
Abstract
Short-term low-frequency electrical stimulation (ESTIM) of proximal peripheral nerve stumps prior to end-to-end coaptation or tubular bridging of small distances has been reported to increase preferential motor reinnervation and functional motor recovery in animal models and human patients undergoing carpal tunnel release surgery. We investigated the effects of ESTIM on regeneration across rat sciatic nerve gaps, which exceed distances that allow spontaneous regeneration. Three different reconstruction approaches were combined with ESTIM in the experimental groups. Nerve gaps (13 mm) were bridged using (I) nerve autotransplantation, (II) transplantation of differentially filled silicone tubes, or (III) transplantation of tubular grafts containing fibroblast growth factor-2 overexpressing Schwann cells (SCs) for gene therapy. The regeneration outcome was followed for up to 8 weeks, and functionally as well as histomorphometrically analyzed in comparison to non-stimulated control groups. Combining ESTIM with nerve autotransplantation significantly increased the nerve fiber density in the regenerated nerve, and the grade of functional recovery as detected by electrodiagnostic recordings from the gastrocnemius muscle. The combination of ESTIM with transplantation of naïve SCs increased the regeneration of gap-bridging nerve tissue. Although macroscopic tissue regeneration was not further improved after combining ESTIM with FGF-221/23-kD gene therapy, the latter resulted in a high rate of regenerated nerves that functionally reconnected to the target muscle. Based on our results, brief ESTIM shows high potential to accelerate axonal as well as functional (motor and sensory) outcomes in the clinical setting of peripheral nerve gap reconstruction in human patients.
Introduction
I
Therapeutic strategies are needed that expedite regenerating axons across surgical repair sites and the placement of potent, regeneration-promoting, SCs into lengthy nerve gaps via tissue engineering. Over the last decades it has been well established that brief electrical stimulation (ESTIM) of the proximal nerve stump prior to nerve reconstruction accelerates axonal outgrowth across the lesion site and supports functional motor recovery in animal models as well as human patients (Ahlborn et al., 2007; Al-Majed et al., 2000; Brushart et al., 2002; Gordon et al., 2010, 2009). So far, most investigations were done in animal models using end-to-end nerve suture or small gap repair. There are, however, two studies describing an acceleration of axonal and functional recovery by applying ESTIM together with reconstruction of 10-mm rat femoral nerve gaps with autotransplantation (Huang et al., 2009), and 15-mm rat sciatic nerve gaps with longitudinally-oriented microchannels (Huang et al., 2010). Our own previous work in a rat model of peripheral nerve regeneration (15-mm gap in adult sciatic nerve) demonstrated that transplantation of genetically-modified SCs overexpressing fibroblast growth factor-2 (FGF-2) within silicone tubes results in a high rate of regenerated gap-bridging tissue, as well as long-distance myelination of regenerated axons. This was especially seen after transplantation of SCs overexpressing high-molecular-weight FGF-2 (FGF-221/23-kD). However, recovery of motor function was less successful than expected (Haastert et al., 2006, 2008).
The aim of the current study was to add to our knowledge about the effects of ESTIM on regeneration outcomes after reconstruction of long peripheral nerve gaps that have no probability of spontaneous regeneration (>12 mm in the rat; Lundborg et al., 1982). Therefore ESTIM was combined with reconstruction of 13-mm rat sciatic nerve gaps by autotransplantation or biosynthetic nerve guidance channels. The study was subdivided into three parts: (I) nerve autotransplantation or nerve bridging by differentially filled silicone tubes, which were (II) cell-free, or seeded with naïve or (III) genetically modified SCs for FGF-221/23-kD ex vivo gene therapy. Our results demonstrate that brief electrical stimulation of the proximal nerve stump prior to long nerve gap reconstruction sufficiently triggers regeneration processes and supports long-distance regeneration and functional reconnection of motor axons.
Methods
Experimental design
The study design is summarized in Table 1. Three sets of experiments were performed in which we reconstructed 13-mm sciatic nerve gaps in three different ways. Under control conditions only bridging of the nerve gap was performed, whereas under experimental conditions brief electrical stimulation (ESTIM: 1 h, 20 Hz, 0.3 mA) preceded surgical nerve repair. In study I, the gaps were reconstructed by nerve autotransplants. In study II and study III, we bridged the gap with silicone tubes which were filled with growth factor reduced Matrigel™ (GFR-Matrigel; Becton Dickinson, Heidelberg, Germany) as implantation matrix. In study II, the effects of ESTIM on regeneration after cell-free reconstruction of the gaps were compared with those after applying naïve neonatal rat SCs into the tubes. In study III, silicone tubes were enriched with genetically-modified SCs to enable ex vivo gene therapy with high-molecular-weight fibroblast growth factor-2 isoforms (FGF-221/23-kD).
Silicone tubes filled with growth factor reduced Matrigel™ are referred to as synthetic nerve grafts. Depending on the experimental conditions the grafts were additionally seeded with Schwann cells of different genotypes.
SCs, Schwann cells; SC-pCIneo, control transfected Schwann cells; SC-FGF-221/23-kD, Schwann cells overexpressing FGF-221/23-kD.
Animals and surgical procedures
Animal experiments were conducted in accordance with the German law on the protection of animals (approval by the animal care committee of Lower-Saxony: 33H-42502-07/1314).
Adult female Sprague-Dawley rats (200 g, 8 weeks old; Charles River, Cologne, Germany) were housed in groups of 4 rats each in cages under standard conditions (room temperature 22±2°C, humidity 55±5%, light:dark-cycle 14-h:10-h) with food and water ad libitum.
Surgical procedures were performed on consecutive days for study I, study II, and study III, and on each day the animals of all experimental groups of each study underwent surgery.
The animals were anesthetized by intraperitoneal injection of chloral hydrate (370 mg/kg body weight). To achieve sufficient analgesia, buprenorphine (0.045 mg/kg body weight) was intramuscularly applied. Body temperature was monitored and the animals were kept on an electric heating pad during anesthesia. The left hind legs were shaved, the skin was disinfected, and aseptic techniques were used to ensure sterility. The left sciatic nerve was exposed by a skin incision along the femur followed by blunt separation of the biceps femoris and superficial gluteal muscles. The nerve was freed from surrounding connective tissue.
For electrical stimulation of the proximal nerve stump prior to surgical reconstruction of the nerve, three needle electrodes (0.3 mm diameter) were inserted: one in the neck as a ground electrode, one in the belly, and one in the tendon of the left gastrocnemius muscle. The wound and nerve were kept dry with pieces of absorbent gelatin sponge, and electrically shielded using latex patches. Proximal to its trifurcation into the tibial, sural, and common peroneal nerve, the sciatic nerve was placed in a U-shaped plastic device containing a bipolar steel electrode (manufactured at Central Research Workshops, Hannover Medical School, Hannover, Germany) as shown in Figure 1A. The software-controlled electrical stimulation and electromyogram recording device (Keypoint® Portable; Medtronic Functional Diagnostics A/S, Skovlunde, Denmark) was connected to a notebook computer. The threshold current intensity (0.1 mA in all animals) was determined by stimulating the intact nerve and recording the compound muscle action potentials (CMAPs) from the gastrocnemius muscle. Afterwards the sciatic nerve was transected proximal to its trifurcation at the distal edge of the plastic tool. The proximal nerve stump was then stimulated for 1 h at a frequency of 20 Hz and a threefold threshold current intensity (0.3 mA) as previously reported (Ahlborn et al., 2007). During stimulation the wound was covered with Braunol®-soaked gauze (B. Braun Melsungen AG, Melsungen, Germany) to avoid drying of the nerve tissue and contamination of the wound.
