., 2002). Other recent studies have shown neuroprotective actions of DNP against different types of neuronal injury (De Felice et al., 2001, 2004; Jin et al., 2004; Maragos et al., 2003; Pandya et al., 2007). Taken together, these previous findings suggested that DNP might exert a protective effect against degeneration of the peripheral nerve.
The current work was driven by the hypothesis that DNP could block axon degeneration in vivo and rescue neurons from death following a crush injury to the peripheral nerve. Results showed a significant reduction in post-injury axonal degeneration in sciatic nerves of DNP-treated animals, verified by morphometric, ultrastructural, and immunocytochemical assays probing important cell-survival-related proteins such as amyloid precursor protein (APP) and Neuregulin-1 (Nrg1), which is directly involved in axonal myelination. Importantly, limb function was significantly preserved in DNP-treated versus untreated injured animals. These findings suggest that DNP can be a novel small-molecule adjuvant in the development of therapeutic approaches in nerve injury.
Methods
DNP solutions and animal treatments
DNP (Sigma, St. Louis, MO) solutions were prepared daily at a final concentration of 20 μM in phosphate buffer saline (PBS), filtered through 0.45 μm Millipore filters, and kept at room temperature and protected from light until use. Twenty adult female Swiss mice (approximate weight 30 g) were randomly divided into four experimental groups: (a) control group, which was maintained under the same conditions as the other groups but did not undergo any surgical procedures; (b) vehicle group, which was subjected to sciatic-nerve injury and received four intraperitoneal (IP) injections of 0.5 ml of PBS (immediately and 12, 24, and 36 h post injury); (c) DNP 2 group, which was subjected to sciatic-nerve injury and received two IP injections of 0.5 ml of 20 μM DNP solution (immediately and 24 h post injury); (d) DNP 4 group, which was subjected to sciatic-nerve injury and received four IP injections of 0.5 ml of 20 μM DNP solution (immediately and 12, 24, and 36 h post injury). Animals used in the functional assay (see Results) received a 0.5-ml injection of 20 μM DNP (or PBS vehicle) immediately after surgery and then once a week for 6 weeks.
Surgical procedures
Surgeries were always performed in the morning period with controlled light and temperature. All animals were deeply anesthetized with ketamine (100 mg/kg), xylazine (5 mg/kg), and atropine (0.2 mg/kg) by intramuscular injection. The left hindlimb was tricotomized, the sciatic nerve was exposed at mid-thigh, and was clipped using a 10-g aneurism clip (Kent Scientific Corp., Torrington, CT) for 1 min. Initial experiments demonstrated that this injury promoted extensive degeneration of the majority of sciatic-nerve fibers after 48 h (data not shown). Animals were sutured, received the first dose of DNP (or vehicle), and were returned to individual cages with food and water ad libitum. All procedures were in agreement with and were approved by the Committee for Ethics on Animal Experimentation of the Federal University of Rio de Janeiro (Protocol # IBQM 023).
Light and electron microscopy
After DNP treatment, animals were sacrificed and subjected to transcardiac perfusion with 4% p-formaldehyde in 0.1M phosphate buffer. Sciatic nerves were dissected and a 1-mm-long segment of the distal portion of the injured nerve located 1 mm away from the lesion site was additionally fixed by overnight immersion in 2.5% glutaraldehyde in 0.1M Na-cacodylate buffer. Corresponding dorsal root ganglia and the proximal segments of sciatic nerves were immersed in the same fixative solution used in perfusion. The glutaraldehyde-fixed material was processed for light and electron microscopy, as described below.
Nerves were removed from glutaraldehyde, washed in 0.1M Na-cacodylate and post-fixed in 1% OsO4 in 0.1M Na-cacodylate buffer containing 5 mM CaCl2 and 0.8% potassium ferricyanide. Nerves were then washed, dehydrated in acetone, and embedded in Epon. Semi-thin sections were collected on glass slides and stained with 1% toluidine blue solution for morphometric analysis. Ultra-thin sections were stained with uranyl acetate and lead citrate, and were imaged on a Zeiss 900 electron microscope operated at 80 kV. In order to allow proper quantitative analysis and reduce possible experimental biases, samples were imaged and documented in a standardized fashion. Briefly, we used 300 mesh microscopy grids (RMS Ultramicrotome MT6000-XC, RMS Inc., EUA) and the upper left corners of 20 individual mesh were photographed at 7000× magnification for each grid. Using axonal cytoskeleton integrity as the ultrastructural criterion for axonal degeneration, two independent observers performed a double-blinded counting of intact axons, partially degenerated axons, completely degenerated axons, and axons with myelin alterations for each experimental condition.
Formalin-fixed material was processed for immunofluorescence microscopy. L4 and L5 dorsal root ganglia and proximal nerve segments from each injured sciatic nerve were washed in 0.1M phosphate buffer, incubated in 30% sucrose overnight, and immersed in Tissue-Tek OCT compound for cryosectioning in a Leica CM1850 cryostat. Twelve-μm-thick cryosections were washed in 0.1M phosphate buffer, pH 8.0, permeabilized with 0.01% Triton X-100, blocked with 50 mM NH4Cl and, subsequently, with 1% and 3% BSA. Sections were incubated overnight with anti-APP (Chemicon, Billerica, MA) or anti-neuregulin (Neomarkers, Fremont, CA) antibodies followed by 40-min incubation with Alexa 555-conjugated anti-mouse IgG (Molecular Probes, Eugene, OR).
