
Abstract
The glial scar formed at the site of traumatic spinal cord injury (SCI) has been classically hypothesized to be a potent physical and biochemical barrier to nerve regeneration. One longstanding hypothesis is that the scar acts as a physical barrier due to its increased stiffness in comparison to uninjured spinal cord tissue. However, the information regarding the mechanical properties of the glial scar in the current literature is mostly anecdotal and not well quantified. We monitored the mechanical relaxation behavior of injured rat spinal cord tissue at the site of mid-thoracic spinal hemisection 2 weeks and 8 weeks post-injury using a microindentation test method. Elastic moduli were calculated and a modified standard linear model (mSLM) was fit to the data to estimate the relaxation time constant and viscosity. The SLM was modified to account for a spectrum of relaxation times, a phenomenon common to biological tissues, by incorporating a stretched exponential term. Injured tissue exhibited significantly lower stiffness and elastic modulus in comparison to uninjured control tissue, and the results from the model parameters indicated that the relaxation time constant and viscosity of injured tissue were significantly higher than controls. This study presents direct micromechanical measurements of injured spinal cord tissue post-injury. The results of this study show that the injured spinal tissue displays complex viscoelastic behavior, likely indicating changes in tissue permeability and diffusivity.
Introduction
C
However, since Ramon y Cajal first hypothesized that the glial scar was a physical and chemical barrier to nerve regeneration in 1928 (Ramon y Cajal and May, 1928), no study has quantified the stiffness of the glial scar. The physical barrier hypothesis can be interpreted in the following two ways. One interpretation is that the scar has increased stiffness in comparison to uninjured neural tissue. Due to this elasticity mismatch, the scar acts as a physical barrier to nerve regeneration. To address this hypothesis, in an in vitro three-dimensional (3D) model of the glial scar, Yu and Bellamkonda (2001) demonstrated the reduced ability of embryonic dorsal root ganglia (DRG) neurites to cross the 3D interface of a mechanical barrier formed by a mismatch in the elasticity of layered agarose gels. The percentage of neurties crossing the interface decreased significantly as the modulus of the agarose gels increased. In the same study, Yu and Bellamkonda monitored the effects of chondroitin sulfate B (CS-B) on DRG neurite extension. The addition of CS-B to agarose gels lowered the shear modulus of the gels, but the CS-B-modified agarose gels were still highly inhibitory to DRG outgrowth. This result indicated that stiffness itself was not a potent inhibitor of neurite outgrowth, but that biochemical inhibition due to the CS-B prevented neurite extension across CS-B-rich interfaces. A second interpretation of Cajal's hypothesis is that the glial scar acts as a barrier to diffusion of growth-promoting molecules (Silver and Miller, 2004). In a study examining the diffusion of molecules across CSPG and reactive astrocyte-rich regions, Roitbak and Syková (Roitbak and Syková, 1999) demonstrated that the glial scar can inhibit diffusion between cells. Roitbak and Syková showed that diffusion was decreased in gliotic tissue, demonstrating that the scar can act as a diffusion barrier to growth-promoting molecules. Further, studies using diffusion tensor imaging have shown that the diffusivity across injured spinal cords is significantly altered in comparison to uninjured controls (Deo et al., 2006; Ellingson et al., 2008).
Due to a paucity of data on the material properties of injured rat spinal cords, the mechanical behavior of injured rat spinal cords was investigated, relative to uninjured control cords, under physiological conditions. A lateral hemisection model of SCI was employed and animals were sacrificed 2 weeks (2W) and 8 weeks (8W) post-injury (PI). Relaxation behavior of injured spinal cord tissue at the site of injury was assessed using compressive indentation testing. This mode of testing was chosen to elicit both a stiffness response (during the ramp phase), and a relaxation response (during the hold phase), which could address both facets of the physical barrier hypothesis, namely increased stiffness and decreased diffusion. We did not directly measure diffusion across the wound site. However, we estimated the viscosity of the wound site from the relaxation response, since Darcy's law states that permeability is inversely related to viscosity (Cowin and Doty, 2007). A modified standard linear model (mSLM) was fit to the data and inferences were made based on estimated model parameters. The SLM was modified to account for a spectrum of relaxation times, commonly observed in biological tissues (Fung, 1993), by the addition of a stretched exponential term, which is another unique feature of this study. The model presented here successfully fits the data. Further, it was observed that the stiffness and elastic modulus of injured spinal cord tissue is lower than control uninjured tissue, and that the relaxation time constants and viscosity of injured cords were higher than those of uninjured control tissue.