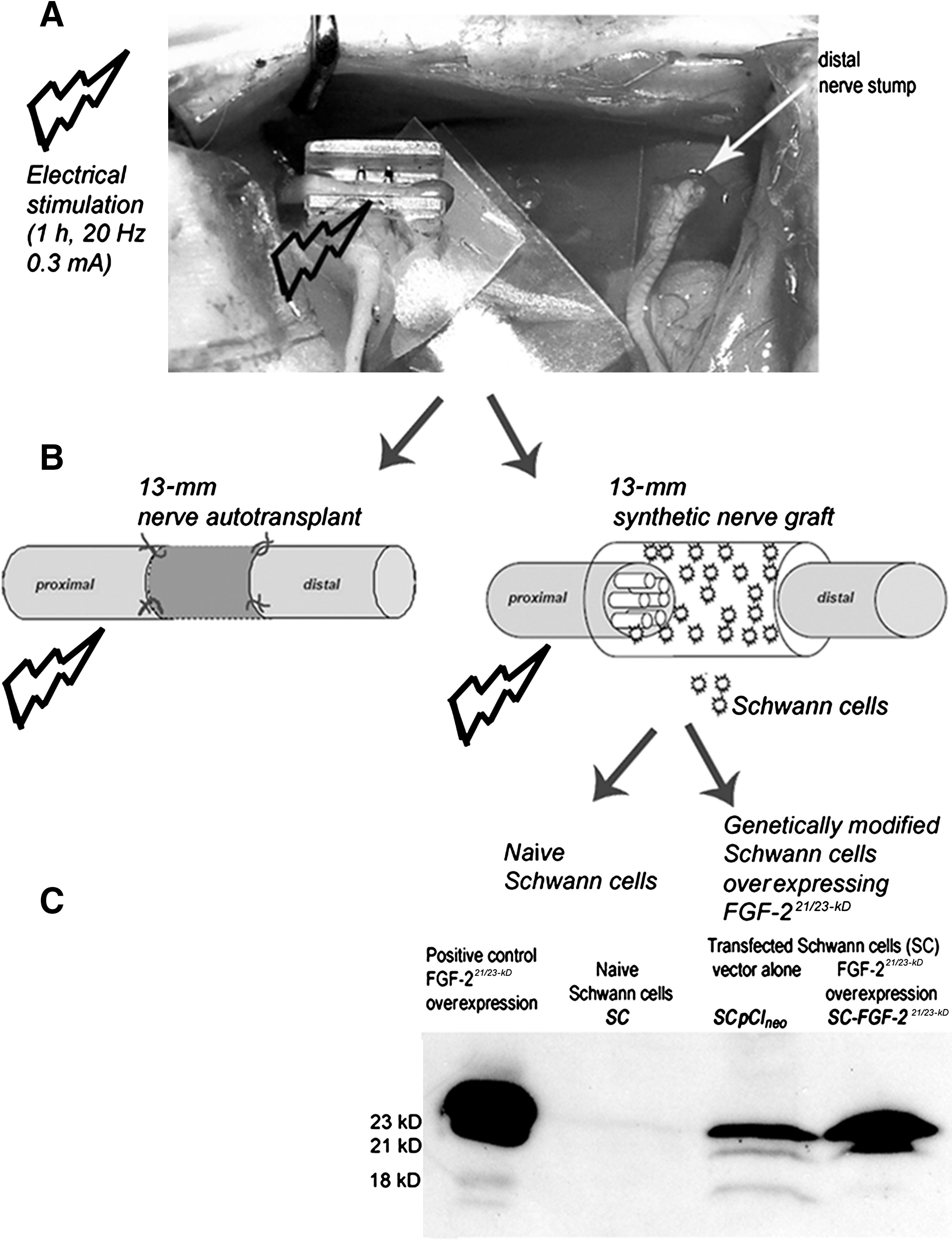
Overview of the approaches that were combined to accomplish brief electrical stimulation in studies I–III. (
For nerve reconstruction using nerve autotransplantation (study I; Fig. 1B), the nerve was transected 13 mm distal to the first transection site, and then reconstructed with epineurial sutures (9/0 Ethilon II; Ethicon, Norderstedt, Germany) 13 mm apart.
For nerve reconstruction using synthetic nerve grafts (study II and study III; Fig. 1B), a 2-mm piece from the proximal stump and a 4-mm piece from the distal nerve stump were removed (6 mm total). Afterward, differentially-filled silicone tubes 16 mm in length were transplanted between the stumps to bridge a 13-mm gap (study II and study III). Then epineurial sutures pulled the separated nerve stumps 2 mm into the proximal and 1 mm into the distal opening of the tube (Haastert-Talini et al., 2010).
Finally, the muscle layers and skin were sutured. The animals were frequently checked for automutilation and anti-bite spray was applied to the paws or a rat collar was used for 1–2 days if necessary.
Primary neonatal rat Schwann cells for transplantation
For transplantation, SCs were isolated from the sciatic nerves of 1- to 3-day-old Sprague-Dawley rat pups as previously described (Haastert et al., 2006). Briefly, primary SCs were seeded on poly-L-lysine-coated cell culture flasks (final concentration 0.5 ng/mL; Sigma-Aldrich Chemie, Munich, Germany). Twenty-four hours later, 1 mM arabinoside-C (Sigma-Aldrich) was added for 48 h to prevent growth of fibroblasts. Any remaining fibroblasts were removed using immunopanning by incubation with magnetic beads (Dynabeads; Dynal, Soro, Denmark) covered with α-Thy1-antibody (Haastert et al., 2005). Pure SC cultures (>95%) were harvested after the ninth or tenth passage, and removed from serum by consecutive medium exchanges every 24 h with medium containing 10%, 5%, and finally 0% fetal calf serum. The serum-free medium was substituted with N1-supplement (apo-transferrin, putrescine, progesterone, and sodium selenite), and naïve SCs were ready for transplantation 24 h later (study II and study III).
Genetically modified SCs for FGF-221/23-kD ex vivo gene therapy
Overexpression of FGF-221/23-kD was induced by SC transfection using Metafectene™ (Biontex, Martinsried, Germany) and plasmid constructs with FGF-221/23-kD in pCIneo. Transfected SCs were positively selected for their co-expressed resistance to geneticin (G418). As shown in Figure 1C, successful transfection and FGF-221/23-kD overexpression was monitored with Western blot analysis (Haastert et al., 2008). Control transfected SCs were exposed to pCIneo vectors solely carrying the geniticin resistance.
Preparation of silicone tubes for transplantation
GFR-Matrigel aliquots and cell pellets were freshly prepared for each animal and kept on ice until silicone tubes (1.5-mm diameter and 0.4-mm wall thickness) were filled for transplantation as indicated in Table 1. Cell-free silicone tubes contained 25 μL GFR-Matrigel, and for SC transplantation the cells were suspended in the matrix to a final density of 114,286 SCs/μL GFR-Matrigel. Each synthetic graft contained approximately 2×106 SCs.
Quantitative RT-PCR
To estimate the effect of ESTIM on the stimulated neuron pools in combination with silicone tube transplantation, 16 additional animals were subjected to analysis of mRNA regulation on the dorsal root ganglion (DRG) and lumbar spinal cord levels. In one-half of the animals proximal nerve stumps were left under unstimulated control conditions, while the other half of the animals received ESTIM. Afterward, 13-mm nerve gaps were reconstructed by cell-free silicone tubes. Two days and 2 weeks after surgery, respectively, DRGs (L4–;L6) and spinal cord sections (L3–L6) from the ipsilateral and contralateral sides were dissected and snap frozen in liquid nitrogen (n=4–5 animals each for the control and ESTIM groups). Individual tissue samples were homogenized in Trizol reagent, and total RNA was extracted per the manufacturer's instructions. To eliminate any genomic DNA contamination a DNase digest (Stratagene; Agilent Technologies Sales & Services, Waldbronn, Germany) was performed. Total RNA (1 μg) was converted into cDNA using random primers and M-MLV reverse transcriptase. The primer sequences were as follows: brain-derived neurotrophic factor (BDNF)-F: 5′-GGACATATCCATGACCAGAAAGAA-3′; BDNF-R: 5′-GCAACAAACCACAACATTATCGAG-3′; growth-associated protein-43 (GAP-43)-F: 5′-GATGGTGTCAAACCGGAGGAT-3′; GAP-43-R: 5′-CTTGTTATGTGTCCACGGAAGC-3′; and glyceraldehyde-3-phosphate dehydrogenase (GAPDH)-F: 5′-GAACATCATCCCTGCATCCA-3′; GAPDH-R: 5′-CCAGTGAGCTTCCCGTTCA-3′. Equal PCR efficiency of all primer pairs was validated by serial cDNA dilutions and estimated to be 100%. Quantitative RT-PCR was performed in 96-well plates using the StepOnePlus instrument (Applied Biosystems, Darmstadt, Germany), running with the cycling program of 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec, and 60°C for 1 min. Finally a dissociation curve was calculated for each well to ensure specificity of the PCR product. The 14-μL reaction mix contained 5 μL cDNA (corresponding to 1.25 ng RNA), 3 μL primer mix (5.25 pmol of the F and R primers), and 7 μL Power SYBR-Green PCR Master Mix (Applied Biosystems). The samples were run in duplicate. Fold changes in mRNA levels compared to the levels measured 2 days after surgery on the contralateral side of control animals were calculated using the 2(−ΔΔCt)-1 method (Applied Biosystems User Bulletin #2, 2001; Jungnickel et al., 2010), and normalized to the housekeeping gene GAPDH.
