
Abstract
Spinal cord injury (SCI) is generally associated with a rapid and significant decrease in muscle mass and corresponding changes in skeletal muscle properties. Although β2-adrenergic and androgen receptor agonists are anabolic substances clearly shown to prevent or reverse muscle wasting in some pathological conditions, their effects in SCI patients remain largely unknown. Here we studied the effects of clenbuterol and testosterone propionate administered separately or in combination on skeletal muscle properties and adipose tissue in adult CD1 mice spinal-cord-transected (Tx) at the low-thoracic level (i.e., induced complete paraplegia). Administered shortly post-Tx, these substances were found to differentially reduce loss in body weight, muscle mass, and muscle fiber cross-sectional area (CSA) values. Although all three treatments induced significant effects, testosterone-treated animals were generally less protected against Tx-related changes. However, none of the treatments prevented fat tissue loss or muscle fiber type conversion and functional loss generally found in Tx animals. These results provide evidence suggesting that clenbuterol alone or combined with testosterone may constitute better clinically-relevant treatments than testosterone alone to decrease muscle atrophy (mass and fiber CSA) in SCI subjects.
Introduction
Some anabolic substances are known to prevent extended body weight loss and muscle wasting in different disorders (Dudgeon et al., 2006; Koopman et al., 2009; Shahidi, 2001). Among these substances, clenbuterol, a β2-adrenergic agonist generally used to treat asthma (Anderson and Wilkins, 1977), has been reported to stimulate muscle growth (Carter et al., 1991; Choo et al., 1992; Lynch et al., 1999; MacLennan and Edwards, 1989) and to prevent muscle atrophy in dystrophic, hindlimb unloaded, or nerve-transected models (Agbenyega et al., 1995; Herrera et al., 2001; Teng et al., 2006; Zeman et al., 2000). Interestingly, we recently found that a significant decrease in testosterone level occurs shortly after spinal cord transection, suggesting a role for this hormone in the rapid muscle mass changes seen after SCI (Rouleau et al., 2007). Testesterone is well known for its muscle-building effects in athletes and normal individuals (Choong et al., 2008; Bhasin et al., 1996; Graham et al., 2008), as well as for its role in preventing muscle wasting in different pathological conditions such as in burn patients or HIV patients (Dudgeon et al., 2006; Ferrando et al., 2001).
These two anabolic substances may thus be of special interest to SCI patients for their muscle-building properties, as well as for their potential neuroprotective effects, as recently suggested by data from CNS injured and SCI animal models (Brown et al., 1999; Junker et al., 2002; Kujawa et al., 1989; Zeman et al., 1999, 2004; Zhu et al., 2001). Indeed, neuroprotection has been proposed as the mechanism behind the functional recovery induced by clenbuterol or testosterone in incomplete SCI animals. However, it remains unknown if these anabolic substances can also enable locomotor recovery in a complete spinal-cord-transected (Tx) animal, by facilitating locomotor network reorganization at the spinal level. Moreover, to our knowledge, their effects on muscle atrophy and adipose tissue (used separately or in combination) have never been compared in an animal model of SCI.
To investigate these questions, we assessed the effects induced by clenbuterol, testosterone, or both on body weight, skeletal muscle and fiber size, fiber phenotype distribution, adipose tissue amount, and locomotor function recovery in low-thoracic Tx mice, a reliable experimental model of motor-complete paraplegia (Landry et al., 2004; Lapointe et al., 2006; Ung et al., 2007).
Methods
Animals and surgical procedures
All experimental procedures were conducted in accordance with the Canadian Council for Animal Care guidelines, and were accepted by the Laval University Animal Care and Use Committee. Eight-week-old male CD1 mice (n = 159; Charles River Canada, St. Constant, Quebec, Canada) initially weighing 30–35 g were used for this study. Preoperative care included subcutaneous injections of an analgesic (0.1 mg/kg buprenorphine), an antibiotic (5 mg/kg enrofloxacin) and lactated Ringer's solution (1 mL). All surgical procedures were performed under aseptic conditions. The mice were anesthetized with 2.5% isoflurane. A small incision was made on their back to expose some thoracic spinal segments. The spinal cord was then completely transected inter-vertebrally with microscissors inserted between the 9th and 10th thoracic vertebrae (Lapointe et al., 2006; Ung et al., 2007). To ensure that complete Tx was achieved, the inner vertebral walls were explored and scraped several times with small scissor tips. The opened skin area was sutured and the animals were placed on heating pads for a few hours. The mice were left in their cage with food and water ad libitum. Postoperative care provided for 4 days included subcutaneous injections of buprenorphine (0.2 mg/kg/d), enrofloxacin (5 mg/kg/d), and lactated Ringer's solution (2 × 1 mL/d). Their bladders were manually expressed twice daily for the first week post-Tx and once daily thereafter. Complete Tx was confirmed by (1) initial full paralysis of the hindlimbs, (2) postmortem visual and microscopic examination of the spinal cord lesion, and (3) histological assessment of coronal or mid-sagittal spinal cord sections stained with luxol fast blue/cresyl violet for myelinated axons and Nissl substance, respectively.
Groups and treatments
Tx mice were randomly divided into four main groups: (1) vehicle-treated (control Tx), (2) clenbuterol hydrochloride–treated, (3) testosterone propionate–treated, and (4) clenbuterol hydrochloride + testosterone propionate–treated. In each main group, the animals were subdivided into four subgroups (treatment durations), those treated daily (first treatment given 1 h post-surgery) for 1, 2, 4, or 8 weeks prior to sacrifice. Clenbuterol hydrochloride and testosterone propionate were subcutaneously injected at 1 mg/kg/d (Zeman et al., 2000), and 5 mg/kg/d (Pansarasa et al., 2002), respectively. Clenbuterol hydrochloride and testosterone propionate, purchased from Sigma Chemical Co. (St. Louis, MO), were dissolved in saline, and 0.5% benzyl alcohol + vegetable oil, respectively. Corresponding vehicle-treated groups were injected either with saline or 0.5% benzyl alcohol + vegetable oil. Since data from saline- or oil-treated animals were not significantly different, they were subsequently pooled for statistical analytical purposes. Also, 10 uninjured non-treated mice were used for body weight comparison at 8 weeks.
Body weight and hindlimb movement recovery
During the first week the animals were weighed daily, and weekly thereafter. One day prior to sacrifice, the mice were placed on a motor-driven treadmill (8–10 cm/sec) and locomotor function recovery was assessed using two complementary methods. Each assessment method was conducted for 2 min. For the first assay, we used a semi-quantitative method called average combined score or ACOS, that was developed for assessment of hindlimb movements in Tx rodents (Guertin, 2005; Lapointe et al., 2006; Ung et al., 2007). ACOS is composed of non-locomotor movement (NLM; number/min), locomotor-like movement (LM; number/min), and amplitude. Values are arithmetically combined as follows: [NLM + (2 × LM)] × amplitude. One LM was defined as a flexion-extension movement occurring bilaterally in alternation. We decided to exclude LM occurring during bowel movements to avoid taking into account sacral reflex-induced LM (Strauss and Lev-Tov, 2003). One NLM was defined as a non-bilaterally alternating movement, including movements such as jerks, fast paw shaking, and twitches. Amplitude was characterized by assigning one of three values: 0, no movement; 1, movements that were less than half the range of motion of normal steps; or 2, movements that were greater than half the range of motion of normal steps. For the second assay we used a method referred to as Antri, Orsal, and Barthe or AOB, that was specifically designed for complete Tx rodents (Antri et al., 2002). In brief, the AOB scale consists of 22 scores: 0, no movement; 1, weak limb jerks; 2, weak rhythmic movements with no bilateral alternation; 3, large rhythmic movements with no bilateral alternation; 4, weak rhythmic movements with occasional bilateral alternation; and 5, large rhythmic movements with occasional bilateral alternation. For additional levels, see Antri and colleagues (2002). The ACOS and AOB scales were preferred over other methods such as the Basso mouse scale (Basso et al., 2006), based on results from our laboratory demonstrating greater sensitivity and specificity in completely spinal-cord-transected mice (Ung et al., 2007).