Immunofluorescence assays were performed using the same antibodies and dilutions for all experimental groups. Image-acquisition parameters and conditions were rigorously the same for all samples. Eighteen images were acquired on a Nikon TE300 microscope from both injured and uninjured (contralateral to the compression injury) nerves for each of the five animals used in each experimental group. The same protocol was used in nerve sections immunostained for either APP or Nrg1. For each animal, each set of 18 images was grouped into a single montage panel using NIH ImageJ software (Abramoff et al., 2004). This panel was then converted into an 18-bit image and the integrated fluorescence intensity of the whole panel was measured after appropriate thresholding. Thus integrated fluorescence intensities shown on Figures 4G and 5G represent results obtained from a total of 90 images per experimental condition.
Morphometry
Toluidine blue-stained semi-thin sections were observed through a 100 × objective and images were acquired as described above. Five animals were used per experimental group and 200 fibers from each animal were randomly imaged, totaling 1000 fibers per condition. Total fiber and axon areas were manually measured using ImageJ (NIH) software specifically calibrated to our microscope's final magnification. The ratio between axon and fiber areas, known as G ratio, was used as a morphofunctional index that is directly related to nerve fiber electric conductance and myelin thickness (Gillespie and Stein, 1983).
Functional assay
Mice were tested for limb function using the walking track test in a wooden corridor 5 cm wide by 30 cm long covered with a 140 g/m2 paper strip, as previously described (Varejão et al., 2001, 2004). Animals were allowed to explore and walk freely in the walking track test corridor for 5 min per day for at least 1 week before surgical procedures. This training was crucial to avoid unwanted animal stops during the paper stripe crossing. After surgery, animals were allowed to recover from anesthesia and were submitted to the walking track test. The test was repeated once a week for 6 weeks. Animals had their hind paws painted with non-toxic ink immediately before crossing the wooden corridor, so as to record their footprints for later analysis. The distance between the first and fifth toes of the injured limb's paw was used as a parameter to evaluate loss of limb function following injury (Bozkurt et al., 2008). Footprints from the uninjured limb of each animal were used as controls.
Statistical analysis
Statistical significances of differences between experimental groups were analyzed by analysis of variance (ANOVA) using GraphPad Prism (GraphPad Software, CA). A p value of 0.05 was regarded as significant and all results are expressed as mean ± SEM.