Behavioral tests
After nerve autotransplantation (study I, Table 1) functional recovery was evaluated weekly over a period of 8 weeks after surgery. Only animals with no signs of automutilation were evaluated, and those with slightly reddened toe tips due to nail biting were excluded from behavioral tests for one or two tests. The changes in numbers of evaluated animals per group are indicated in the results. Motor recovery was assessed by walking track analysis, and sensory recovery was evaluated using the pinch test. The observers were blinded to study group. After surgery all animals were numbered consecutively by the surgeon (K.H.T.), and the animal codes were revealed after all measurements were performed and analyzed for each animal.
Walking track analysis: Sciatic Function Index (SFI)
The rats were allowed to walk up a small inclined gangway (slope 20°, length 1 m, width 14 cm) lined with white paper after their hind paws were carefully pressed onto inkpads. The footprints were digitized and the print length (PL; distance from the heel to the third toe), the toe spread (TS; the distance between the first and fifth toes), and the intermediate toe spread (ITS; the distance between the second and fourth toes) were calculated using the Footprint program (Haastert-Talini et al., 2010; Klapdor et al., 1997). Measurements were taken for the left experimental paw (EPL, ETS, and EITS), as well as the right non-operated paw (NPL, NTS, and NITS). The SFI was calculated using the following formula (Bain et al., 1989; Haastert-Talini et al., 2010):
In healthy animals the SFI is about 0, and it decreases to −100 after complete impairment of the sciatic nerve.
Pinch test of mechanical sensitivity
In cases of recovery of mechanical sensitivity, pinching the most distal portion of the three lateral digits on the lesioned and on the third toe of the unlesioned control hindpaw with flattened forceps results in foot withdrawal and vocalization (Haastert-Talini et al., 2010). To ensure consistent pressure with the forceps for all trials, all tests were performed by the same person.
Macroscopic tissue regeneration and electrodiagnostic measurements
At the end of the observation period, all animals were anaesthetized and the silicone tubes were inspected for gap-bridging tissues between the nerve stumps (studies II and III, Table 1).
Afterward, functional reinnervation of the gastrocnemius muscle was analyzed prior to tissue explantation. The ipsilateral and contralateral sciatic nerves were exposed and electrically shielded from the surrounding tissue using latex patches. A bipolar hook steel electrode was placed on the nerve proximal and distal to the suture site. Single rectangular stimuli of 0.1-msec duration were pulsed by a software-controlled stimulus generator. The stimulus intensity was gradually raised from the threshold of a minimum response to a level 30% above the maximum response (not above 8 mA). To determine threshold current intensities, current intensity was raised from 0 mA in gradual steps of 0.1 mA (0–2 mA), or 0.2 mA (2 mA and higher), and the CMAPs were recorded from the gastrocnemius muscle (with bipolar electromyography needle electrodes inserted into the tendon and belly), and displayed on the screen of a notebook computer connected to the Keypoint Portable EMG-System. CMAPs or evoked hindpaw movements were considered signs of successful reinnervation. The term “evoked hindpaw movements” describes spreading of the lateral toes of a hindpaw upon direct nerve stimulation, and is illustrated in Figure 2A. Furthermore, the motor nerve conduction velocity (MNCV) was calculated. The MNCV of the right unoperated sciatic nerve served as an internal reference.
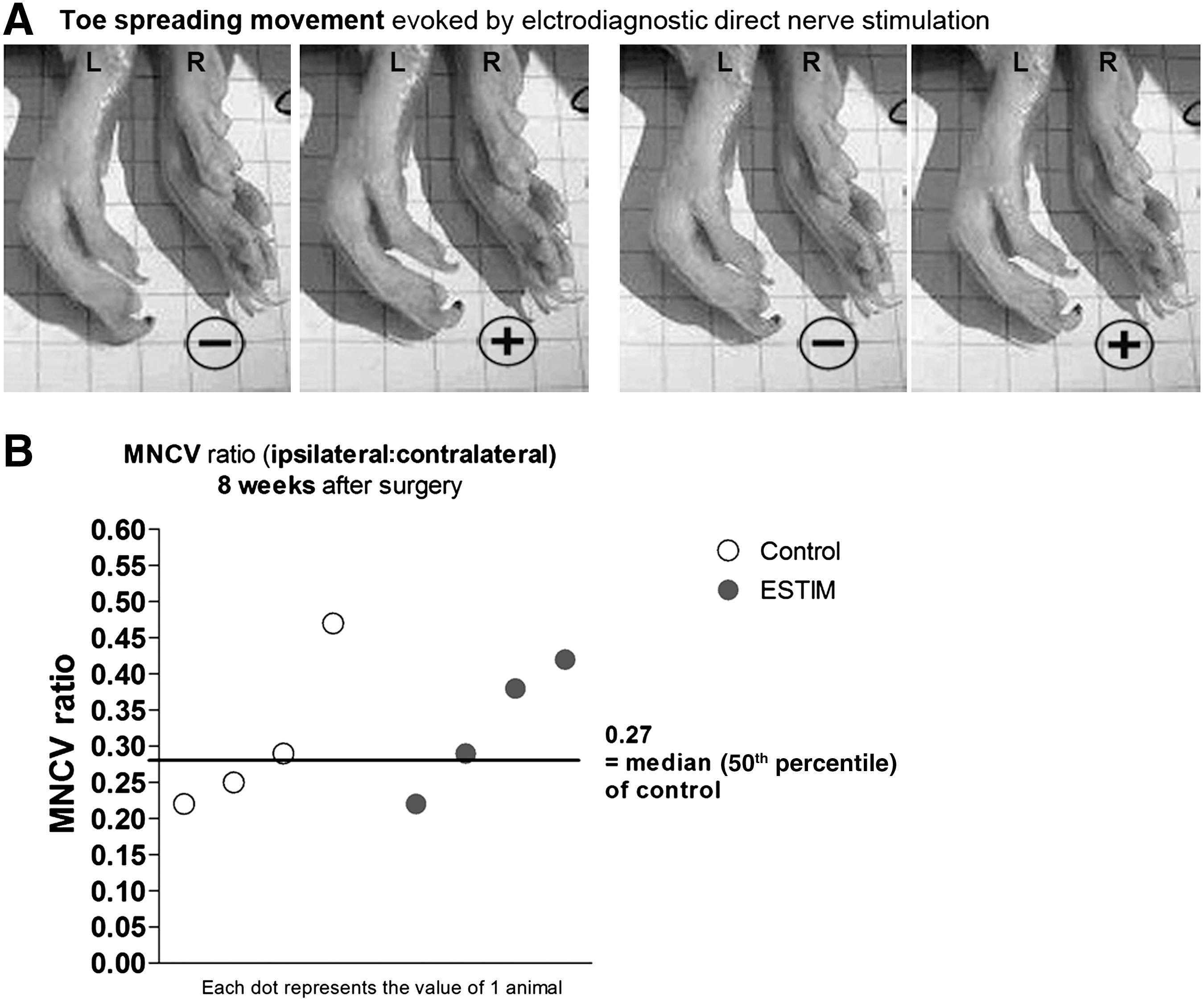
(
Nerve morphometry
Upon explantation, the regenerated nerve tissue was transected at mid-graft level, 6.5 mm distal to the original proximal nerve stump. The sutures connecting the proximal and distal nerve stumps to the autotransplant (study I), or nerve tube (study II and study III), were still visible to identify the original nerve stumps. In addition a 5-mm segment of the distal nerve was harvested 1 mm distal to the autotransplant or nerve tube. The regenerated nerve tissue was transferred to a fixative according to Karnovsky (2% PFA and 2.5% glutaraldehyde in 0.2 M sodium cacodylate buffer, pH 7.3; Haastert et al., 2006; Haastert-Talini et al., 2010) for 24 h, and then rinsed three times with 0.1 M sodium cacodylate buffer containing 7.5% sucrose, prior to post-fixation in 1% OsO4 for 1.5 h. Myelin staining was performed as previously described (Haastert et al., 2006; Haastert-Talini et al., 2010), for 24 h in 1% potassium dichromate, followed by 24 h in 25% ethanol, and for another 24 h in hematoxylin (0.5% in 70% ethanol). After dehydration, the tissue was epon-embedded and thin (1-μm) cross-sections were cut with glass knives (Ultramikrotome System, 2128 Ultrotome®; LKB, Bromma, Sweden), mounted on uncoated glass slides, and stained with toluidine blue to further enhance the myelin staining (Haastert et al., 2006; Haastert-Talini et al., 2010).