Tissue collection and measures
Upon sacrifice, extensor digitorum longus (EDL) and soleus muscles were dissected and weighed. Area-specific adipose tissues were collected from the abdominal subcutaneous, inguinal, and visceral regions. Seminal vesicles were removed, weighed, and used as positive control tissues for testosterone efficacy.
Immunohistochemistry
Upon dissection, the soleus and EDL were frozen in melting isopentane and stored at −80°C until use. Then 10-μm-thick serial sections from the mid-belly were cut with a cryostat maintained at −20°C (2800E Frigocut; Leica Instruments, Nussloch, Germany) and mounted on Superfrost® plus glass slides (VWR Canlab, Mississauga, Ontario, Canada). Sections were incubated for 2 h in either myosin heavy chain slow or fast (MHCs or MHCf ) primary antibodies, specific for MHC isoform type I and isoform type II, respectively (Vector Laboratories, Burlingame, CA; dilution 1:50 in 0.1 M PBS containing 1% rabbit serum and 1% Triton X-100). For controls, some sections were not incubated with the corresponding primary antibodies. The sections were then rinsed in 0.1 M PBS before incubation with goat anti-mouse IgG (H + L) Alexa Fluor 488 secondary antibody (Molecular Probes, Eugene, OR; dilution 1:500 in 0.1 M PBS containing 1% rabbit serum and 1% Triton X-100). The slides were then rinsed in 0.1 M PBS and mounted with PBS-glycerol (50:50). Immunofluorescence labeling was visualized with an Olympus BX61WI confocal microscope using a 10 × water immersion objective. Images were captured using FluoView 300 (Olympus Canada Inc., Markham, Ontario, Canada) and analyzed with ImageJ software (ImageJ 1.40; Research Services Branch, National Institutes of Health, Bethesda, MD). Analyses consisted of determining muscle fiber phenotype, measuring corresponding cross-sectional area (CSA; averages were from 50 fibers per fiber type/muscle, when available), and calculating relative fiber type distribution per muscle. Type I fibers displayed labeling only with MHCs antibodies, and type II fibers displayed labeling only with MHCf antibodies (no distinction was made between type IIa, IIx, or IIb isoforms), whereas hybrid fibers displayed labeling with both antibodies (adjacent sections).
Statistical analysis
Differences between groups for locomotor recovery, adipose tissue, muscle mass, fiber CSA and fiber type distribution were analyzed using a one-way analysis of variance (ANOVA), followed by a Bonferroni post-hoc test. Body weight changes were analyzed using a two-way repeated-measures ANOVA, followed by a Bonferroni post-hoc test. Results are reported as mean ± SEM.
Results
Prior to assessing the effects of anabolic treatments on muscles, testosterone bioavailability and efficacy had been confirmed in standard tissue controls (testosterone-induced seminal vesicle hypertrophy). The results showed that testosterone with or without clenbuterol significantly (p < 0.05) increased by 35% on average seminal vesicle size and weight compared with control Tx or clenbuterol-treated animals (data not shown).
Body weight
As previously shown elsewhere, a significant loss in body weight is normally found within the first week post-Tx in paraplegic animals (Landry et al., 2004; Ung et al., 2007). This finding was again confirmed in the present study. After combining data from all subgroups, Figure 1A reveals that only vehicle-treated Tx mice (control Tx) displayed a significantly greater decrease in body weight, peaking (84.1 ± 0.9% of initial body weight) at 5 days post-Tx (in Fig. 1A the solid black line includes control Tx mice from all four subgroups). Tx animals treated with clenbuterol (solid grey line), testosterone (dashed grey line), or both (dashed black line) also displayed significant body weight loss that reached its peak (88–89% of initial body weight) at day 3 post-Tx (Fig. 1A). Thus the extent of body weight loss in control Tx animals was significantly (p < 0.05) greater at 3, 5, and 7 days post-Tx compared with treated animals. For Figure 1B–E, only mice that were sacrificed on that specific week were taken for analysis. Data from each of the four main groups (treatments and control Tx) examined individually showed decreased body weight loss and increased body weight values at 1, 2, 4, and 8 weeks post-Tx (Fig. 1B–E). Differences that reached significant levels (p < 0.05) were found specifically at 4 and 8 weeks post-Tx in clenbuterol-treated (105% of initial body weight), and clenbuterol + testosterone-treated mice (103% of initial body weight), compared with vehicle-treated mice and testosterone-treated animals (Fig. 1D–E). At 8 weeks, uninjured mice showed an average body weight of 122.4 ± 2.2%, which represented a 2.8 % increase per week (Fig. 1F). Conversely, after Tx, the control Tx (1.4%), clenbuterol (1.7%), testosterone (1.5%) and clenbuterol + testosterone (1.8%) groups showed lower average body weight gain per week. These results show that comparable short-term effects (i.e., within 1 week post-Tx) on body weight were induced by all three treatments, although only clenbuterol or clenbuterol +testosterone induced significantly increased greater body weight at 4 and 8 weeks post-Tx.