Results
DNP blocks axon degeneration after a crush injury
Sciatic nerves from control, uninjured mice presented a mixture of myelinated and unmyelinated fibers. Myelinated fibers were characterized by a thick and continuous myelin sheath and by axons presenting normal cytoskeleton content. Unmyelinated fibers, on the other hand, were difficult to characterize by light microscopy and were further investigated by electron microscopy (see below). In uninjured animals, nerve cross-sections were filled with fibers presenting no indication of degeneration (Fig. 1a). Forty-eight h after injury, sciatic nerves from mice that received vehicle injections exhibited extensive areas of edema and a significant decrease in fiber density (Fig. 1e). The sizes and shapes of fibers in sciatic nerves from vehicle-treated injured animals were considerably more heterogeneous than in nerves from control mice and exhibited morphologies suggestive of ongoing WD. Inspection of nerves from animals in the DNP 2 group (which received two IP injections of DNP immediately after and 24 h after injury) revealed clear reductions in edema and in signs of degeneration; however, the sizes and shapes of fibers were still more heterogeneous than in control nerves (Fig. 1i). Remarkably, nerves from the DNP 4 group of mice (which received five IP injections of DNP immediately and 12, 24, and 36 h after injury) presented almost no indication of fiber degeneration and very little or no edema (Fig. 1m).
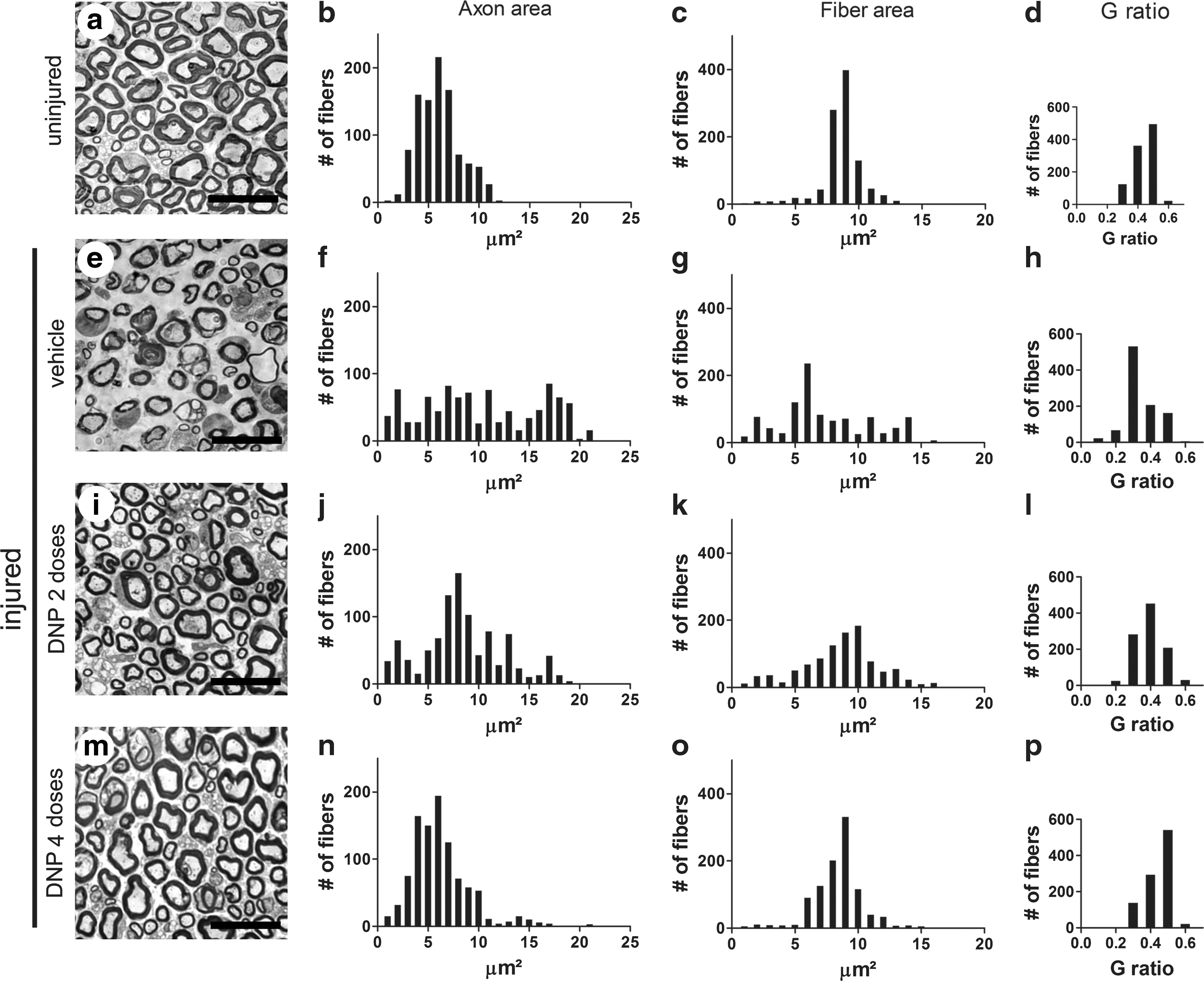
Morphometric analysis of sciatic nerves after injury. Representative semi-thin transversal sections of sciatic nerves from (
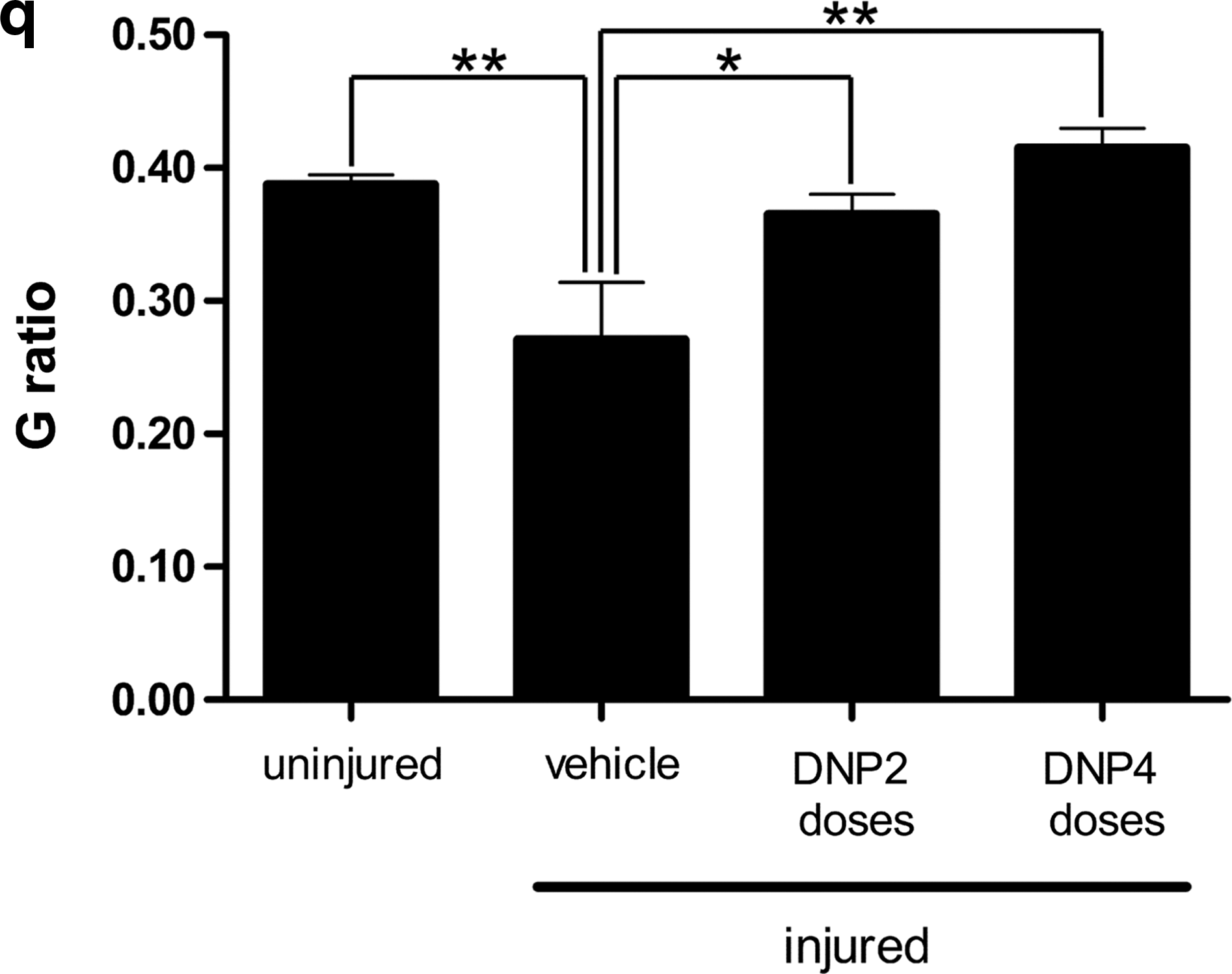
Toluidine blue-stained semi-thin sections of injured and uninjured nerves were used to measure the areas of myelinated fibers and the corresponding axon areas. Fiber and axon areas are defined by the outer and inner limits of the myelin sheath respectively. Figure 1 shows distributions of fiber and axon areas obtained from quantitative analysis of 1000 fibers from five animals (200 fibers from each animal) for each experimental group. Uninjured nerves presented fairly homogeneous distributions of axon areas (centered about 6 μm2) and myelinated fiber areas (centered about 9 μm2; Fig. 1b,c). By contrast, nerves from vehicle-treated injured animals presented very broad distributions of both axon and fiber areas (Fig. 1f,g). The distributions of fiber and axon areas were considerably more homogeneous in the DNP 2 group of injured mice (Fig. 1j,k) and were virtually indistinguishable from the distributions in uninjured control animals in the DNP 4 group (Fig. 1n,o).