Regenerated myelinated axons (mAx) were analyzed as indicated in the results section. The cross-sections were digitized using an Olympus BX60 microscope at 400×magnification and 1288×966 dpi, and the images were aligned into total cross-sections using Cell^P® software, and were analyzed using AnalySISPro® 3.1/3.2 software (Soft Imaging System GmbH, Munster, Germany; Haastert et al., 2010; Haastert-Talini et al., 2010).
Because of the large size of the cross-sections after nerve autotransplantation (study I), the number of mAx was determined only in defined parts of the cross-sections (Geuna et al., 2004), followed by measuring the cross-sectional area and extrapolation to estimate the total number of mAx. The number of mAx was set in relation to the entire cross-sectional area to calculate the nerve fiber density (mAx/mm2). The g ratio, an index of the grade of axon myelination determined by the axon diameter divided by the total fiber diameter, was evaluated for at least 200 mAx for each section (Haastert et al., 2010), by an indirect calculation using the following formula: g ratio=axonal diameter of myelinated fibers/(axonal diameter of myelinated fibers +2×myelin thickness).
After regeneration through a synthetic nerve graft (study II and study III), the entire cross-sectional area was analyzed to determine the total number of regenerated mAx and the nerve fiber density (no. of mAx/cross sectional area [mm2]). In experimental groups with a minimum of three regenerated nerves, myelin thickness (μm) and g ratio were determined for all mAx within 2–5 grid pattern fields (100×100 μm) for up to 200 axons where possible (modified from Geuna and associates, 2001). All quantification was done by an observer blinded to experimental conditions. Blinding was achieved by consecutive numbering of each nerve sample. The code numbers were revealed after histomorphometric analysis was completed for all samples.
Statistical analysis
For statistical analysis GraphPad InStat software, version 3.06 for Windows (GraphPad Software, San Diego, CA) was used. The non-parametric Mann-Whitney U test was chosen because only a few data passed the normality test. In study I and study III, for which animal numbers of 7–10 per group were analyzed, the overall effect of SC transplantation or ESTIM on regeneration parameters was analyzed using the chi-square test. A p value of <0.05 was taken as statistically significant. The statistical tests used are indicated in the results section.
Results
Results of study I: 13-mm nerve autotransplantation
In our first attempt, we investigated the effect of brief electrical stimulation of the proximal sciatic nerve stump (ESTIM) on peripheral nerve regeneration in combination with reconstruction of 13-mm sciatic nerve gaps by nerve autotransplantation. As indicated in Table 1, the observation periods were 2, 4, and 8 weeks for 3–4 animals under unstimulated control conditions, and 4 animals after ESTIM.
Table 2 summarizes the differences that appeared between the control and ESTIM groups during the evaluation of functional regeneration parameters. Analysis of motor recovery by calculation of the sciatic function index (SFI) did not reveal any differences between both during the first 5 weeks after surgery. No significant recovery of the SFI was seen (SFI values remained at the fully-impaired level of −100). At 6 and 7 weeks 1 animal from each group had to be excluded from the measurement due to signs of toe biting, and 1 week later 1 more control animal had also been excluded from the measurement. However, 3 animals per group were available for footprint analyses at 6 and 7 weeks after surgery, and although not significantly different due to the small sample size, the mean±standard error of the mean (SEM) of the best SFI values of the animals that underwent ESTIM were higher than the mean of the control group (Table 2). Eight weeks after surgery the values had not increased further, and only 2 control animals remained available for analysis.
Over 8 weeks of evaluation, recovery of mechanical sensitivity as tested by the pinch test was never seen in the most lateral fourth and fifth digits. A positive pinch reflex of the third toe was detectable significantly earlier after ESTIM than after nerve autotransplantation alone (Table 2).
Electrical stimulation of the reconstructed sciatic nerve 4 weeks after surgery resulted in visible muscle contraction of the gastrocnemius muscle in all animals analyzed at this time point. However, functional reinnervation of the hindpaw muscles differed significantly, as indicated by the fact that more animals showed spreading of their lateral toes evoked by direct nerve stimulation in the ESTIM group. Figure 2A shows a representative sequence of photographs taken of toe-spreading movements upon direct electrical stimulation of the regenerated left sciatic nerve 4 weeks after ESTIM. The mass of the gastrocnemius muscle was still too small to allow recording of CMAPs 4 weeks after surgery, but 8 weeks after surgery this and the calculation of the MNCV were possible. After direct electrical stimulation of the nerve, latencies and maximal amplitudes still differed significantly (two-tailed p<0.02 by the Mann-Whitney U test) between healthy control nerves (latency: 1.86±0.07 msec, max amplitude: 50.07±2.39 mV), and those with autotransplantation (latency: 2.9±0.36 msec, max amplitude: 15.50±0.94 mV), and those with ESTIM+autotransplantation (latency: 2.65±0.06 msec, max amplitude: 14.86±2.66 mV). Regarding latencies and amplitudes, no differences could be detected after nerve autotransplantation or its combination with ESTIM. However, direct stimulation of the nerve distal to the autotransplant revealed some differences. Threshold current intensities needed to elicit CMAP signals within the gastrocnemius muscles, and current intensities that were needed to elicit maximal amplitude CMAPs were lower in animals that underwent ESTIM than in control animals (Table 2). Figure 2B presents a scatterplot of the values of the MNCV ratios (ipsilateral MNCV/contralateral MNCV) of control and ESTIM animals. The median MNCV ratio (0.27) of the control animals is helpful in elucidating the difference in motor recovery between the groups.
Results from the morphometric analysis of semi-thin nerve cross-sections after autotransplantation are depicted in Figure 3A–F. Clear differences were seen in nerve fiber density values between the control and the ESTIM groups. Furthermore, significant differences were detected by the Mann-Whitney U test for comparison of g ratio mean values. Two weeks after surgery, the nerve fiber density (mAx/mm2) at the midpoint of the autotransplant was visibly increased after ESTIM (Fig. 3A). At 4 and 8 weeks after surgery the endpoint of the autotransplant was analyzed (Fig. 3C–F), and similarly demonstrated higher nerve fiber densities when ESTIM was combined with nerve autotransplantation (Fig. 3C and E). As depicted in Figure 3B, D, and F, at all time points analyzed the g ratio of the axons was lower after ESTIM (2 weeks: 0.73±0.02, 4 weeks: 0.72±0.01, and 8 weeks: 0.72±0.02) than in the control group (2 weeks: 0.83±0.01, 4 weeks: 0.81±0.02, and 8 weeks: 0.82±0.01). This difference became statistically significant at 8 weeks after surgery (Fig. 3F).