Methods
Hemisection surgery and spinal cord harvesting
The Department of Laboratory Animal Resources and the Committee for the Humane Use of Animals at SUNY Upstate Medical University, and the Institutional Animal Care and Use Committee at Syracuse University approved this study, following Association for Assessment and Accreditation of Laboratory Animal Care guidelines. Adult female Sprague-Dawley rats (average weight 255 g) were used for all experiments. The rats were anesthetized with ketamine and xylazine (80 mg/kg and 10 mg/kg, respectively), and body temperature was maintained using a heating pad. Aseptic surgery was performed using the following technique. The skin over the upper thoracic area was shaved and cleaned with povidone-iodine solution. The skin was incised, and then the connective and muscle tissue were bluntly dissected to expose the ninth thoracic (T9) vertebral body. A T9 laminectomy was completed, taking care not to damage the spinal dura during removal of the dorsal lamina. A lateral hemisection was performed at T9; initially a surgical needle punctured the spinal cord dorsoventrally at the midline, avoiding the dorsal spinal artery; angled microscissors were then used to cut the right half of the spinal cord, followed by scraping of the vertebrae with an angled needle ventrally and laterally surrounding the lesion to ensure completeness of the hemisection. Finally, the lesion was closed in layers with individual sutures. The animals were euthanized at 2W and 8W post-injury by perfusion under deep anesthesia with 1000 mL phosphate-buffered saline (PBS), followed by a dorsal laminectomy between the first cervical vertebra (C1) and the fourth lumbar vertebra (L4). The nerve roots were carefully severed and the spinal cord was cut at levels C1 and L4, then carefully removed and placed in a bath of isotonic PBS. No attempt was made to remove the dura mater. The cords were then stored in a refrigerator at 4°C until testing. All testing was performed within 4 h of excision of the spinal cords. Uninjured rat spinal cords were used as controls. A total of 18 animals were used for the study with 6 animals per group. However, due to improper dissection, only 5 spinal cords were used from the control and 2W post-injury groups.
Immunofluorescence and histological staining
Uninjured controls and experimental animals (2W and 8W post-spinal hemisection, n=4 per group) were euthanized with an intraperitoneal injection of sodium pentobarbital (50 mg/kg), followed by transcardial perfusion with PBS (500 mL) prior to perfusion with 4% paraformaldehyde in PBS (500 mL). Excised spinal cords were left overnight in 4% paraformaldehyde and then placed in a 20% sucrose solution in PBS for 2 days. Next, 8–10 mm of the spinal cords centered on the injured region were frozen in optimal cutting temperature (OCT) medium, and 20-μm serial sections in the transverse plane were cut using a cryostat. All serial sections were collected on glass slides and stained for immunofluorescence with the following antigens: glial fibrillary acidic protein (GFAP, 1:400 rabbit anti-GFAP; Dako, Carpinteria, CA), collagen IV (M3F7 supernatant, 1:10, mouse monoclonal anti-collagen IV; Hybridoma Bank, University of Iowa, Iowa City, IA), and CSPGs (CS-56, mouse monoclonal anti-chondrotin sulfate, C-8035, 1:100; Sigma-Aldrich, St. Louis, MO). Following overnight incubation at 4°C with primary antibodies, the sections were washed and incubated with the appropriate secondary antibodies: Alexa-Fluor 488 goat anti-mouse or anti-rabbit IgG (1:400), and Alexa-Fluor 594 goat anti-mouse IgM (1:400). Histological stains for myelin (eriochrome cyanine [EC]), and a triple stain (eosin, cresyl violet, and Luxol fast blue), for assessing general tissue structure were also employed.
Both fluorescence and bright-field imaging was done using a Zeiss Axioskop microscope (Carl Zeiss, Oberkochen, Germany). Digital images were captured using a Spot RT Slider camera (Diagnostic Instruments, Sterling Heights, MI), and analyzed with the accompanying Spot Advanced software (v.3.3.4 for Macintosh; Diagnostic Instruments). Images were captured at the same exposure times for each antigen-antibody pair across all groups, and the images were compared qualitatively to assess changes in tissue composition.