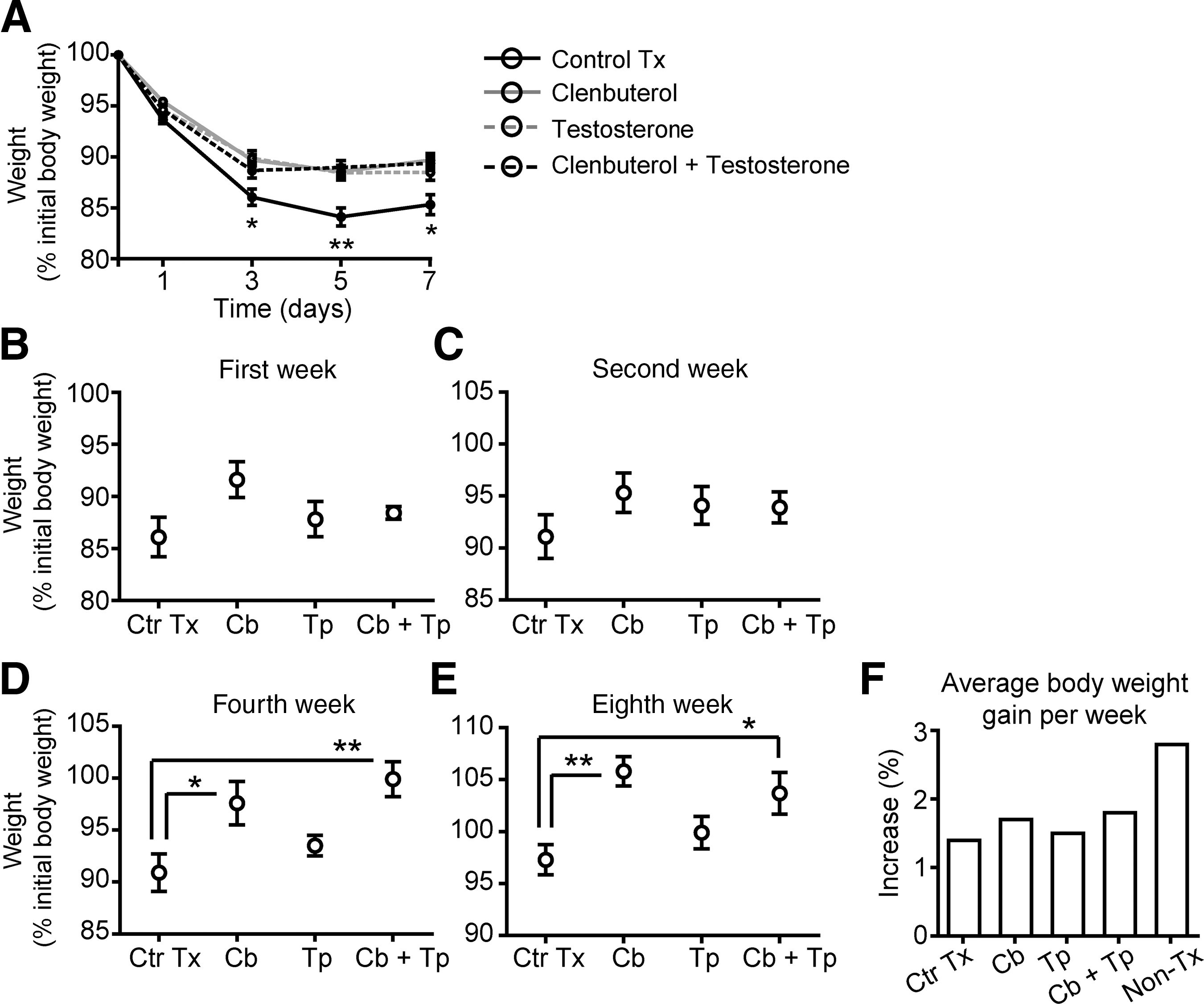
(
Muscle mass
The soleus and EDL were dissected out and weighed. When comparing subgroups (treatment durations), soleus values were significantly (p < 0.05) greater in clenbuterol-treated animals at 4 and 8 weeks (0.0244 ± 0.0024% and 0.0207 ± 0.0017% of body weight, respectively), than at 1 week (0.0158 ± 0.009% of body weight; Fig. 2AII). Comparable results were found with soleus in testosterone-treated or clenbuterol + testosterone-treated animals with significant (p < 0.01) increases at 4 and 8 weeks post-Tx, but not at 1 week (Fig. 2AIII and 2AIV). The greatest significant (p < 0.05) increase of soleus mass, which was revealed by comparing treatments, was identified in clenbuterol + testosterone-treated animals, with values reaching 0.0265 ± 0.0020% of body weight (Table 1).
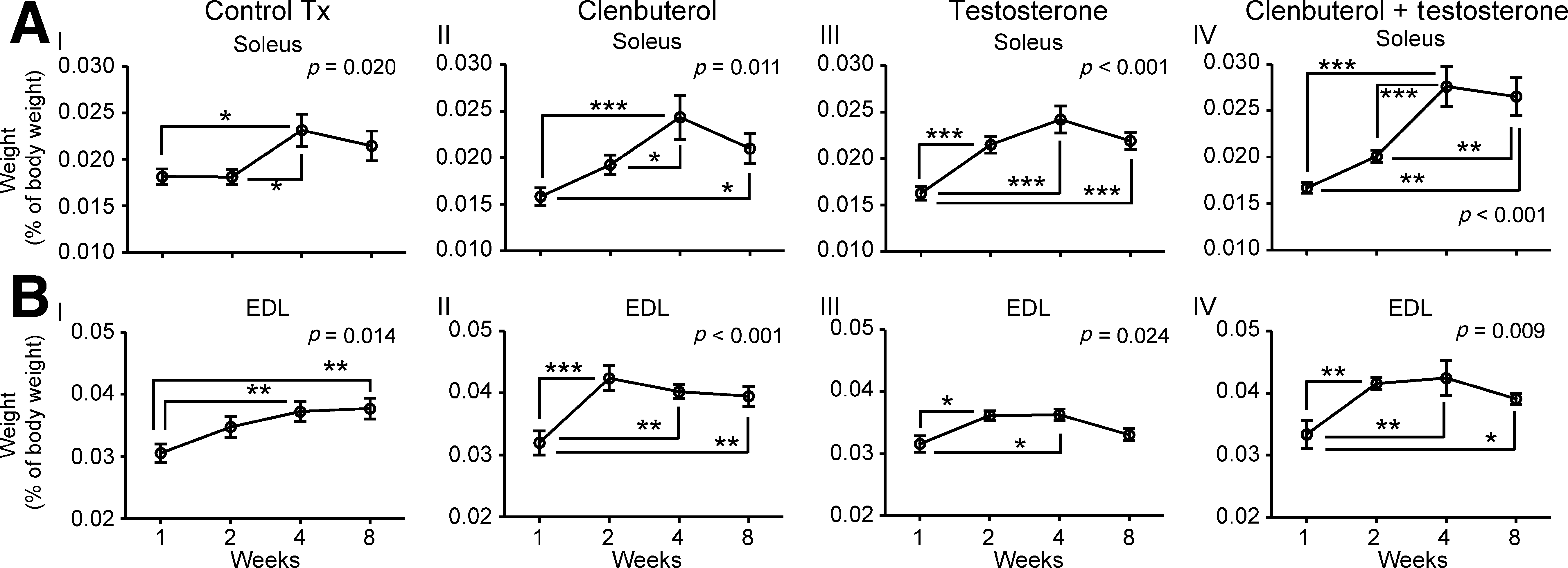
Soleus (
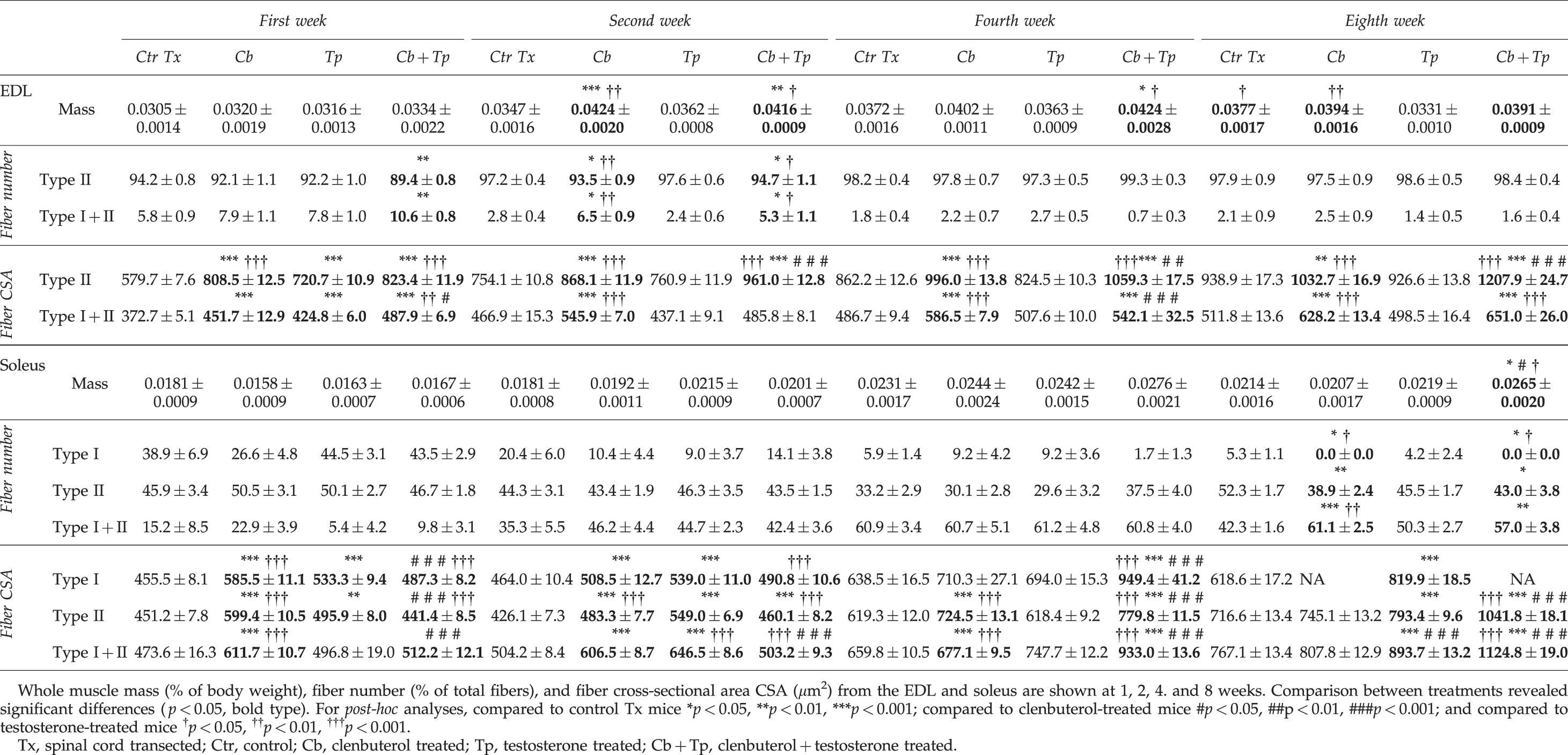
EDL also increased in mass with time in all subgroups, as shown in Fig. 2BI–IV. In contrast to the soleus, EDL masses were significantly higher in both clenbuterol + testosterone-treated (0.0416 ± 0.0009% of body weight) and clenbuterol-treated (0.0424 ± 0.0020% of body weight) animals compared with control Tx animals at 2 weeks (0.0347 ± 0.0016% of body weight; Table 1). This indicates that comparable effects on EDL mass were induced by clenbuterol alone and clenbuterol +testosterone.
Extensor digitorum longus and soleus fiber distribution
We also studied potential changes in fiber type conversion, since a shift from slow-twitch to fast-twitch fibers (e.g., type I → hybrid fibers → type II fibers) is generally known to occur after spinal cord transection (Lieber et al., 1986a,b).
The results showed that significantly (p < 0.001) fewer hybrid fibers were found in the EDL with time (Fig. 3B). Consequently, this was associated with a progressive increase in type II fibers (Fig. 3A). No type I fibers were found in the EDL. When comparing treatments, we found significantly (p < 0.05) greater percentages of hybrid fibers in clenbuterol + testosterone-treated mice than in control Tx animals at 1 week and 2 weeks (Table 1). At 2 weeks, greater percentages of hybrid fibers were also found in clenbuterol-treated animals compared with testosterone-treated or vehicle-treated mice. However, at 4 and 8 weeks, only low percentages of hybrid fibers (<3% of the total fiber number) were found, with no significant difference between groups. In contrast, percentages of type II fibers kept increasing over time, with relatively high values seen at 8 weeks that ranged from 97.5 ± 0.9 to 98.6 ± 0.5 % of the total fiber number (Table 1). Results in this section showed that no long-term difference between treatments was found.