The ratio between axon and fiber areas (known as G ratio) defines a useful morphofunctional index that is related to the efficiency of impulse conduction by the nerve (28). Optimal conduction is observed at G ratios close to 0.5 and deviations from this value predict defective conductance and impaired nerve function (28). Figure 1d shows that, as expected, uninjured nerves presented a distribution of G ratios ranging from 0.3 to 0.5. A marked shift in the distribution of G ratios to lower values was observed in the vehicle-treated group of injured animals (Fig. 1h). Interestingly, the distributions of G ratios were considerably preserved in the DNP 2 and especially in the DNP 4 groups of injured mice (Fig. 1l,p respectively). Another way to perform the G-ratio analysis is to calculate the mean value of G ratio in each experimental condition. Figure 1q shows that uninjured fibers presented a mean G ratio of 0.38 ± 0.07. At 48 h post injury, mean G ratio values of 0.27 ± 0.04, 0.36 ± 0.01, and 0.41 ± 0.01 were obtained for vehicle-treated, DNP 2, and DNP 4 groups respectively. Taken together, these results show that DNP treatment preserved tissue architecture, reduced edema, and prevented fibers from entering WD, resulting in morphological and functional preservation of the injured nerves.
Nerve-fiber ultrastructure is preserved by DNP
Disruption of the cytoskeleton and of the integrity of the myelin sheath are ultrastructural criteria for axon degeneration and functional impairment. Transmission electron microscopy analysis of uninjured nerves showed well-preserved myelinated fibers with axons completely filled with cytoskeleton components (microtubules and neurofilaments) and surrounded by thick, electron-dense ring-shaped myelin sheaths (Fig. 2a). Mitochondria profiles were easily found embedded in the axoplasm and showed well-defined cristae. Unmyelinated fibers (Remak bundles) were observed in many regions of the nerve and showed a similar axoplasmic content. At 48 h post injury, partially and completely degenerated