Relaxation testing
All testing of spinal cords was performed at room temperature while the spinal cords were completely submerged in PBS (isotonic, pH 7.4). Compressive indentation testing was performed using a custom built microindentation system (Saxena et al., 2009). Samples were oriented rostrocaudally with the dorsal surface facing up. The spinal cords were adhered to a steel sample stage using a slightly adhesive double-sided tape. The stage was then positioned under the indenter tip inside a glass holder. The sample and the tip were then fully submerged in PBS by filling the glass holder with PBS, and relaxation testing was performed after allowing the sample to equilibrate for 20 min. The indentation system operates in load control mode during loading, and the maximum load (25 μN) applied to the sample was the only controlled variable. The indenter tip was a spherical glass bead 336 μm in diameter, which was attached to an aluminum cantilever. Initially the tip and the sample were separated by a gap. The tip was moved towards the sample using a stepper motor and after contact, continued displacing the sample until the programmed load was reached. This constituted the ramping phase of the test. With the tip fixed at that position, load relaxation data and stage and sample displacement data were continuously collected using a reluctance transducer and linear variable differential transformer, respectively, for up to 100 sec at a frequency of 100 Hz. Starting at the rostral edge of the lesion and moving caudally, serial indents were made at locations 400 μm apart until uninjured portions of the sample were reached, as determined by visual inspection.
Unrelaxed elastic modulus estimation
The elastic modulus was determined from the ramping portion of the load-displacement curve. We calculate the unrelaxed elastic modulus (
where
where
Modified standard linear model and curve fitting
Load versus time data were collected for each sample at each indentation during testing. These data sets were then normalized by the final displacement, at the end of each individual test, to obtain the relative stiffness as a function of time (k(t)), for each indent. In order to analyze the relaxation phase of the experiment, the relaxation portion of each stiffness function was fitted by a modified version of the standard linear model (SLM). The SLM consists of a spring of stiffness
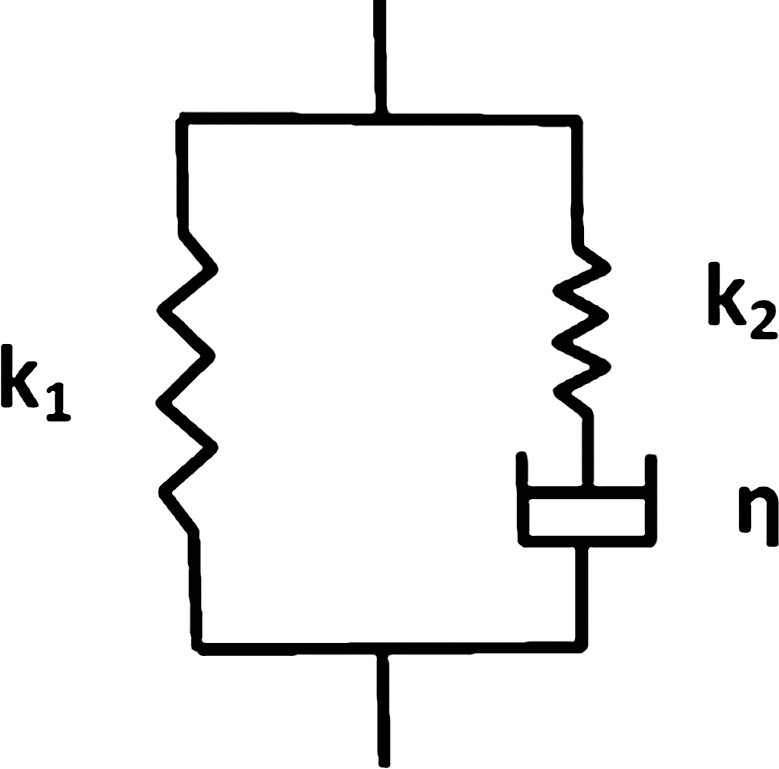
A schematic of the standard linear model. The spring of stiffness k1 is in parallel with a series combination of a spring of stiffness k2 and a dashpot with coefficient of viscosity η.