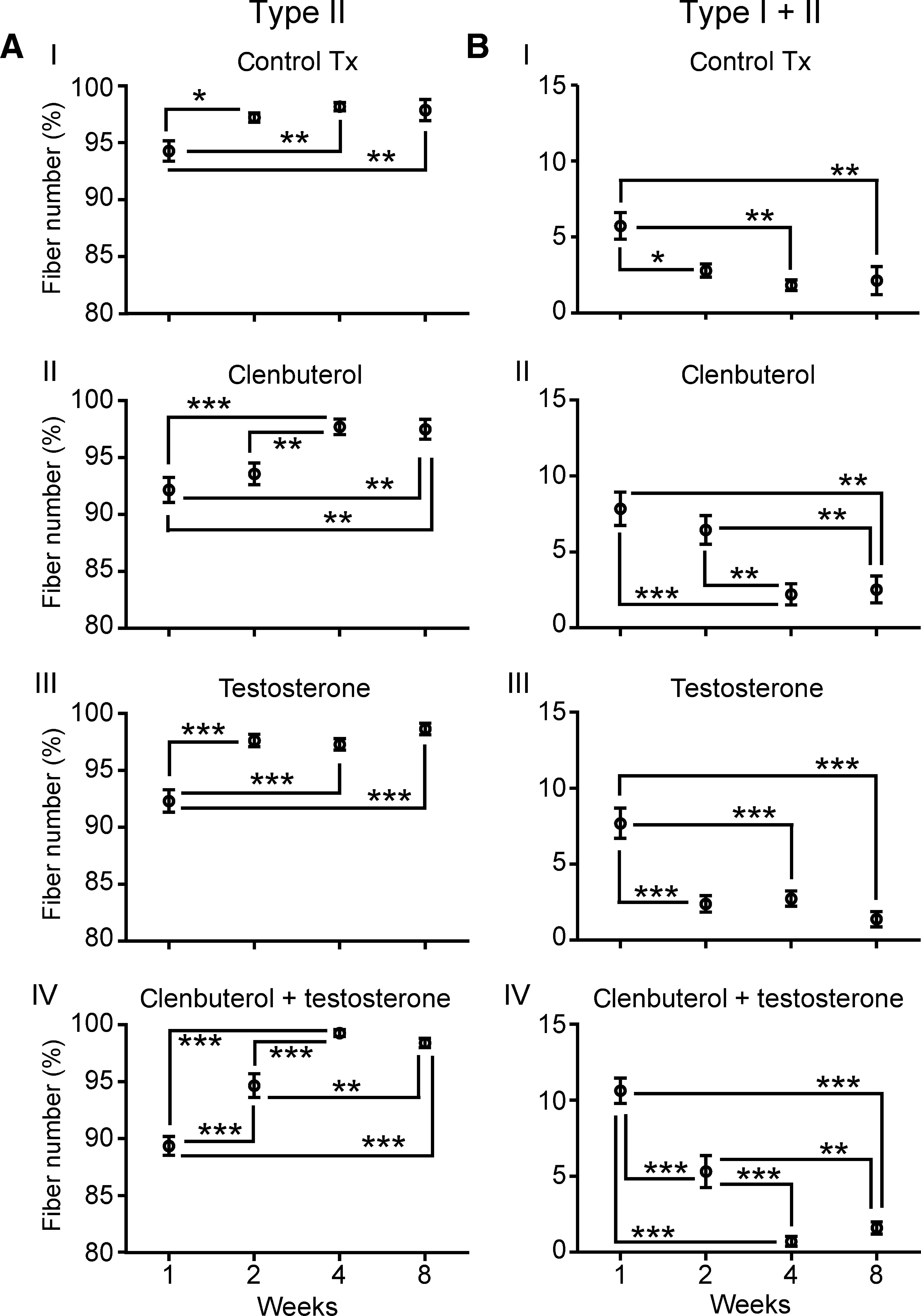
Extensor digitorum longus (EDL) fiber phenotype distribution. Comparisons are between subgroups (treatment durations). (
In the soleus, where all three main types of fibers (type I, type II, and hybrid) were found, the results also showed significant changes in fiber type conversion, although they were different than those seen in the EDL. Across all treatments, percentages of type I fibers ranging from 26.6 ± 4.8% to 44.5 ± 3.1% of total fiber number at 1 week were found to decrease to values ranging between 0 and 5.3 ± 1.1% by 8 weeks (Fig. 4AI–AIV and Table 1). Type II fiber percentages were found to remain relatively unchanged, between 40% and 50%, when weeks 1 and 8 were compared. However, at 4 weeks a transient decrease in percentage was found in all groups except the clenbuterol + testosterone-treated mice (Fig. 4BI–BIV). In clear contrast, percentages of hybrid fibers were found to increase with time in all groups (generally from 5–20% up to 40–60% of total fiber number; Fig. 4CI–CIV). When comparing groups (treatments), differences were only found at week 8 for all fiber types. The testosterone-treated group had a greater percentage of type I fibers than the clenbuterol-treated and clenbuterol + testosterone-treated mice. Greater percentages of type I and type II fibers were also found in vehicle-treated groups than in clenbuterol-treated and clenbuterol + testosterone-treated mice. In contrast, more hybrid fibers were labeled in clenbuterol-treated and clenbuterol + testoterone-treated mice (Table 1). Data in this section show that none of the treatments could prevent the drastic loss of type I fibers, or the significant increase in type I + II (hybrid) fibers. This said, clenbuterol and clenbuterol +testosterone further exacerbated the loss of type I fibers, which completely disappeared (none were labeled at 8 weeks).
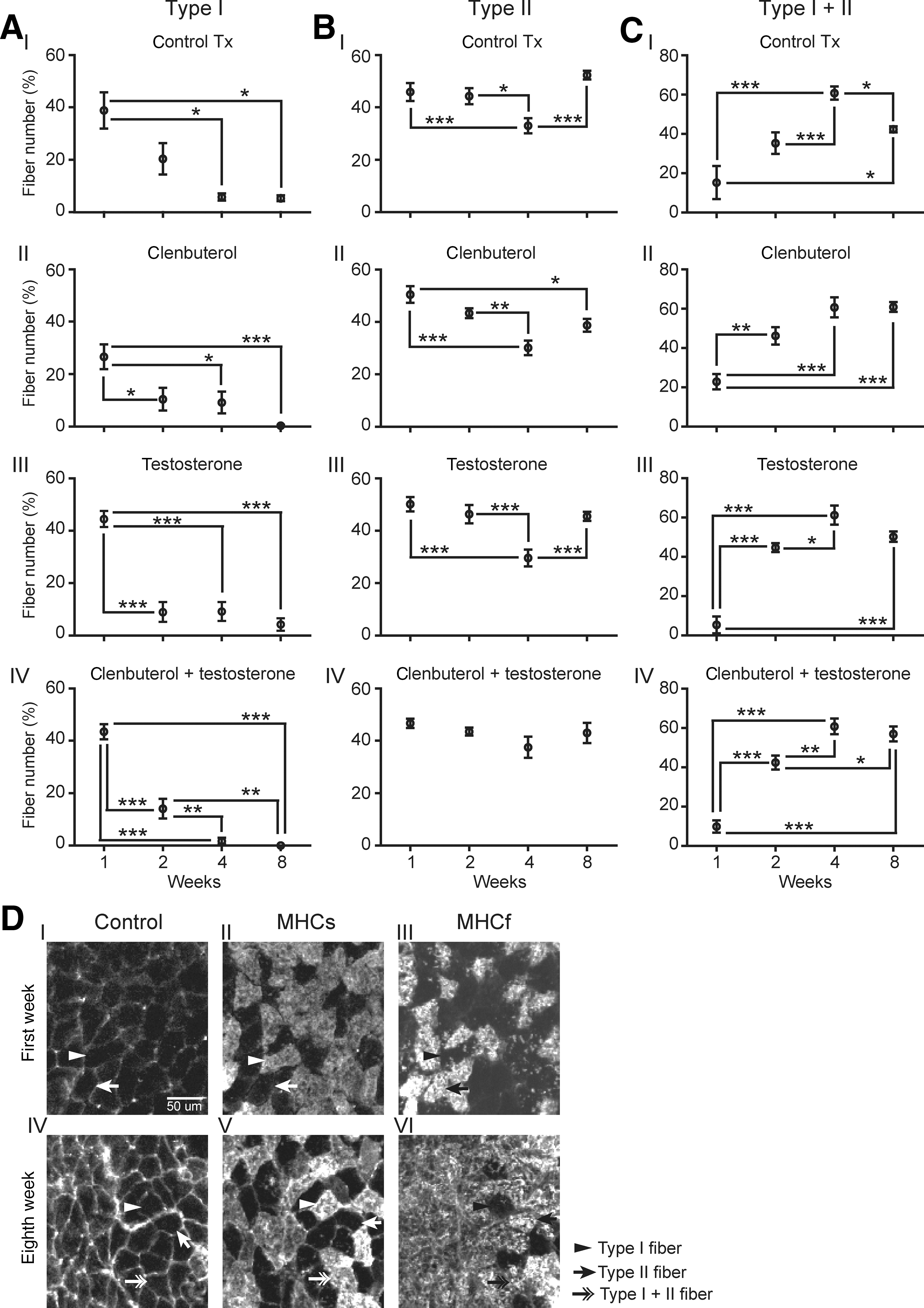
Soleus fiber phenotype distribution. Comparisons are between subgroups (treatment durations). (
Extensor digitorum longus and soleus fiber cross-sectional area
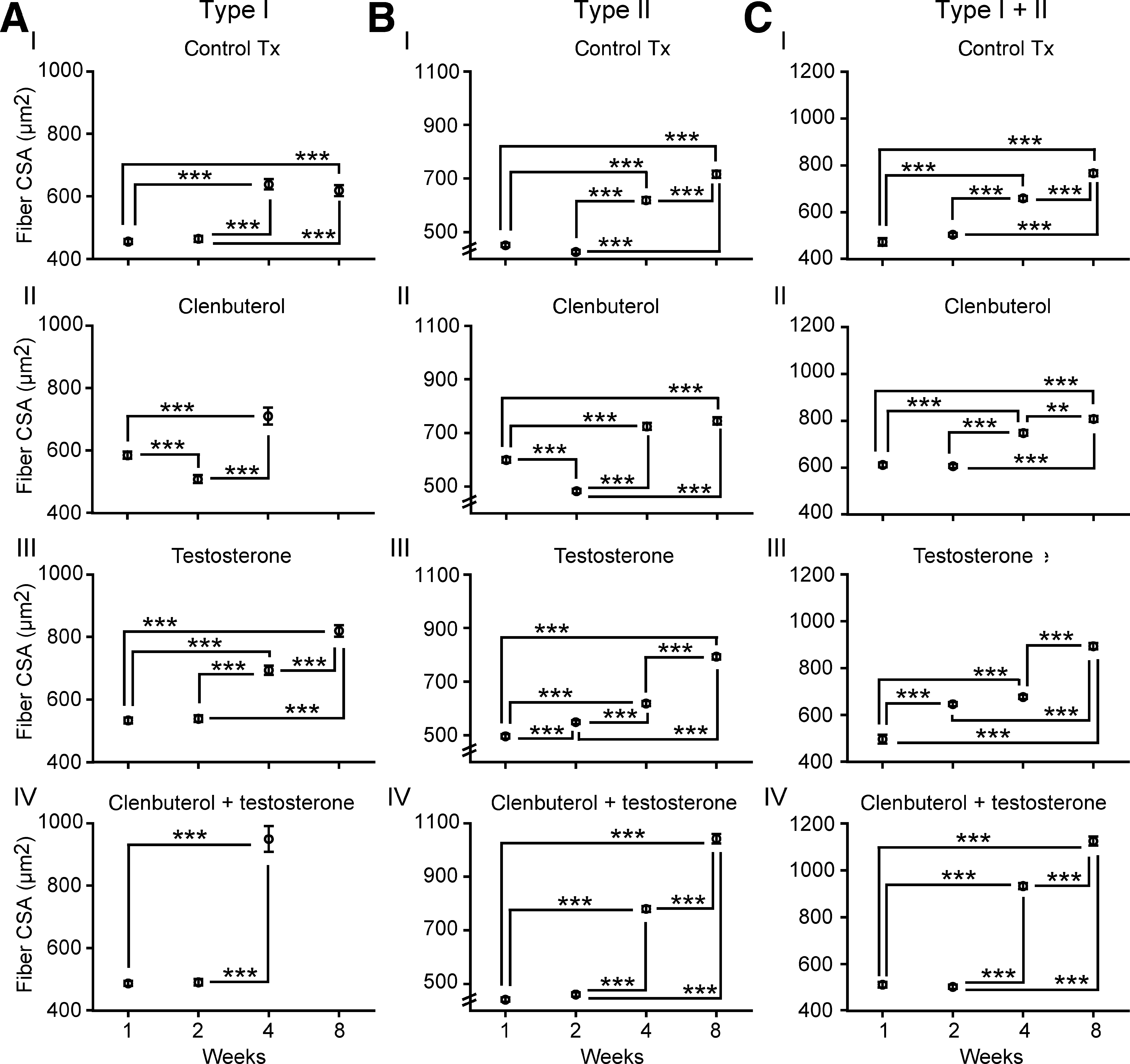
The results showed, in all subgroups, a progressive increase of EDL fiber CSA values (Fig. 5). In vehicle-treated animals, type II fiber CSA values went from 579.7 ± 7.6 μm at 1 week up to 938.9 ± 17.3 μm at 8 weeks (Fig. 5AI). Type II fiber CSA values also progressively increased in clenbuterol-treated, testosterone-treated, and clenbuterol + testosterone-treated animals, with values that reached 1032.7 ± 16.9 μm, 926.6 ± 13.8 μm, and 1207.9 ± 24.7 μm at 8 weeks, respectively (Fig. 5AII–AIV). Comparable changes