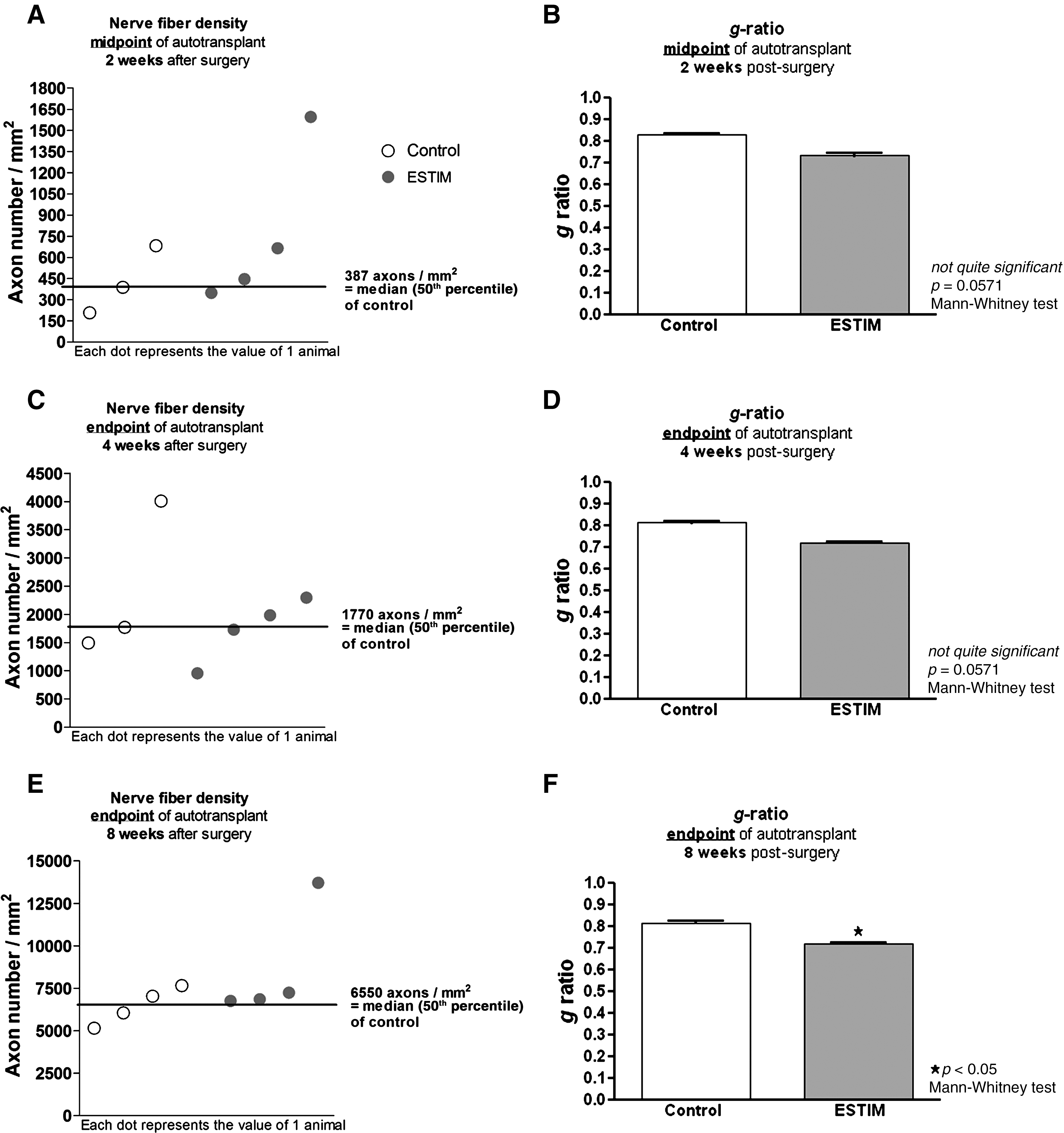
Graphical overview of the animal nerve fiber densities shown as scatterplots (
Results of study II: Synthetic nerve graft bridging 13-mm nerve gaps
In study II we investigated the ESTIM effect on peripheral nerve regeneration in combination with the use of cell-free or SC-seeded silicone tubes to bridge a 13-mm sciatic nerve gap. Observation periods were 4 and 8 weeks (Table 1).
As depicted in Figure 4A, only few animals in the cell-free groups had silicone tubes containing regenerated gap-bridging tissue at the 4-week time point. While macroscopic tissue regeneration was only seen in 11% of unstimulated nerves, this rate was increased to 22% by ESTIM. Transplantation of naïve SCs significantly increased the rate of macroscopic tissue regeneration of unstimulated nerves. A combination of SC transplantation with ESTIM further supported macroscopic tissue regeneration.
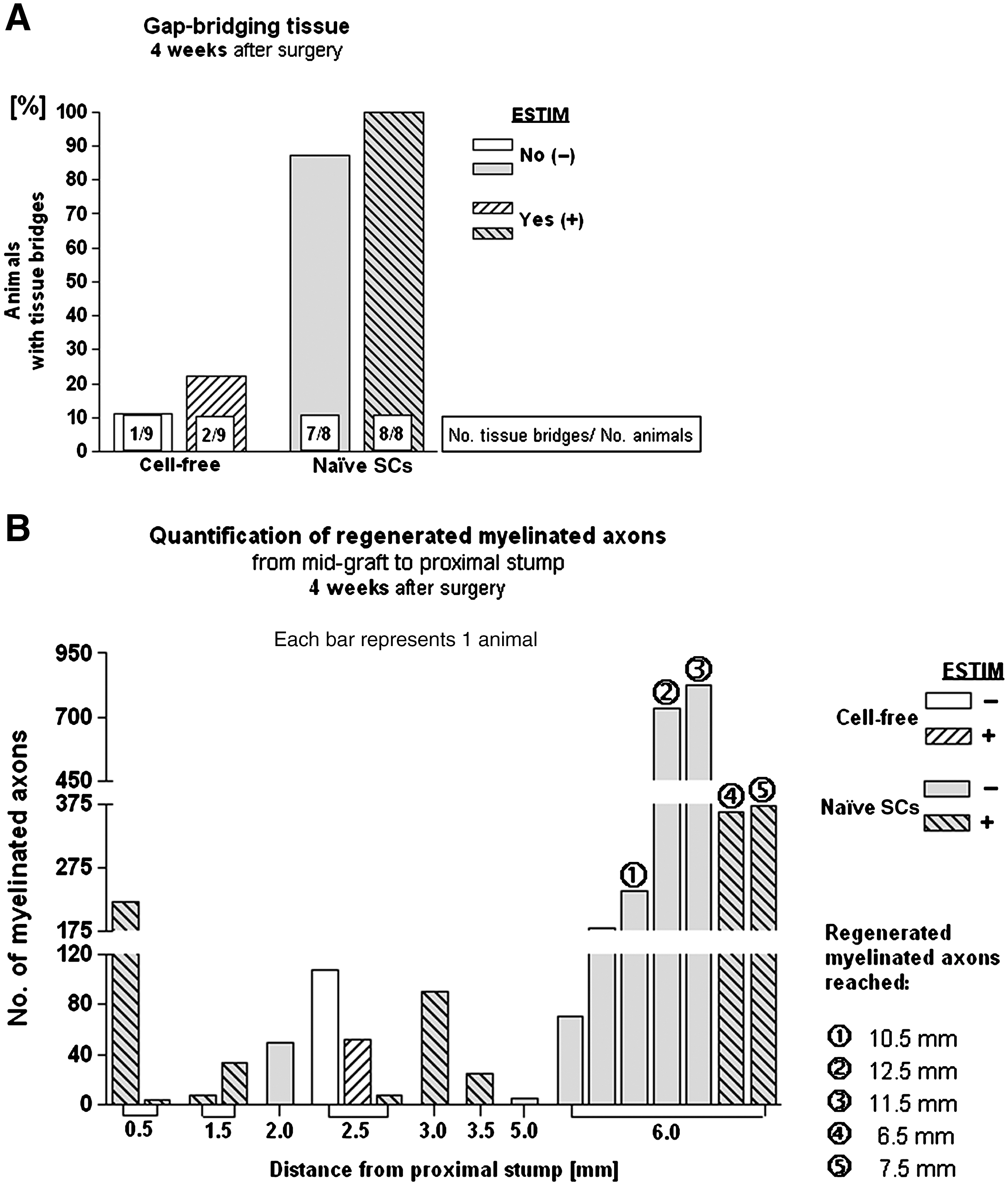
Results of study II at the 4-week time point. (
However, electrical stimulation of the regenerated nerve proximal or distal to the synthetic graft did not result in evocable hindpaw movement in any of the animals at 4 weeks after surgery.
Semi-thin cross-sections through the regenerated tissue were morphometrically evaluated for the distances across which mAx regenerated at 4 weeks after surgery. Thus starting at mid-graft level (6.5 mm from the proximal nerve stump), sections were analyzed every 0.5 mm in both the proximal and distal directions. Figure 4B shows that mAx did not cross the midline of the graft in most samples. The regeneration distance of mAx was significantly enhanced after SC transplantation (p<0.001 by the chi-square test). Comparison of axonal regeneration under control conditions or after ESTIM revealed in control group samples significantly more and longer mAx (p<0.001 by the chi-square test). No differences in the g ratios of regenerated mAx could be detected (0.68–0.7 in all animals analyzed).