The solution for the relaxation response of the SLM is
where
where
In order to fit the model to the ramp phase, the Boltzmann superposition integral approach was used (Fung, 1993; Saxena et al., 2009), based on the parameters estimated from the mSLM fit to the relaxation data. Since [Eq. 4] is the relaxation function, according to the superposition principle:
where
Curve fitting the mSLM to the relaxation data
Parameters of the mSLM were estimated by numerically minimizing the sum of squared errors using the Levenberg-Marquardt algorithm (Abramowitch et al., 2004), using custom-written scripts in Matlab. The injured regions were 1–2 mm in width, and since indentation locations were visually monitored, data obtained from injured regions were pooled together to get a single set of fit parameters for each injured animal. In other words, if it was determined that the injured portion for a particular animal of a treatment group was 2 mm long, there would be 3–4 indents on the injured locations. After curve fitting, the fit parameters were pooled together to obtain a single set of representative fit parameters for injured regions for that animal.
Estimating viscosity from mSLM parameters
After estimating the mSLM parameters, the viscosity was calculated from load-displacement curves as follows. Analogous to the constitutive equation of the mSLM shown in [Eq. 4], we can write an expression for the relaxation modulus (Er(t)), which is proportional to the relaxation stiffness and given by:
where
and since stiffness is directly proportional to elastic modulus, f can also be expressed in terms of the moduli as follows:
Further, the unrelaxed modulus, Eu, which was determined from the ramping portion of the indentation experiment, is equal to the sum of the
Since Eu is known, we can now calculate ΔE from [Eqns. 8 and 9]. Since τ is known from the mSLM parameters, we calculate the viscosity (ηE) of the dashpot as