were found in hybrid fiber CSAs, with values lower than 400 μm in vehicle-treated animals, and up to 650 μm in clenbuterol-treated or clenbuterol + testosterone-treated mice (Fig. 5BI–BIV). When comparing treatments at 8 weeks, the results revealed that greater (p < 0.001) augmentations of type II fiber CSA values were found in clenbuterol + testosterone-treated mice than in the other groups (Table 1). Clenbuterol-treated mice also showed greater type II fiber CSA values than testosterone-treated and control Tx animals. For hybrid fibers, CSA values were not significantly different between the clenbuterol-treated and clenbuterol + testosterone-treated groups. However, both groups showed significantly greater CSA values (p < 0.001) than the vehicle-treated or testosterone-treated mice (Table 1). These data show that clenbuterol and clenbuterol + testosterone induce greater effects on EDL CSA values.
Extensor digitorum longus fiber cross-sectional area (CSA) values. Comparisons are between subgroups (treatment durations). (
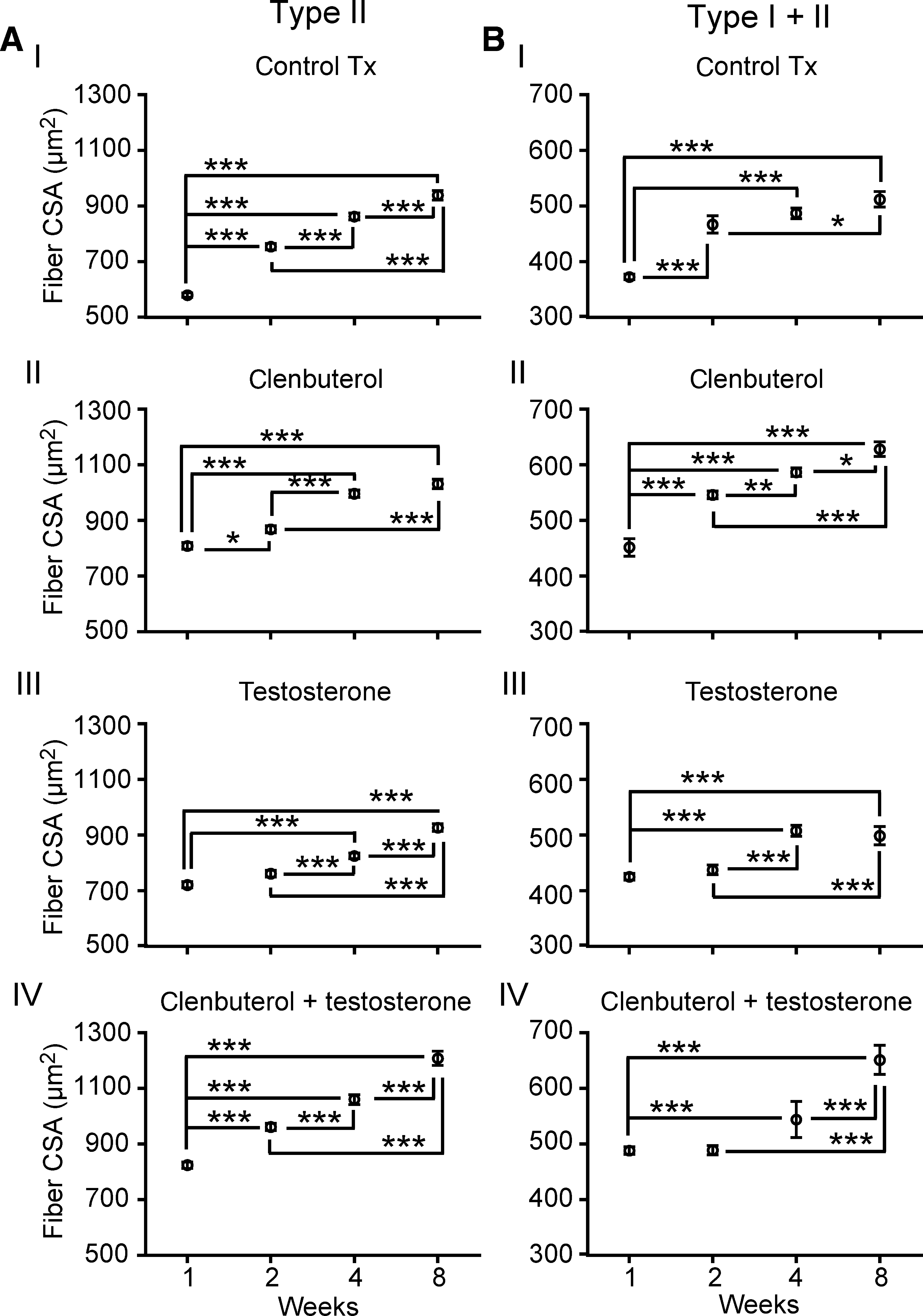
For the soleus, CSA values were also found to generally increase with time, except for type I fibers in clenbuterol-treated and clenbuterol + testosterone-treated animals, for which at 8 weeks, no more type I fibers were found, as mentioned previously (Fig. 6AII and IV and Table 1). In vehicle-treated animals, type I fiber CSA values ranged around 450 μm (at 1 week) and 620 μm (at 8 weeks), whereas in clenbuterol-treated or clenbuterol + testosterone-treated mice, values ranged between 485 μm and 950 μm (e.g., at 4 weeks in clenbuterol + testosterone-treated animals; Fig. 6AIV). When comparing groups, we found that greater augmentations were seen at 4 weeks in clenbuterol + testosterone-treated animals compared with the other groups (p < 0.001; Table 1), and at 8 weeks in testosterone-treated compared with vehicle-treated mice (p < 0.001; Table 1). Regarding type II and hybrid fiber types, CSA values generally increased over time (Fig. 6). Comparisons between treatments showed significantly greater augmentations at 8 weeks in clenbuterol + testosterone-treated mice (approximately 1100 μm) than in the other groups (Table 1). Data in this section show that greater effects on soleus fiber size were induced by clenbuterol + testosterone than in the other groups.
Soleus fiber cross-sectional area (CSA) values. Comparisons are between subgroups (treatment durations). (
Adipose tissue