axons, presenting partial or complete degradation of cytoskeleton components respectively, were abundantly found in nerve sections (Fig. 2b). Degraded or abnormal myelin sheaths were frequently observed in injured nerves (Fig. 2c). Unmyelinated fibers were also observed, and presented prominent alterations in cytoskeleton content (Fig. 2b). Macrophage and neutrophil infiltration into the distal segment of the degenerating nerve are critical steps in WD and, as expected, those cell types were indeed found in injured nerves of all the animals used in this study (Fig. 2b).
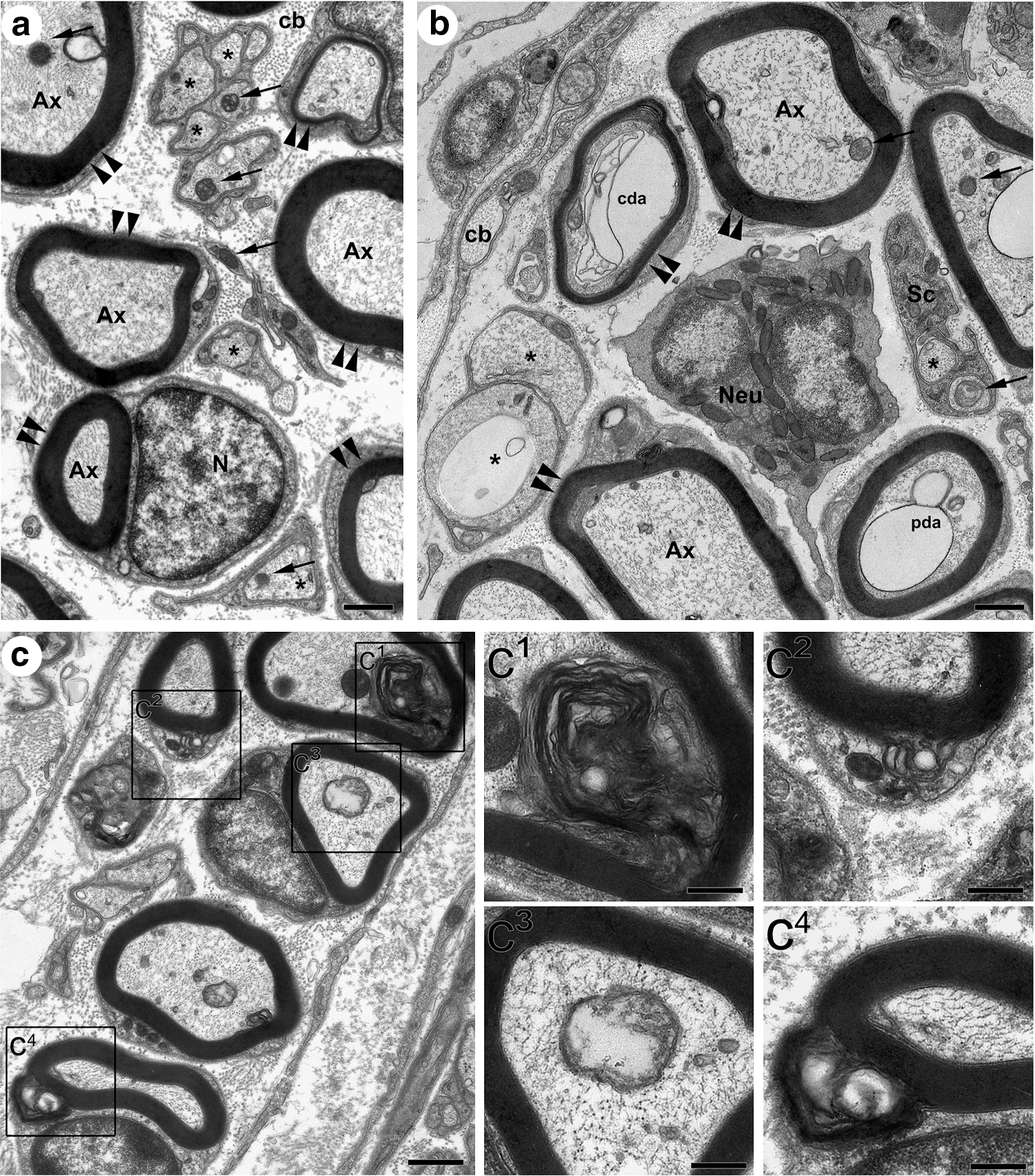
Ultrastructural analysis of sciatic nerve. (
Based on quantitative analysis of fiber ultrastructure (see Methods), fibers were classified as normal, partially degenerated, completely degenerated, or fibers exhibiting myelin abnormalities. Even though normal fibers were present in all experimental groups, their abundance differed sharply between groups. Control, uninjured nerves presented, on average, 120 ± 4 normal fibers counted in 20 independent microscopic fields (n = 5 animals; Fig. 3a). At 48 h post injury, there was a massive reduction to an average of 12 ± 2 normal fibers (counted in 20 microscopic fields per animal; n = 5) in the vehicle-treated injured group, representing a 90% decrease compared to control animals. Significantly, DNP treatment preserved axons from degeneration in a dose-dependent manner (Fig. 3a).
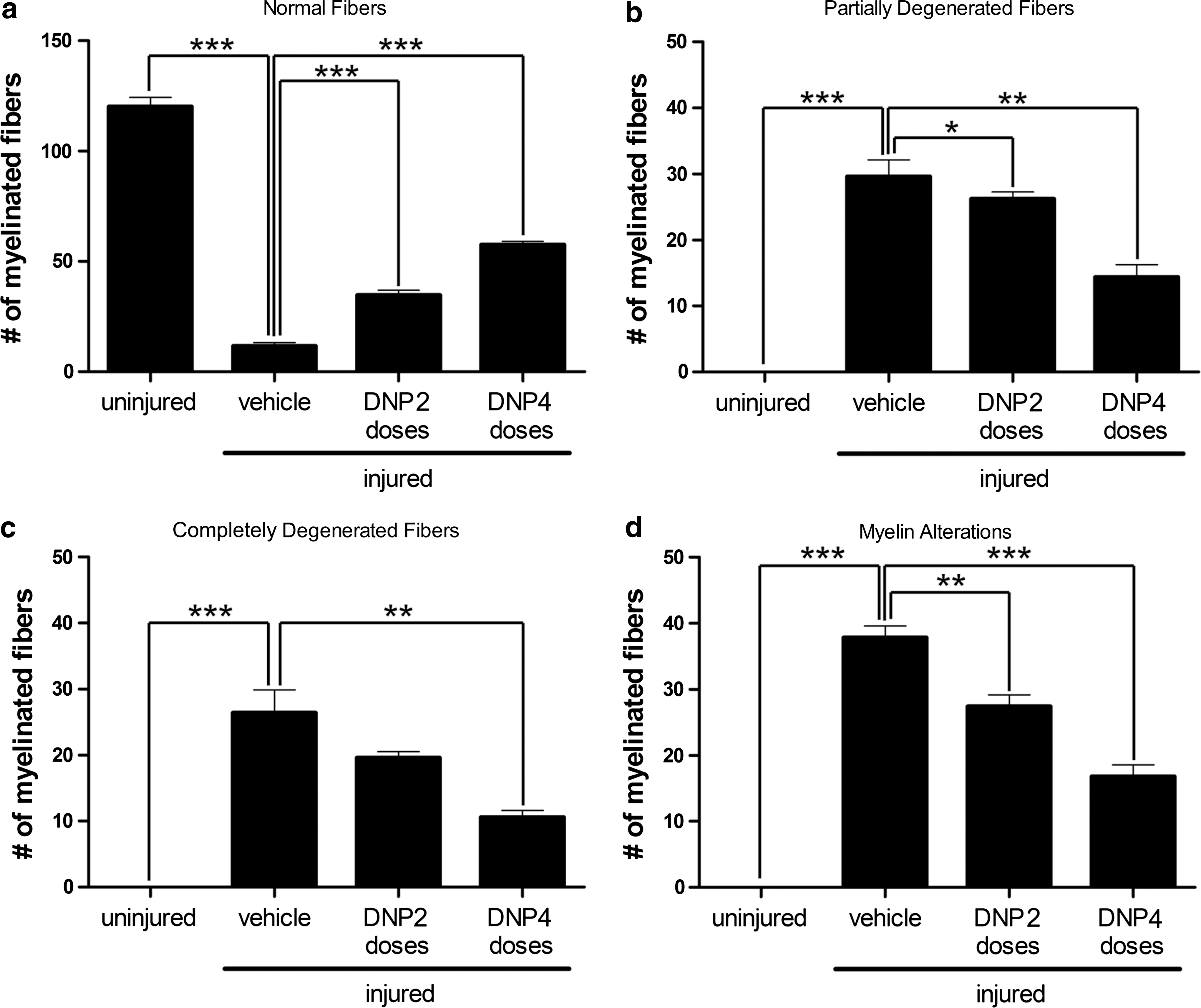
Quantitative analysis of normal and altered myelinated fibers. Twenty independent electron micrographs were analyzed per animal in each experimental group and fibers were classified (see Methods) as (
Conversely, while we did not find any partially or completely degenerated fibers in the uninjured control group, nerves from vehicle-treated injured animals presented 30 ± 3 partially (Fig. 3b) and 26 ± 3 completely degenerated fibers (Fig. 3c). Numbers of partially and completely degenerated fibers were significantly decreased in the DNP 2 and DNP 4 groups (Fig 3b,c).