Frequency response of tissue from relaxation data using numerical Laplace transformation
Once the mSLM parameters were estimated, using the mSLM parameters for each treatment group, relaxation curves were generated for each animal per group. These curves were passed onto a custom script in Matlab, which performed a piecewise linear numerical Laplace transform on the relaxation curves (Bearinger et al., 2003; Gilbert and Dong, 1994; Gilbert, 1998), to generate real and complex stiffness and compliance values, along with phase angles for each group. The purpose of this approach was to obtain a measure of the frequency response of the tissue from time domain data. This is analogous to obtaining the response of the tissue to dynamic mechanical testing at different frequencies, at a given temperature. The parameters estimated were the real stiffness (k′), the complex stiffness (k″), and the phase angle theta (θ).
Statistical analysis
A one-way analysis of variance (ANOVA) was performed, followed by a Tukey's least significant difference post-hoc test to determine differences in model parameters and ramping slopes between groups with 95% confidence. All statistical analyses were done with SAS 9.0 software. All data are presented as mean±standard deviation.
Results
Stress relaxation testing of injured and uninjured cords
The control (uninjured) spinal cords had significantly higher unrelaxed modulus Eu (1248±293 Pa) than injured spinal cords (690±330 Pa for 2W PI, and 541±277 Pa for 8W PI cords). The representative ramping and relaxation behavior of injured cords and controls is shown in Figure 2. From Figure 2, it can be seen that the peak stiffness and ramping slope of uninjured cords are higher than those of injured cords (p<0.0005, Supplementary Fig. 1; see online supplementary material at