Adipose tissues were collected from specific regions of the body to study the extent to which these treatments could selectively alter fat storage in Tx mice (Table 2). In brief, fat tissue amounts were generally reduced in the clenbuterol and clenbuterol + testosterone groups than in the vehicle-treated mice. This was found at all time points, although significant levels were reached only at 2 and 4 weeks (e.g., levels were reduced from 2.81% to less than 2.0% of body weight at 4 weeks). At 4 weeks, compared to control Tx mice, testosterone-treated mice also had less adipose tissue. No significant difference was found between treatments at 8 weeks. In brief, no trend or clear region-specific loss was identified.
Adipose tissue results expressed in percentage of body weight. Significant differences (bold) were only noted for total adipose tissue, as all specific areas showed similar differences. For post-hoc analyses, compared to control Tx mice *p < 0.05, **p < 0.01; compared to clenbuterol-treated mice ††† p < 0.001.
Tx, spinal cord transected; Ctr, control; Cb, clenbuterol treated; Tp, testosterone treated; Cb + Tp, clenbuterol + testosterone treated.
Hindlimb motor recovery
We found in all groups a significant increase with time of motor/locomotor scores (i.e., involuntary hindlimb movements) as assessed with the ACOS and AOB scales (p < 0.05), both of which are designed to assess hindlimb movements in completely spinal-cord-transected animals. ACOS scores ranged between 0 and 0.1 ± 0.1 at 1 week, and increased to scores ranging between 8.3 ± 3.6 and 14.9 ± 3.8 at 8 weeks (Table 3). Comparable results were found with the AOB scale, with scores ranging between 0 and 0.2 ± 0.1 at 1 week, that increased up to scores ranging between 2.0 ± 0.6 and 3.1 ± 0.6 at 8 weeks (Table 3). However, no significant (p > 0.05) difference was found between treatments.
All groups showed significant recovery from the first to the eighth week (p < 0.05). No difference between treatments was found after 8 weeks.
Tx, spinal-cord-transected; ACOS, average combined score; AOB, Antri, Orsal, and Barthe.
Discussion
This study is the first to directly assess under comparable conditions, the effects of testosterone, clenbuterol, or both, in the same animal model of SCI. The results clearly show that (1) clenbuterol and clenbuterol + testosterone (but not testosterone alone) induced significantly greater body weight increases at 4 and 8 weeks post-Tx; (2) greater effects were induced by clenbuterol + testosterone on soleus mass than with testosterone or control Tx; (3) greater effects were elicited by clenbuterol alone and clenbuterol + testosterone on EDL mass than testosterone or control Tx; (4) no difference between treatments was found for EDL fiber type conversion after 8 weeks; (5) no treatment prevents or reverses the drastically reduced proportion of soleus type I fibers and the increased proportion of