Similar results were obtained when the number of fibers exhibiting myelin abnormalities was analyzed (Fig. 3d). Control uninjured nerves showed no cases of myelin abnormalities (in 20 independent microscopic fields analyzed per animal; n = 5), whereas nerves from vehicle-treated injured animals showed large numbers of fibers with myelin alterations. Compared to vehicle-treated animals, nerves from the DNP 2 group presented a 25% reduction and those from the DNP 4 group presented a 60% reduction in the number of fibers exhibiting myelin alterations.
Expression of APP and neuregulin 1 is preserved in DNP-treated nerves
Neuronal APP mediates cell adhesion to substrate and to glial cells (Breen et al., 1991), and its expression is elevated during degenerative events (Wallace et al., 1993). In peripheral nerves, adhesion to Schwann cells determines the thickness of the myelin sheath (Falls, 2003). A key axonal signaling molecule involved in regulating the thickness of the myelin sheath is neuregulin 1 (Nrg1), a growth factor that anchors to specific ligands in the Schwann-cell membrane (Michailov et al., 2004). Given the important roles played by these proteins in cell survival, adhesion, and myelination, we evaluated APP and neuregulin levels by immunohistochemistry in the proximal segments of both injured and uninjured nerves. Compared to uninjured (contralateral) sciatic nerves, a significant reduction in APP immunoreactivity was observed in injured nerves from vehicle-treated animals 48 h post injury (Fig. 4a,b). Quantitative immunofluorescence analysis showed that injury reduced APP labeling by 42% compared to the uninjured side (Fig. 4g) and administration of DNP prevented the reduction in APP labeling (Fig. 4c–g). In fact, APP immunoreactivities in injured nerves from DNP-treated mice were slightly higher than the levels in uninjured nerves. Uninjured (contralateral) nerves from vehicle-, DNP3-, and DNP5-treated animals presented no significant differences in APP labeling.

APP immunoreactivity in proximal segments of injured sciatic nerves. Panels
Similar results were observed with Nrg1 staining (Fig. 5). Injured nerves presented a 50% decrease in Nrg1 staining compared to uninjured nerves from vehicle-treated animals (Fig. 5g), and this reduction was blocked by DNP treatment.

Neuregulin1 (Nrg1) immunoreactivity in proximal segments of injured sciatic nerves. Panels
As a control, we performed experiments following exactly the procedures described above and immunolabeling nerve sections for neurofilament. Because the proximal segment of the injured nerve does not undergo wallerian degeneration, the axonal cytoskeleton would not be expected to exhibit significant changes under these experimental conditions. Indeed, results showed no alteration in immunostaining for neurofilament, suggesting that changes in APP and Nrg1 immunoreactivities were not part of a global, generalized alteration in staining for proteins in the proximal segment of sciatic nerves (supplementary Fig. 1 [see online supplementary material at
DNP upregulates APP expression in dorsal root ganglia neurons
The cell bodies of sensory neurons whose fibers constitute the sciatic nerve are located in the L4 and L5 dorsal root ganglia (DRG). Because DNP treatment was found to preserve APP expression in the proximal segments of injured nerves, we further sought to examine APP expression in DRG in injured animals.
Figure 6a,b shows typical staining for APP in dorsal root ganglion cells. Neither satellite cells nor Schwann cells presented staining for APP, and similar results were obtained for both L4 and L5 ganglia (data not shown). Cell bodies from both injured and uninjured nerves presented a punctate perinuclear staining pattern for APP, showing that injury itself did not alter the APP immunolabeling pattern after 48 h. DRG neurons from DNP-treated mice showed increased punctate APP staining around the nuclei and, in some cases, close to cell boundaries (Fig. 6c–f), with a more prominent effect in nerves from the DNP4 group than from the DNP2 group.

APP immunoreactivity in dorsal root ganglion neurons. Representative images are shown for APP immunolabeling in neuronal cell bodies in DRG from both injured (
Changes in APP expression were further analyzed by calculating integrated fluorescence intensities in panels containing twenty images per experimental group. Panels g and h show integrated immunofluorescence intensities (normalized by cell numbers) for APP immunostaining in DRG corresponding to injured and uninjured nerves. Compared to vehicle-treated animals, DRG from DNP-treated animals presented significant increases in APP immunoreactivity.
DNP treatment attenuates post-injury decline in limb function
Limb function was assessed following injury using a walking track test (see Methods). Figure 7a shows typical examples of footprints of the left hindlimb from animals belonging to the four experimental groups used in the present study. At 6 weeks post injury, a dramatic reduction was verified in toe spread (i.e., the distance between the first and fifth toes) of the injured limb in vehicle-treated animals, whereas DNP 2- and DNP 4-treated animals presented partial and complete preservation respectively of the distance between toes.