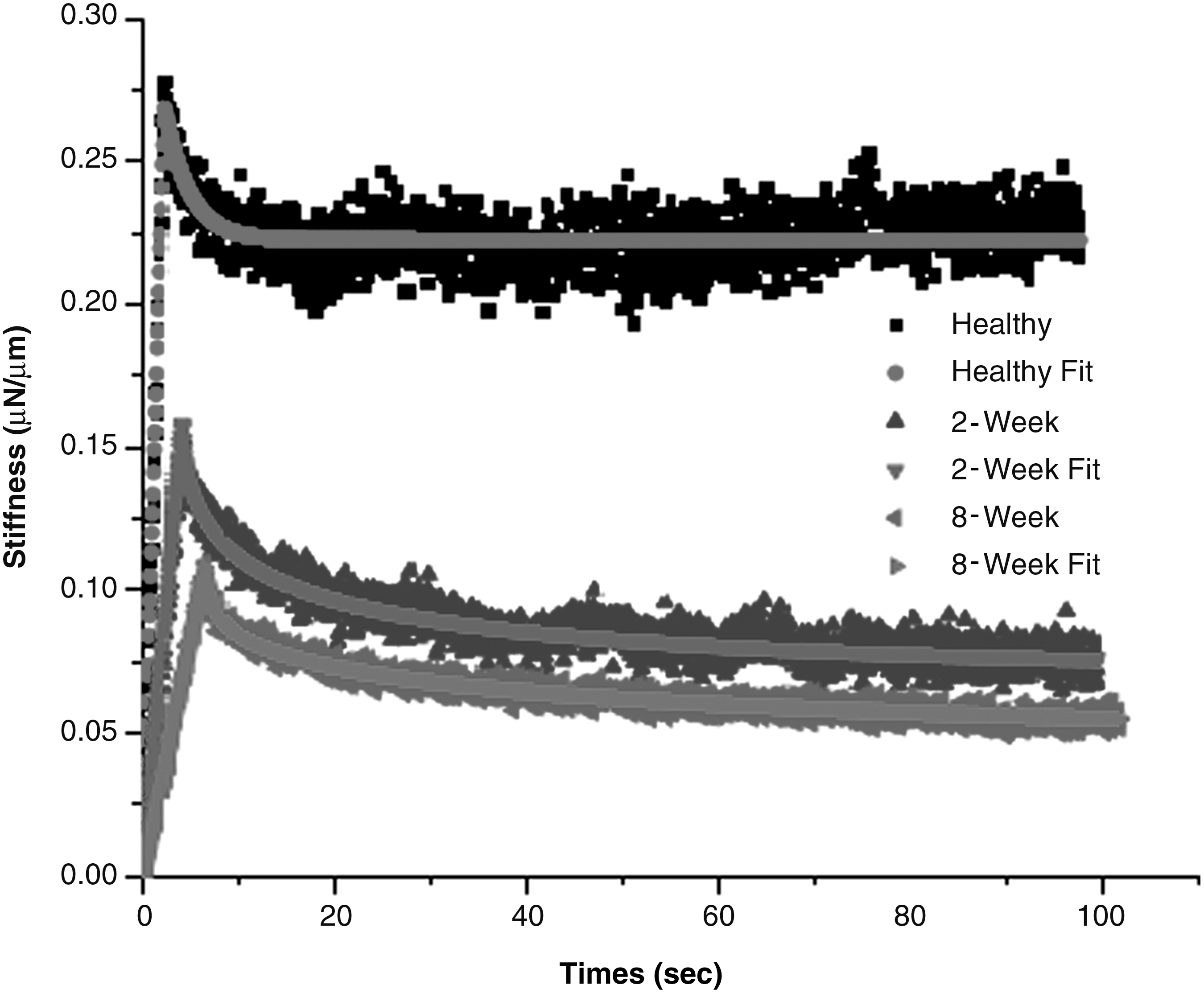
Representative raw data for the ramping and relaxation behavior of healthy, 2-week (2W) post-injury (PI), and 8-week (8W) PI spinal cords, and their corresponding modified standard linear model (mSLM) fits. The raw data were noisy. Every 10th point is plotted for clarity. It can be seen that the stiffness and ramping slopes of healthy cords is higher than that of injured cords. The model fits the data very well. The healthy cords have a rapid relaxation and reach equilibrium quickly, whereas the injured cords have a much slower relaxation in the given time period.
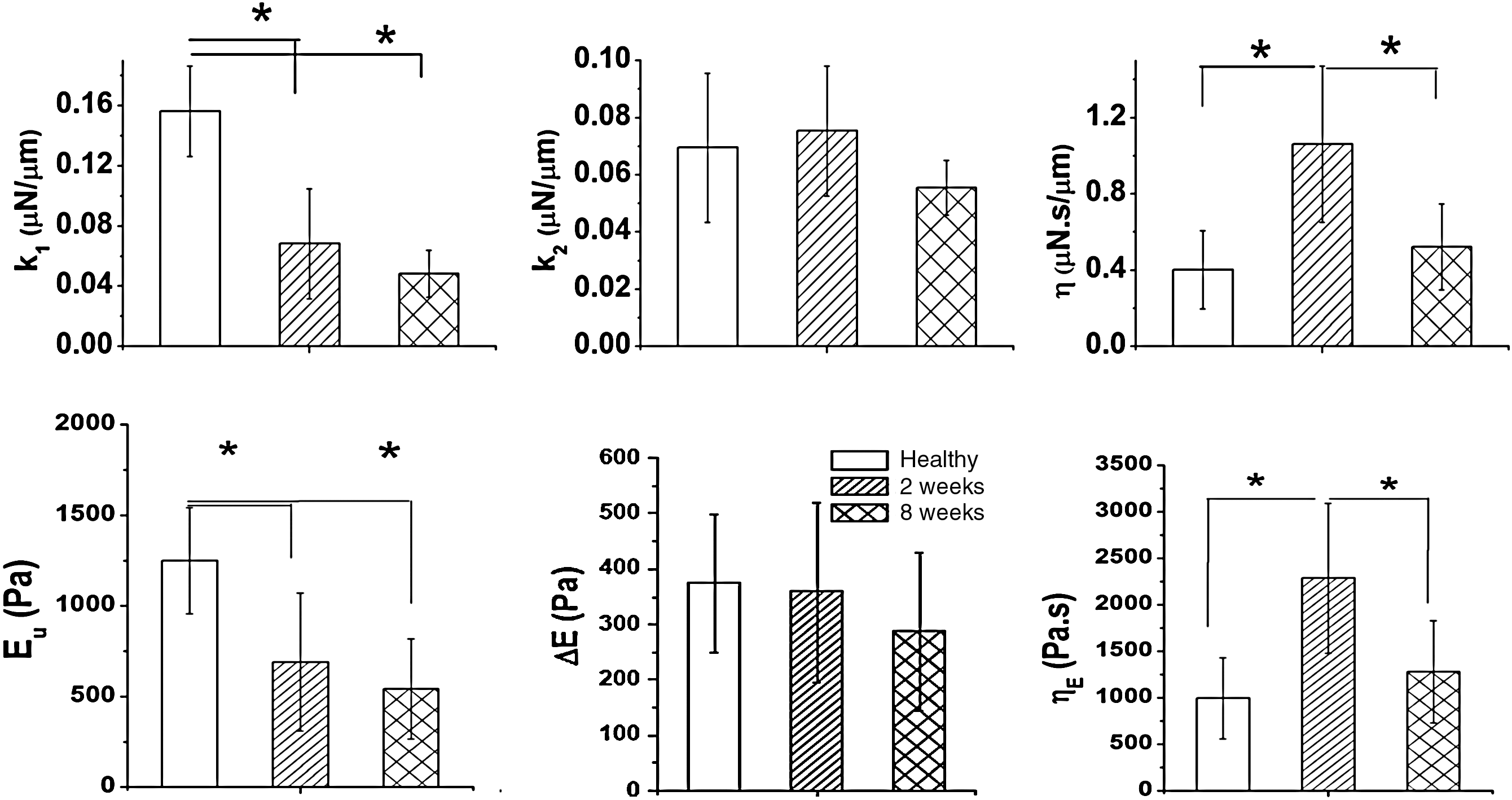
Comparison of various modified standard linear model (mSLM) model parameters, elastic modulus, and viscosity, between control and injured spinal cords. Significant differences between groups are indicated (*p<0.05). With respect to the mSLM,
Figures 4 and 5 show stiffness versus time data for a series of sequential indentations across a wound site in the 2W PI and 8W PI cords, respectively. Each graph represents a single spinal cord. At the top left corner of each figure, representative behavior of an uninjured spinal cord is shown for comparison. To clearly demonstrate the change in slope, as spinal cords were indented moving rostral (injured) to caudal (uninjured), data from just the first 15 sec of the test are shown. We noted that there is little variability in the control uninjured cords; however, there is clear variability in the injury groups. The 2W PI groups show a consistent trend toward increasing slope as the indents proceed in a caudal direction. This result was not consistent within the 8W post-injury group. We also observed that the uninjured cords have a steeper ramping slope and higher stiffness in comparison to both the injury groups.