soleus hybrid fibers (although clenbuterol and clenbuterol + testosterone exacerbated the loss of type I fibers, and induced increased hybrid fiber phenotype); (6) clenbuterol and clenbuterol + testosterone induced greater effects on EDL CSA values; (7) greater effects on soleus CSA values were induced by clenbuterol + testosterone; (8) the loss of fat tissue was generally greater in all treated groups than in control Tx animals; and (9) the progressive development of involuntary motor/locomotor hindlimb movements was not altered by these treatments.
In other words, testosterone did not generally alter most changes normally found in SCI mice, although it significantly increased fat tissue loss, as did the other treatments. In contrast, clenbuterol reduced and reversed body weight loss (although it decreased adipose tissue storage), and reduced EDL muscle mass and fiber atrophy. Also greater effects, although not often significantly greater than clenbuterol alone, were generally induced by clenbuterol + testosterone, which decreased and reversed body weight loss, and reduced EDL and soleus muscle mass, as well as fiber atrophy.
Effects of anabolic treatments on overall body composition
Drastic changes in overall body composition are generally known to occur after SCI. For instance, a rapid decrease in body weight was reported in both SCI patients (Cox et al., 1985) and Tx mice (Landry et al., 2004). Here, although all three treatments reduced body weight loss within the first week post-Tx, only clenbuterol and clenbuterol + testosterone increased body weight gain after 1 or 2 months of treatment (e.g., to 105% of initial body weight in clenbuterol-treated animals). This effect on body weight was most likely attributed to corresponding changes in muscles rather than fat tissues, since clenbuterol and clenbuterol + testosterone both largely increased skeletal muscle mass and muscle fiber CSA values, whereas fat tissue amounts decreased with all treatments. It has been suggested that testosterone and clenbuterol induce proliferation, differentiation, and recruitment of satellite cells into muscle fibers, to promote muscle hypertrophy or inhibit muscle atrophy (Sinha-Hikim et al., 2002; Spurlock et al., 2006). However, the mechanisms underlying this effect on muscle atrophy/hypertrophy are not completely understood, but regulation of protein synthesis and breakdown may include inhibition of ubiquitin-proteasome pathways, inhibition of 3-methylhistidine, and upregulated expression of MAFbx, as was shown in other models of unloading (Benson et al., 1991; Yimlamai et al., 2005; Zhao et al., 2008 a,b; for reviews see Herbst and Bhasin, 2004 and Lynch and Ryall, 2008).