Figure 5 summarizes the results seen 8 weeks after surgery. With regard to macroscopic tissue regeneration, the outcome nearly reflected the results seen after 4 weeks (Fig. 4A), with significantly better tissue regeneration seen after SC transplantation, which was further increased by ESTIM (Fig. 5A). Morphometric analysis of the regenerated tissue at mid-graft level revealed no significant differences between control-SC and ESTIM-SC samples (Fig. 5B). Evaluation of the regeneration distance of mAx again reflected the situation seen 4 weeks after surgery, with more mAx regenerated further distally in unstimulated SC animals. However, exclusively after ESTIM, one animal showed substantial axonal regeneration through the graft, resulting in reinnervation of hindpaw muscles 8 weeks after surgery (Fig. 5C). In this animal toe-spreading movements could be evoked by electrical nerve stimulation proximal and distal to the graft prior to nerve dissection.
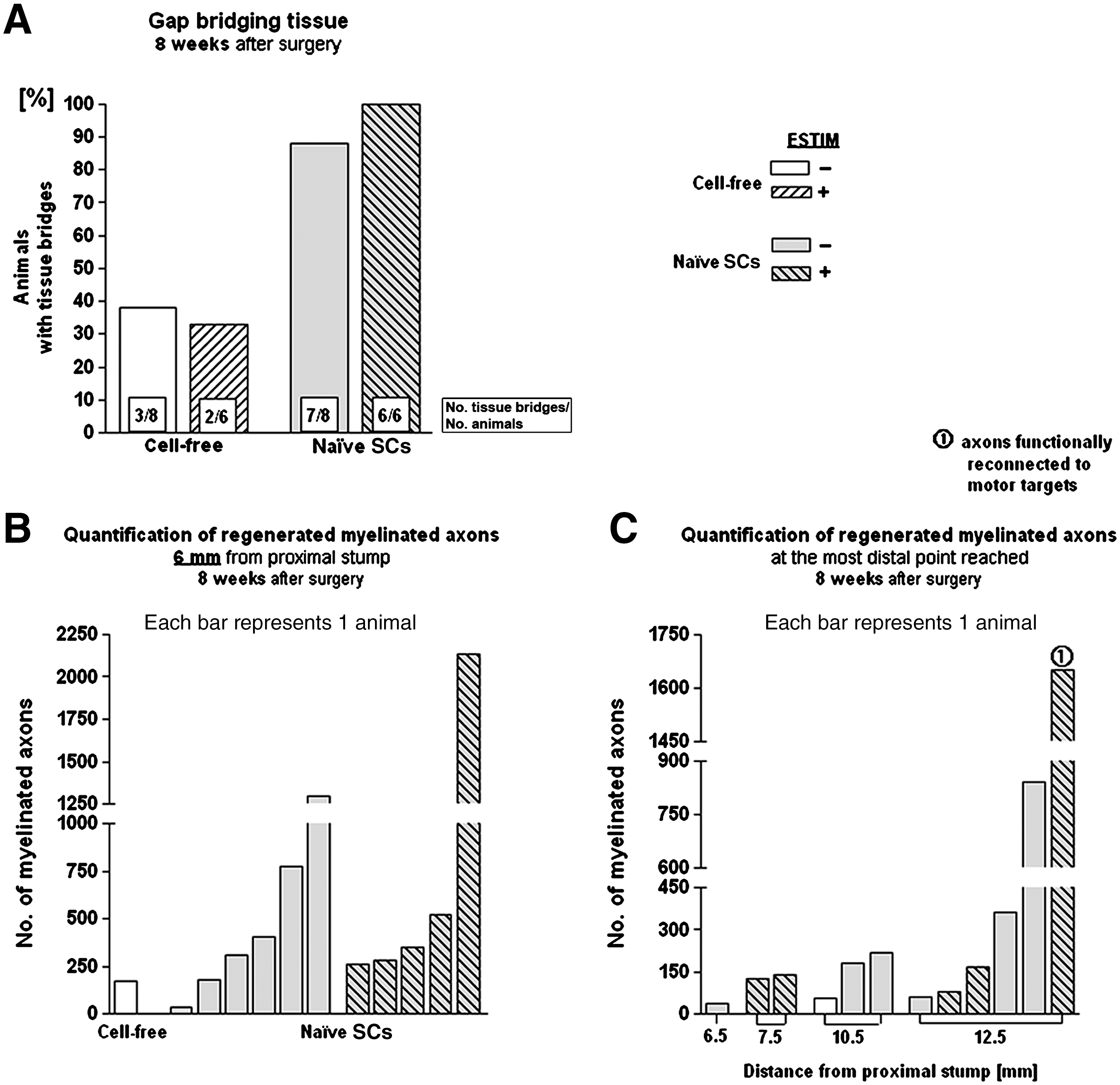
Results of study II at the 8-week time point. (
Results of study III: Synthetic nerve graft+FGF-221/23-kD gene therapy
The results of study I indicated a regeneration-promoting effect of ESTIM in the scenario of long nerve gap repair. The combination of ESTIM with bridging of 13-mm nerve gaps with SC-seeded synthetic nerve grafts in study II resulted in an outcome of regeneration that strongly indicated that the synthetic nerve grafts have to be further modified to allow ESTIM to substantially support axonal and functional regeneration. Our hypothesis was that ESTIM would lead to motor recovery after transplantation of FGF-221/23-kD overexpressing Schwann cells (SC-FGF-221/23-kD), which have previously been shown to account for long-distance regeneration of myelinated axons (Haastert et al., 2006, 2008).
As depicted in Figure 6, in comparison to SC transplantation alone, ESTIM again clearly increased the rate of macroscopic regeneration of gap-bridging tissue, while the combination of ESTIM with FGF-221/23-kD gene therapy dramatically reduced this rate (Fig. 6A). However, in the SC-FGF-221/23-kD group, 2 out of only 3 regenerated nerves (67%) demonstrated functional recovery as indicated by toe-spreading movements that could be evoked by electrical nerve stimulation proximal and distal to the graft prior to nerve dissection. The rate of functional recovery was therefore dramatically higher after a combination of ESTIM with FGF-221/23-kD gene therapy as in the other groups investigated (Fig. 6B). Figure 6C depicts the results from quantification of the number of regenerated mAx, revealing that ESTIM significantly increased the regeneration distances, in combination with the transplantation of naïve SCs (p<0.001 by the chi-square test). Figure 6D shows representative details of semi-thin cross-sections through the regenerated nerve tissue after a combination of ESTIM with (I) nerve autotransplantation, (II) transplantation of SCs, or (III) transplantation of SC-FGF-221/23-kD. Exclusively after ESTIM functional muscle reinnervation occurred 8 weeks after surgery (Fig. 6C), an effect that was seen in study II as well (Fig. 5C).