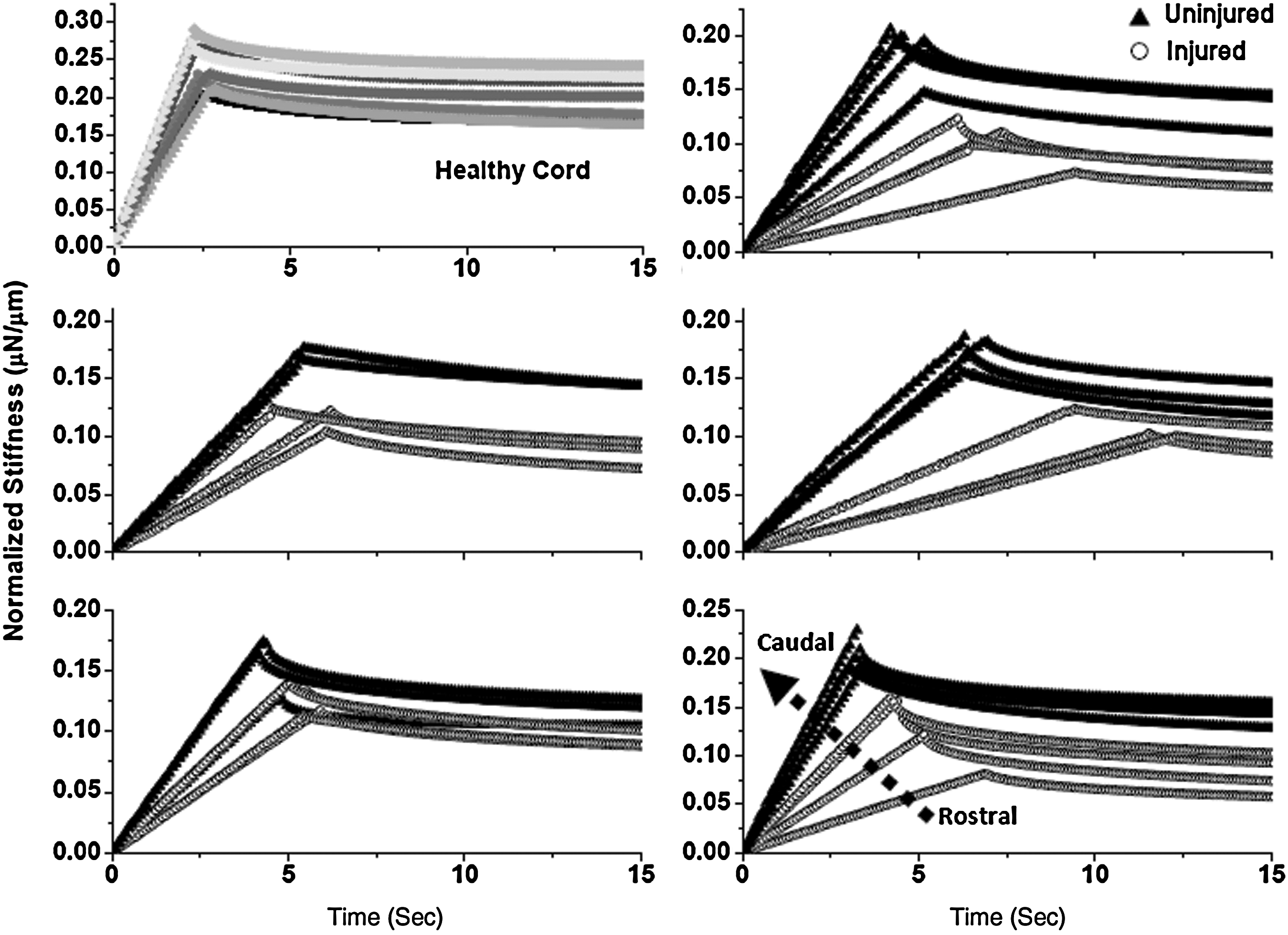
Relaxation behavior of indents across a wound site for 2-week post-injury rats. Model fits are shown for clarity and every 15th point is plotted. Representative behavior of a control healthy cord is shown in the top left corner. The stiffness and ramping slope of healthy cords is higher than that of injured cords. Stiffness and ramping slope increase moving from rostral to caudal. The dotted arrow indicates rostral-to-caudal direction. Refer to Figure 6 for associated histology.