This said, these drugs could have decreased weight loss through actions on other systems such as on bones (not assessed in this study), which are known to undergo extensive tissue loss after SCI in both humans and rodents (Modlesky et al., 2004; Zehnder et al., 2004). In fact, in Tx mice, Picard and colleagues (2008) reported a rapid (within 1 month post-Tx) 22% and 14% reduction of femoral bone volume and femoral bone mineral content, respectively. In accordance with this, clenbuterol was reported to prevent bone tissue loss under conditions of disuse, such as in the denervated hindlimb or tail-suspended model (Apseloff et al., 1993; Bloomfield et al., 1997; Zeman et al., 1991). There is also evidence suggesting that testosterone can prevent bone tissue loss in rats (Stuermer et al., 2009), and increase bone density in hypogonadal men (Katznelson et al., 1996; Snyder et al., 2000). Therefore, clenbuterol- and clenbuterol + testosterone-induced effects on body weight may have also been partially caused by protecting against bone tissue loss in Tx animals.
Clenbuterol and testosterone are also well known to increase lean body mass through lipolytic actions on fat tissue (Katznelson et al., 1996; Kearns et al., 2001). Results of this study may suggest lipolytic effects induced in Tx animals, since amounts of specific fat tissues were reduced by all treatments, especially on week 4. This is of particular clinical interest, specifically in late chronic SCI patients (>1 year post-SCI), since increased fat tissue and obesity are often seen several months to several years post-trauma (Cox et al., 1985). This said, we cannot conclude from our data that lipolytic effects were found in these animals, since caloric intake or recompartmentalization of adipose tissues has not been examined by dual x-ray absorptiometry.
Effects on muscle fiber type distribution and size
To our knowledge, no study prior to this one has tested clenbuterol-induced effects on muscle fiber type distribution and size after SCI. Results from various animal models showed that clenbuterol can prevent muscle mass loss, although it remains unclear what its actions are at the single cell level in individual fibers (Zeman et al., 1997, 1999). Also, some tests have been conducted with testosterone in SCI patients, but mainly to study spermatogenesis, muscle strength, and locomotor recovery (Clark et al., 2008; Gregory et al., 2003; Huang et al., 1999).
In contrast with data from Gregory and colleagues (2003), we found that testosterone did not prevent type I fiber conversion in hindlimb muscles (e.g., the soleus). Moreover, clenbuterol exacerbated this phase shift, since no more type I fibers were found in the soleus of clenbuterol-treated and clenbuterol + testosterone-treated mice after 2 months of treatment. In contrast, hybrid fibers were increased in proportion, suggesting a phase shift from type I towards type I + II fibers. This is supported by previous studies, showing that clenbuterol can promote a slow-to-fast fiber transition in the soleus of normal or hindlimb-unloaded rats (Criswell et al., 1996; Oishi et al., 2002; Picquet et al., 2004). On the other hand in EDL, where type I fibers do not exist in rodents, a different but comparable conversion was found: fewer hybrid fibers and more type II fibers in treated and control Tx animals.
Regarding fiber CSA changes, it is well known that muscle activity level is positively correlated with muscle mass and strength, especially in SCI patients, in whom functional electrical stimulation, passive movement, and locomotor training were shown to induce beneficial effects on muscle size and strength recovery (Forrest et al., 2008; Gerrits et al., 2001; Krause et al., 2008). However, the amount of spontaneously and progressively-occurring hindlimb movements normally found in Tx mice (Lapointe et al., 2006; Ung et al., 2007) could not explain the levels of muscle mass and CSA values found in this study, since clenbuterol-treated and clenbuterol +testosterone-treated animals displayed greater levels of muscle growth than vehicle-treated mice, whereas locomotor recovery in all groups was similar. In other words, fiber CSA increases were most probably directly induced by clenbuterol and clenbuterol + testosterone.
Some reports have provided data suggesting that slow-twitch muscles may be more sensitive to testosterone treatment than other types of muscles (Axell et al., 2006), whereas clenbuterol would primarily affect fast-twitch muscle groups (Ryall et al., 2002). Our results provide additional evidence supporting the idea of differential effects between testosterone and clenbuterol on slow- and fast-twitch fibers, since we found that EDL fibers were larger (CSA) in clenbuterol-treated or clenbuterol + testosterone-treated mice. However, the combination of clenbuterol + testosterone induced greater hypertrophic effects (e.g., soleus mass and fiber CSA values) than testosterone alone, suggesting that synergistic actions may exist between these compounds.
Locomotor recovery
As mentioned earlier, a phenomenon called spontaneous hindlimb movement recovery has been found to occur progressively (after 1 week post-Tx) in motor-complete paraplegic mice (Lapointe et al., 2006; Ung et al., 2007). However, only very low levels of activity were found, including non-locomotor-like and locomotor-like movements of small amplitude without weight-bearing capabilities. In this study, clenbuterol, testosterone, or both, did not enhance that level of spontaneous recovery, since all groups (treated and control Tx) displayed non-significantly different scores at 8 weeks post-Tx. This said, it is likely that further muscle activity induced by specific locomotor training or pharmacological approaches (CPG-activating drugs) would have affected locomotor recovery levels (Fong et al., 2005; Ichiyama et al., 2008; Landry et al., 2006). This is in contrast with data found with clenbuterol's effects on functional recovery in incompletely SCI rodents (Zeman et al., 1999), probably because clenbuterol-induced effects in their study was mainly produced by spinal-cord-repair-mediated functional recovery, rather than sublesional network-mediated property changes (e.g., increased CPG excitability).
Conclusions
Although testosterone propionate did not induce large effects on SCI-related secondary complications as shown in this study, this does not exclude the possibility that testosterone-related compounds such as specific anabolic steroids may induce significant effects in SCI patients. In fact, oxandrolone (Anavar), an anabolic steroid with a high therapeutic index (low androgenic side effects and high myogenic desired effects), was found to improve skin wound healing (pressure sores) and blood coagulation and homeostasis in SCI patients (Kahn et al., 2006; Spungen et al., 2001). Given the results obtained in our study, this may even suggest that greater effects may be elicited with combination treatments such as clenbuterol and anabolic steroids (e.g., Anavar), or selective androgen receptor modulators (SARMs), which are currently being developed by pharmaceutical companies such as GlaxoSmithKline and Schering-Plough.
Footnotes
Acknowledgments
This study was supported by the National Science and Engineering Research Council of Canada (NSERC). We also wish to thank Dr. Nicolas Lapointe for his contribution to some experiments and tissue collection.
Author Disclosure Statement
No competing financial interests exist.