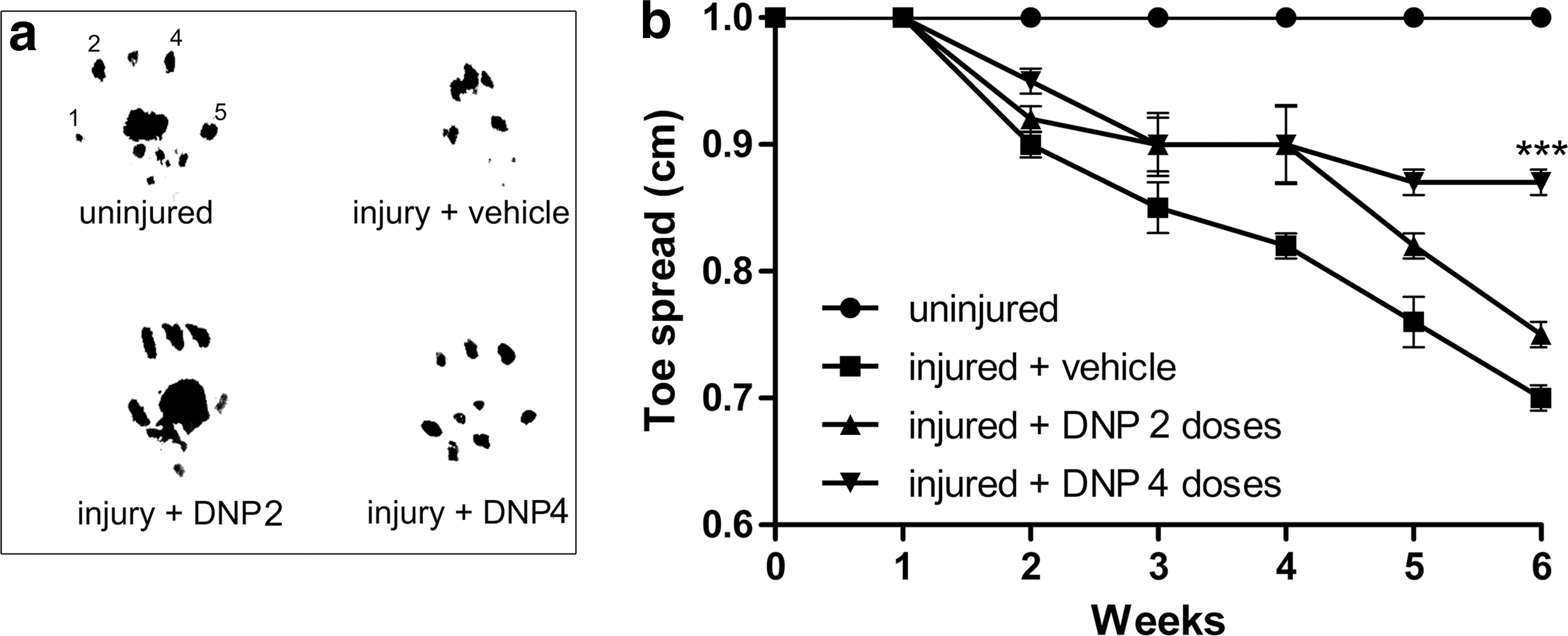
Paw finger-printing and sciatic function assay. Panel (
Measurements of toe spread (specifically the distance between the first and fifth toes) were used as a parameter for evaluation of limb function as previously proposed (Bozkurt et al., 2008). Following sciatic injury, animals were submitted to a walking track test once a week for 6 weeks and their toe spreads were measured (Fig. 7b). As expected, uninjured animals maintained a constant toe spread (averaging 1 cm) during the 6 weeks of investigation. By contrast, vehicle-treated injured animals presented a progressive decrease in toe spread over the course of 6 weeks. Interestingly, injured mice from the DNP2- and DNP4-treated groups exhibited significantly less reduction in toe spread, indicating that DNP treatment attenuated limb functional loss in the weeks following injury.
We note that during the 6 weeks of treatment, DNP-treated animals showed no signs of weight loss (supplementary Fig. 2 [see online supplementary material at
Discussion
Following traumatic nerve injury, the distal segments of injured axons undergo WD. This process can also be triggered by ischemia, metabolic disorders, and neurodegenerative diseases, suggesting a common mechanism underlying these pathologies. Strategies to prevent or block WD could lead to a better understanding of mechanisms of neurodegeneration, ischemia, and trauma of the nervous system and to the development of novel therapeutic drugs for these conditions. Despite this obvious importance, anti-degenerating strategies still constitute a neglected target for pharmacological therapy.
Peripheral nerve injury results in loss of motor and sensory functions. Different strategies have been proposed to prevent degeneration and/or to improve fiber regeneration following injury. Although recent studies have reported promising results, effective anti-degenerative approaches are still lacking. We now demonstrate that DNP blocks sciatic-nerve degeneration following compression injury. We also show that DNP treatment preserves the levels of APP and Nrg1, proteins implicated in neuronal survival and myelination, in injured nerves and protects against loss of limb function following injury.