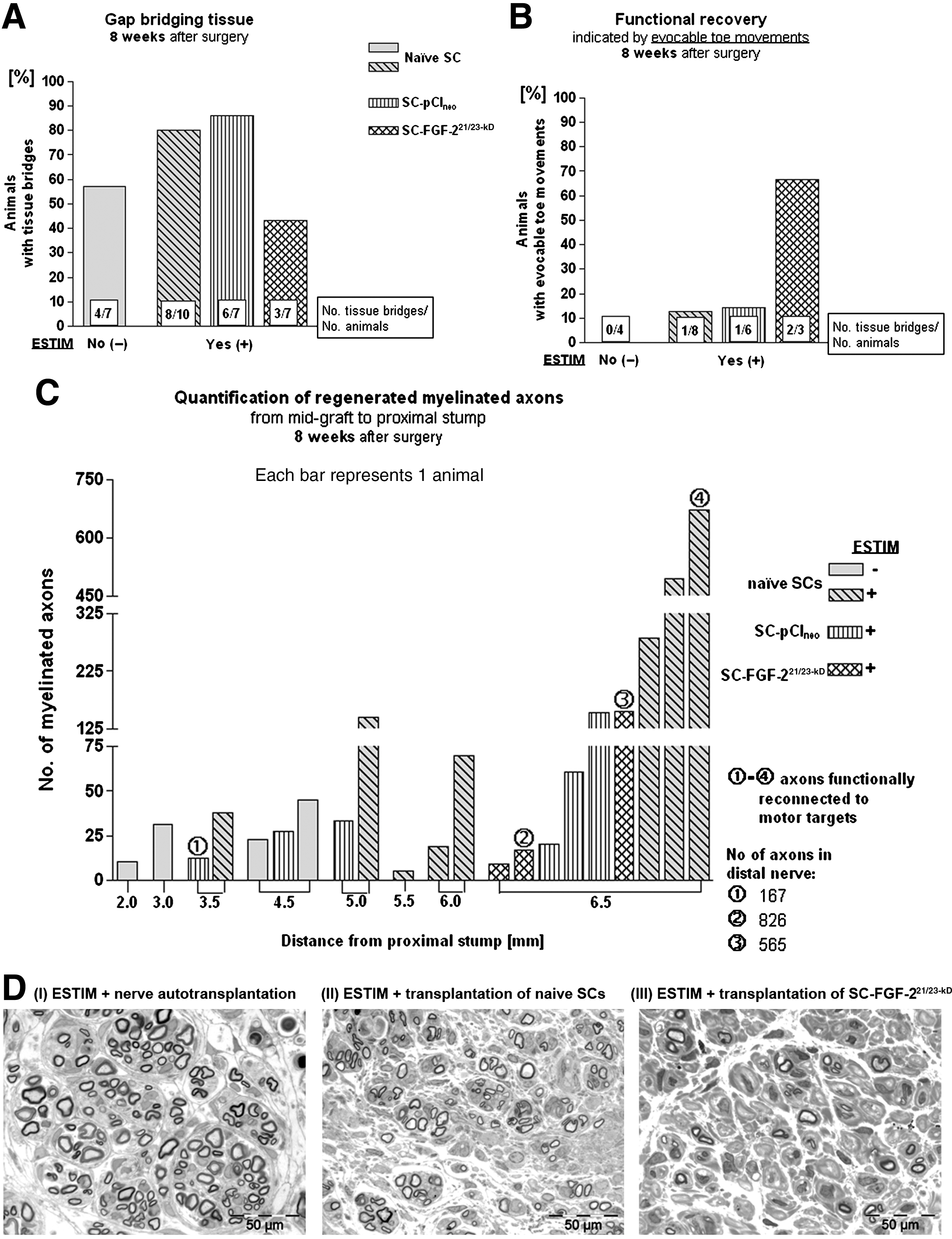
Results of study III at the 8-week time point. (
Results of quantitative RT-PCR
In addition to the effect on actual regeneration outcome, we further analyzed the molecular consequences of ESTIM on the associated neuron pools in the dorsal root ganglia (DRGs; L4–L6), and the lumbar spinal cord (L3–L6). Therefore we performed mRNA level analyses 2 days as well as 2 weeks after reconstruction of 13-mm nerve gaps with cell-free synthetic nerve grafts according to study II (Table 1). Ipsilateral and contralateral effects on the mRNA levels of brain-derived neurotrophic factor (BDNF) and growth-associated protein-43 (GAP-43) were subjects of investigation.
As depicted in Figure 7, ipsilateral BDNF mRNA levels were significantly upregulated in DRGs of control animals (without ESTIM) 2 days after surgery compared to contralateral levels (Fig. 7A). The post-surgical ipsilateral upregulation was not significant after ESTIM because of an additional partial contralateral upregulation (Fig. 7A). Two weeks after surgery, ipsilateral DRG BDNF mRNA levels were significantly downregulated in both groups compared to the 2-day levels, and reached reference levels measured 2 days after surgery on the contralateral side of control animals (without ESTIM, Fig. 7A).
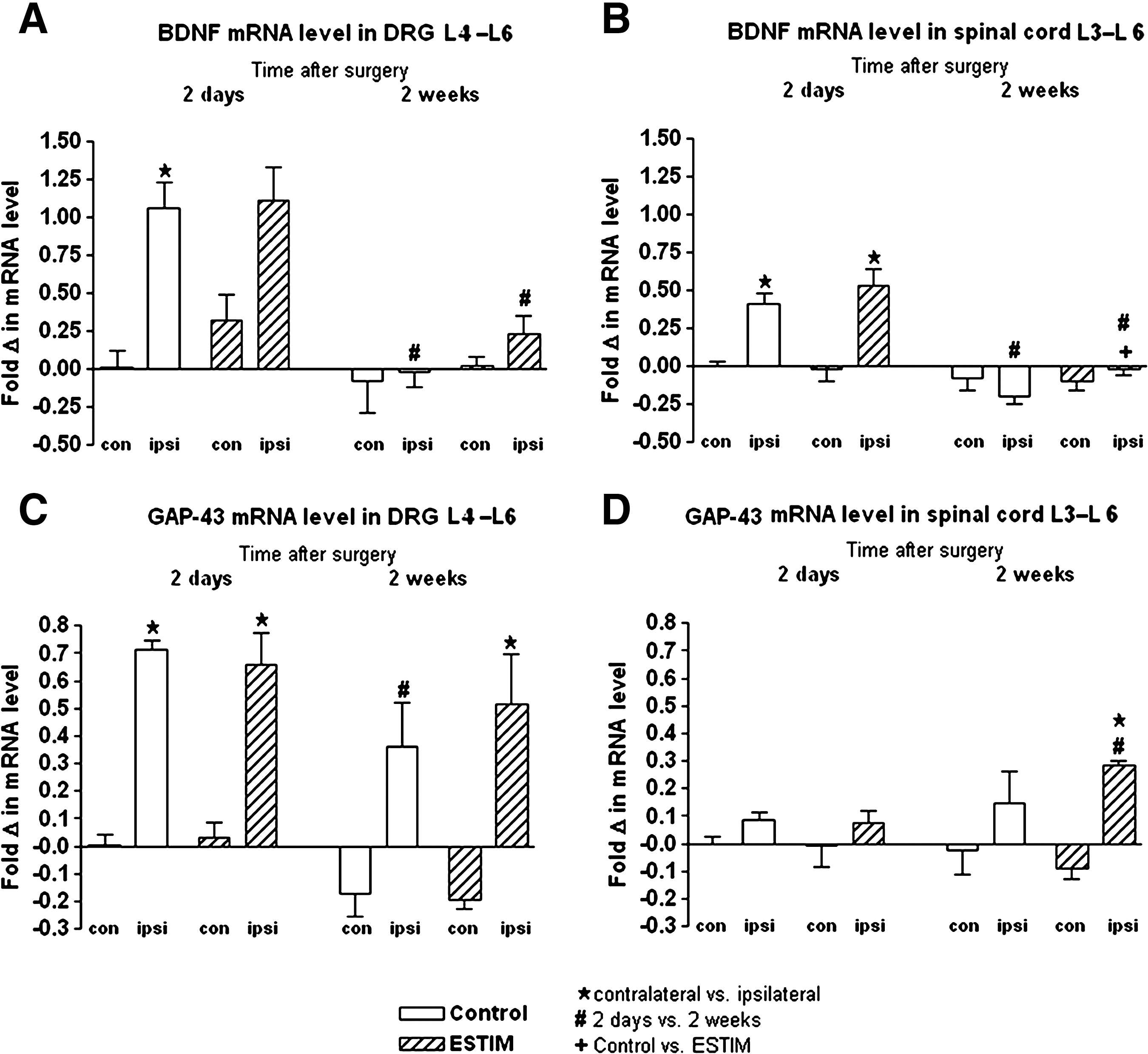
(
Within the lumbar spinal cord, ipsilateral BDNF mRNA levels were significantly upregulated 2 days after surgery in both groups investigated, and this effect was no longer seen 2 weeks after surgery (Fig. 7B). Two weeks after surgery, however, ipsilateral BDNF mRNA levels within the lumbar spinal cord were significantly lower under unstimulated control conditions as after ESTIM (Fig. 7B).
Significant upregulation of GAP-43 mRNA levels was seen 2 days after surgery in ipsilateral versus contralateral DRGs in both groups (Fig. 7C). Under control conditions, ipsilateral DRG GAP-43 mRNA levels were significantly downregulated 2 weeks after surgery compared to 2 day levels, and side differences were no longer detectable (Fig. 7C). After ESTIM, however, ipsilateral DRG GAP-43 mRNA levels remained significantly higher as on the contralateral side 2 weeks after surgery (Fig. 7C).