The first ultrastructural alteration in WD is a progressive disintegration of the cytoskeleton in the distal stump of the injured nerve. The current results show that DNP treatment rescued a significant number of myelinated fibers from undergoing WD. Blockade of cytoskeleton degradation constitutes a possible mechanism for preservation of the axoplasm. We recently reported that DNP increases levels of microtubule-associated protein tau in primary neurons in culture (Wasilewska-Sampaio et al., 2005). Tau stabilizes microtubules by interacting with tubulin to promote its assembly. Indeed, one of the main functions of tau is to regulate the stability of axonal microtubules (Drubin and Kirshner, 1986; Weingarten et al., 1975). It is thus possible that stabilization of axonal cytoskeleton in the nerves of DNP-treated animals is related to increased tau expression.
DNP has also been shown to increase intracellular levels of cAMP and to promote expression of genes related to cAMP signaling pathways in neurons (De Felice et al., 2007; Sebollela et al., 2009; Wasilewska-Sampaio et al., 2005). cAMP is an important cellular second messenger and is involved in many signaling cascades, including cell-survival pathways (Hanson et al., 1998). Interestingly, elevated cAMP levels have been shown to abrogate myelin-MAG inhibition of neurite outgrowth in mature neurons (Cai et al., 2001) and to promote increased neurite extension and neuronal differentiation (Wasilewska-Sampaio et al., 2005). Moreover, increased cAMP levels upregulate expression of APP (R.K. Lee et al., 1997), likely explaining the increase in APP levels in sciatic nerves from DNP-treated animals (see below). These observations suggest that another mechanism by which DNP promotes neuronal survival and preserves axonal ultrastructure may be related to increased cAMP levels.
Although, at high concentrations, DNP is a well-known mitochondrial uncoupler, we have previously demonstrated that at low concentrations (up to 20 μM), DNP does not affect neuronal viability (De Felice et al., 2001) and does not cause mitochondrial uncoupling or interfere with cellular respiration in a neuronal cell line and in hippocampal neurons (Sebollela et al., 2009; Wasilewska-Sampaio et al., 2005). In addition, we have found that 20 μM DNP does not cause any changes in ATP production or mitochondrial redox activity (as measured by MTT reduction) in primary neurons in culture (Sebollela et al., 2009). Moreover, assuming that DNP would be uniformly distributed in the mouse organism, an estimated overall concentration of 0.3 μM DNP can be calculated upon intraperitoneal injection of the doses used in the present study. It seems unlikely, therefore, that this low dose of DNP would result in mitochondrial uncoupling in mice. In this regard, we note that mice treated with IP injections of DNP for up to 6 weeks showed no signs of weight loss or behavioral alterations, which would be expected upon mitochondrial uncoupling.
In the 1930s, DNP was administered to humans as part of weight-reduction programs. However, due to reports of serious side effects including cataracts, gastrointestinal, cardio-respiratory, and other types of complications, even deaths in individuals subjected to high concentrations of DNP, the U.S. Food and Drug Administration (FDA) banned the use of DNP in 1938 (De Felice and Ferreira, 2006; Tainter et al., 1935). As noted above, however, recent studies have demonstrated that low, sub-toxic doses of DNP can have interesting neuroprotective actions against a variety of insults (reviewed in De Felice and Ferreira, 2006; De Felice et al., 2007). The current results, therefore, add to the body of evidence indicating that the dual nature of DNP effects should be carefully reexamined with the goal of determining therapeutically effective dosing regimes that can be safely administered in different conditions.
Results also showed that DNP preserves the expression of APP and Nrg1 in injured sciatic nerves. APP is involved in a number of important neuronal processes including growth, differentiation, and survival (reviewed in Gralle and Ferreira, 2007). Furthermore, APP increases neuronal adhesion to substrate and to glial cells (Breen et al., 1991) and its expression is elevated during degenerative events (Wallace et al., 1993). Preserved APP expression in the proximal segments of injured sciatic nerves and in dorsal root ganglia from DNP-treated animals may help to prevent propagation of degenerative signaling by maintaining and strengthening axon/Schwann-cell interactions.
Preserved Nrg1 expression in DNP-treated animals may be directly related to preservation of conductance (as indicated by the G ratio) in injured nerves. Nrg1 plays important roles in the development of the nervous system and in neurodegeneration (Falls, 2003). One of the most important functions of Nrg1 is in the control of myelin thickness and of the axon/myelin ratio in the PNS (Michailov et al., 2004). Results thus suggest that preservation of APP and Nrg1 expression induced by DNP treatment may play an important role in the stabilization of the cytoarchitecture and function of nerve fibers.
The sciatic function index (SFI) has been described as a reliable indicator of limb functional preservation or recovery (Bain et al., 1989; de Medinaceli et al., 1982). Among several possible ways to evaluate SFI (Bain et al., 1989; de Medinaceli et al., 1982; Dijkstra et al., 2000), we have used toe-spread measurements, which show the highest correlation with any other functional recovery index (Bozkurt et al., 2008). Results showed that DNP treatment significantly attenuated the decrease in SFI that takes place over the course of several weeks post injury. Corroborating the SFI results, we also found that DNP-treated animals presented normal G-ratio values, similar to uninjured control animals.
In conclusion, DNP prevents sciatic nerve degeneration after a crush injury by blocking cytoskeleton degradation and by preserving the expression of APP and Nrg1, proteins that play key roles in neuronal survival and myelination. DNP treatment also prevented mitochondrial abnormalities and limb functional loss following injury. These results argue that DNP can be used as a tool in the development of novel therapeutic approaches for treatment of nerve trauma and neurodegenerative diseases.
Footnotes
Acknowledgments
We thank Ms. Diana Pontes for her contribution to morphometry and immunofluorescence experiments, Dr. Paulo R. Louzada for suggestions and valuable insight into the nerve injury model and Dr. Fernanda G. De Felice for critical reading of this manuscript. This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro and Instituto Nacional de Neurociência Translacional.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.