As indicated in Figure 7D, GAP-43 mRNA levels within the ipsilateral spinal cord were significantly upregulated exclusively 2 weeks after ESTIM (compared to contralateral 2-week levels, as well as ipsilateral 2-day levels).
Discussion
The current study combined three different approaches to reconstruct lengthy nerve defects with brief ESTIM of the proximal nerve stump, and clearly demonstrates that ESTIM is sufficient to accelerate long-distance regeneration of motor and sensory axons, resulting in an increase of functional regeneration, especially after nerve autotransplantation. The current results indicate that brief ESTIM at the time of nerve reconstruction may be a useful tool to accelerate the regeneration outcome after nerve autotransplantation in humans.
When trauma or tumor resection leads to long distances between transected peripheral nerve stumps, the mechanisms impeding timely and appropriate target reinnervation become even more obstructing, as after tension-free end-to-end suture or short-gap repair. It has long been discussed that the rate of irreversible muscle atrophy accounts for the insufficient functional recovery seen after severe peripheral nerve injuries, although elaborate microsurgical techniques are used for reconstruction (Fu and Gordon, 1997; Gordon et al., 2009). We know today that it is instead the duration of chronic motorneuron target deprivation, as well as the duration of SC deprivation from axons that determine the recovery rate (Gordon et al., 2009). The two main impeding factors for axonal regeneration are therefore slow growth across coaptation sites and/or across nerve gaps, and the relatively short time frame when the injured neurons and axons have the capacity to regenerate, and when the denervated SCs can support axonal regeneration (Gordon et al., 2009). It has been reported before that brief ESTIM combined with end-to-end nerve coaptation or short nerve gap reconstruction accelerates the rate of axonal regeneration, and shortens the time for regenerating axons to functionally reconnect to their appropriate targets (Gordon et al., 2009).
The current study was conducted with the aim to study the effects of ESTIM on the outcome of axonal and functional regeneration after reconstruction of nerve gaps that are not subject to spontaneous regeneration (13 mm in the rat; Lundborg et al., 1982). Our results clearly demonstrate that also under the more challenging conditions of long-distance regeneration after nerve autotransplantation, ESTIM positively affects axonal outgrowth, functional muscle reinnervation, and appropriate myelination of regenerated axons. Nerve fiber density, as well as myelination of regenerated axons according to their diameter (g ratio), was supported by ESTIM, which finally resulted in increased nerve functionality (MNCV ratio). Previous studies indicate that sensory nerve regeneration can also be supported by ESTIM (Brushart et al., 2005; Geremia et al., 2007; Wang et al., 2009). The subject of sensory recovery has not been investigated before in studies combining ESTIM with the reconstruction of lengthy nerve defects (Huang et al., 2010). Our results, however, further demonstrate that ESTIM accelerates recovery of mechanical sensitivity when combined with 13-mm nerve autotransplantation.
Reconstruction of the 13-mm nerve gap with biosynthetic grafts further aggravates the regeneration process, because the growth-permissive environment (Lundborg, 2004) of the autotransplant or distal nerve stump cannot be reached immediately by the outgrowing axons. In the current study, nerve tissue regeneration through the synthetic nerve grafts was enhanced after transplantation of SCs, an effect that could have been expected (Haastert et al., 2006; Lundborg, 2004). Our results further demonstrate an ESTIM-related increase in the rate of tissue regeneration, as well as the elongation of regenerating myelinated motor axons across the nerve gaps, leading to successful muscle reinnervation that could be demonstrated with electrodiagnostic measurements. In the chosen time window of 8 weeks post-surgery, functional muscle reinnervation could exclusively be achieved in stimulated animals. The overall axonal regeneration outcome after reconstruction of the 13-mm nerve gap with differentially filled biosynthetic grafts was, however, much less successful after nerve autotransplantation, which indicates the well-identified need for elaborated innovative biomimetic nerve grafts (Battiston et al., 2009; Chiono et al., 2009). A recent study did combine ESTIM with reconstruction of 15-mm rat sciatic nerve gaps using longitudinally-oriented microchannel grafts, and demonstrated axonal and motor regeneration rates like those seen after nerve autotransplantation without ESTIM (Huang et al., 2010). Following our results it still has to be postulated, however, that nerve autotransplantation is the undisputed gold standard for reconstruction of lengthy peripheral nerve defects, and it will not be replaced by biomimetic nerve transplants in the near future, but it may be amplified in its regeneration-promoting effect by combining it with ESTIM.
Regarding the mechanism behind the beneficial effects of ESTIM, it is well established that ESTIM in combination with end-to-end nerve suture upregulates mRNA or protein expression of BDNF, as well as of its tyrosine kinase B (trkB) high-affinity receptor in single motor and sensory neurons (Al-Majed et al., 2000; English et al., 2007). An upregulation of GAP-43 mRNA related to ESTIM was also reported for motor and sensory neurons after single-cell analysis (Al-Majed et al., 2004; Geremia et al., 2007). Regeneration-associated genes like GAP-43 are thought to be upregulated as a result of increased BDNF expression after ESTIM (Gordon et al., 2009). To elucidate if ESTIM exerts its effects in long-gap nerve tube repair via a mechanism similar to the mechanism postulated for ESTIM in combination with end-to-end neurorrhaphy, in the current study homogenized DRG or lumbar spinal cord tissue was analyzed 2 days and 2 weeks after a 13-mm nerve gap was bridged with cell-free biosynthetic grafts. Quantitative RT-PCR was used to elucidate if the same effects as those described above occur when the distal nerve stump or regeneration-promoting SCs are not immediately available to the regenerating neurons. A significant increase in BDNF mRNA levels related to ESTIM in lumbar DRGs or spinal cord was not detected. However, quantitative RT-PCR showed a more pronounced downregulation of BDNF mRNA levels 2 weeks after surgery in control animals that were not subjected to ESTIM. This indicates that ESTIM results in prolonged BDNF upregulation. This effect is likely to account for the better long-distance regeneration seen after ESTIM in the current study. Al-Madjed and colleagues previously demonstrated that together with increased BDNF expression, Tα1-tubulin is upregulated and neurofilament is downregulated, which presumably accounts for increased axonal growth (Al-Majed et al., 2004). Furthermore, ESTIM in the current study resulted in a significant upregulation of GAP-43 mRNA levels within the lumbar spinal cord 2 weeks after surgery, indicating that the processes of motor axonal regeneration had been activated to a higher extent after ESTIM than under control conditions. These results show that ESTIM has similar effects when applied in combination with long nerve gap reconstruction as in combination with end-to-end suture or short-gap bridging (Al-Majed et al., 2004; Geremia et al., 2007).
Regarding the potential for ESTIM to become a clinically useful technique in combination with already established microsurgical peripheral nerve repair techniques, a pilot study in patients with median nerve carpal tunnel release surgery did recently demonstrate the clinical applicability of brief low-frequency post-surgical ESTIM for the acceleration of axonal regeneration and target reinnervation in humans (Gordon et al., 2010). The results presented here provide further evidence of the utility and effectiveness of ESTIM for the induction of significant peripheral nerve-regenerating effects. In the future, brief ESTIM should be considered as an accompanying measure to improve functional outcome after nerve reconstruction surgery in humans. Appropriate pathway selection by regenerating axons of motor and sensory neurons has been reported to be accelerated by ESTIM, either combined with direct nerve transection and repair (Al-Majed et al., 2004; Geremia et al., 2007), or after end-organ deprivation of the distal stump for 3 weeks in rats (Wang et al., 2009). This indicates that ESTIM may be a useful tool to improve regeneration outcomes after delayed nerve autotransplantation. However, further investigation of this issue should be the subject of future studies.
Footnotes
Acknowledgments
This work was financially supported by the Stiftung Neurochirurgische Forschung of the German Society of Neurosurgery (DGNC), and the Hochschulinterne Leistungsförderung (HiLF) of the Hannover Medical School (both to K.H.T.). A scholarship was provided to R.S. by the Akademie für Tiergesundheit (ATF). Plasmids for genetic modification of Schwann cells were kindly provided by Prof. Claus (Institute of Neuroanatomy, Hannover Medical School). For excellent technical assistance we thank Silke Fischer, Natascha Heidrich, Gesa Hellmich, Kerstin Kuhlemann, Hildegard Streich and Maike Wesemann.
Author Disclosure Statement
No competing financial interests exist.