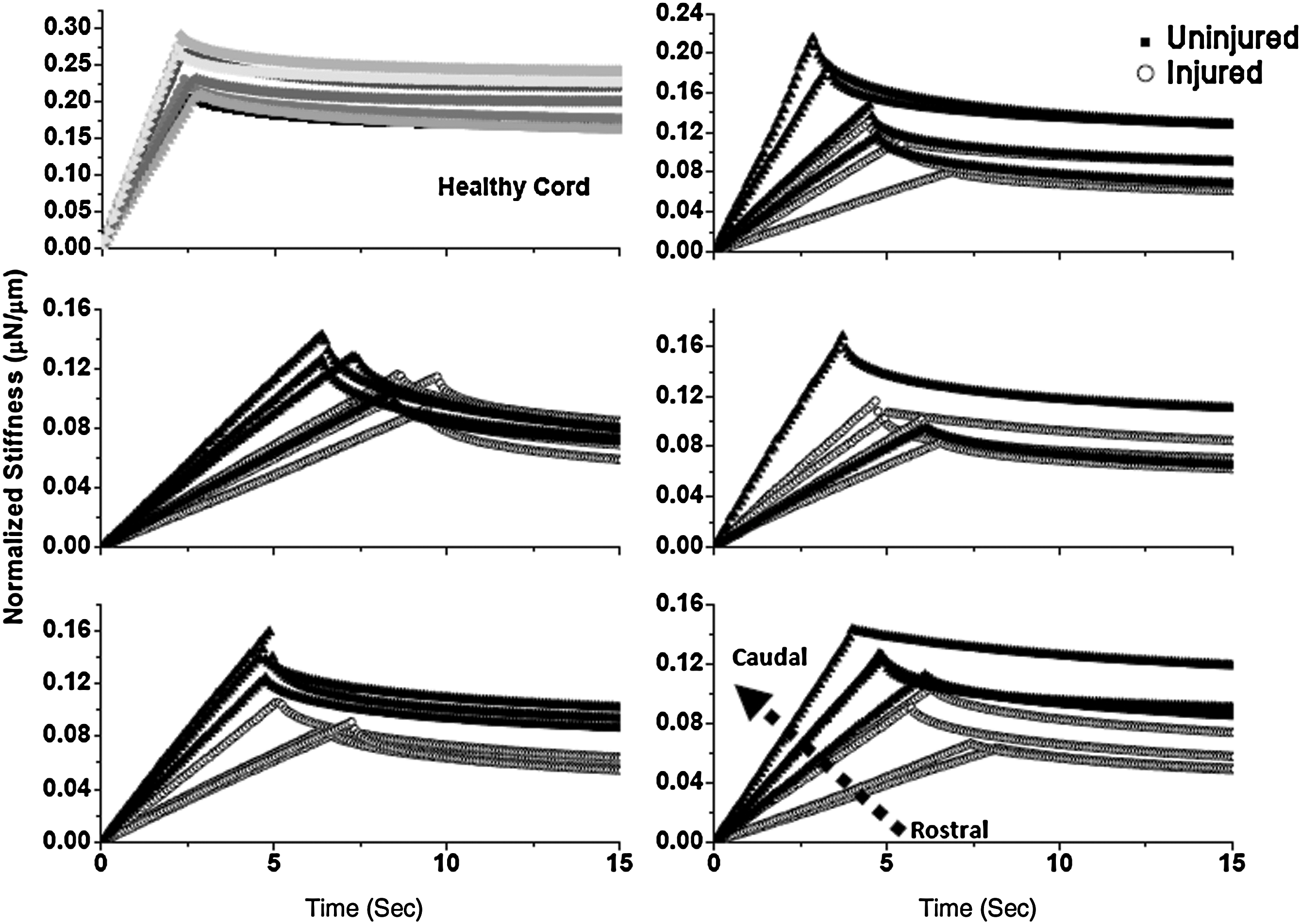
Relaxation behavior of indents across a wound site for 8-week post-injury rats. Model fits are shown for clarity and every 15th point is plotted. Representative behavior of a control healthy cord is shown in the top left corner. The stiffness and ramping slope of healthy cords is higher than that of injured cords. The dotted arrow indicates rostral-to-caudal direction. In comparison to the 2-week post-injury spinal cords, the mechanical behavior of the spinal cords was not as consistent. Refer to Figure 6 for associated histology.
Immunohistochemical and histological analysis of the lesion site
The gross structure of injured and control cords was visualized using an eosin, Luxol fast blue, and cresyl violet triple stain for connective tissue, myelin, and cell bodies, respectively. The representative micrographs are shown in Figure 6. Sections are spaced 500 μm apart, moving rostral to caudal starting at the lesion epicenter. It can be seen that myelination increases moving caudally. However, in 8W PI cords, cyst formation was noted. CSPG levels remained upregulated at 8W PI, although at lower levels than the 2W PI cords. Collagen IV was also upregulated in the lesion core at 2W PI and remained upregulated at 8W PI. GFAP staining was absent in the lesion core. The EC stain for myelin showed pronounced demyelination at 2W PI, which continued at 8W PI on the ipsilateral side of injury. Demyelination was not observed contralaterally. The Luxol fast blue, eosin, cresyl violet, triple stain clearly showed dense connective tissue formation at 2W PI and spinal cord atrophy at 8W PI (Supplementary Fig. 2; see online supplementary material at
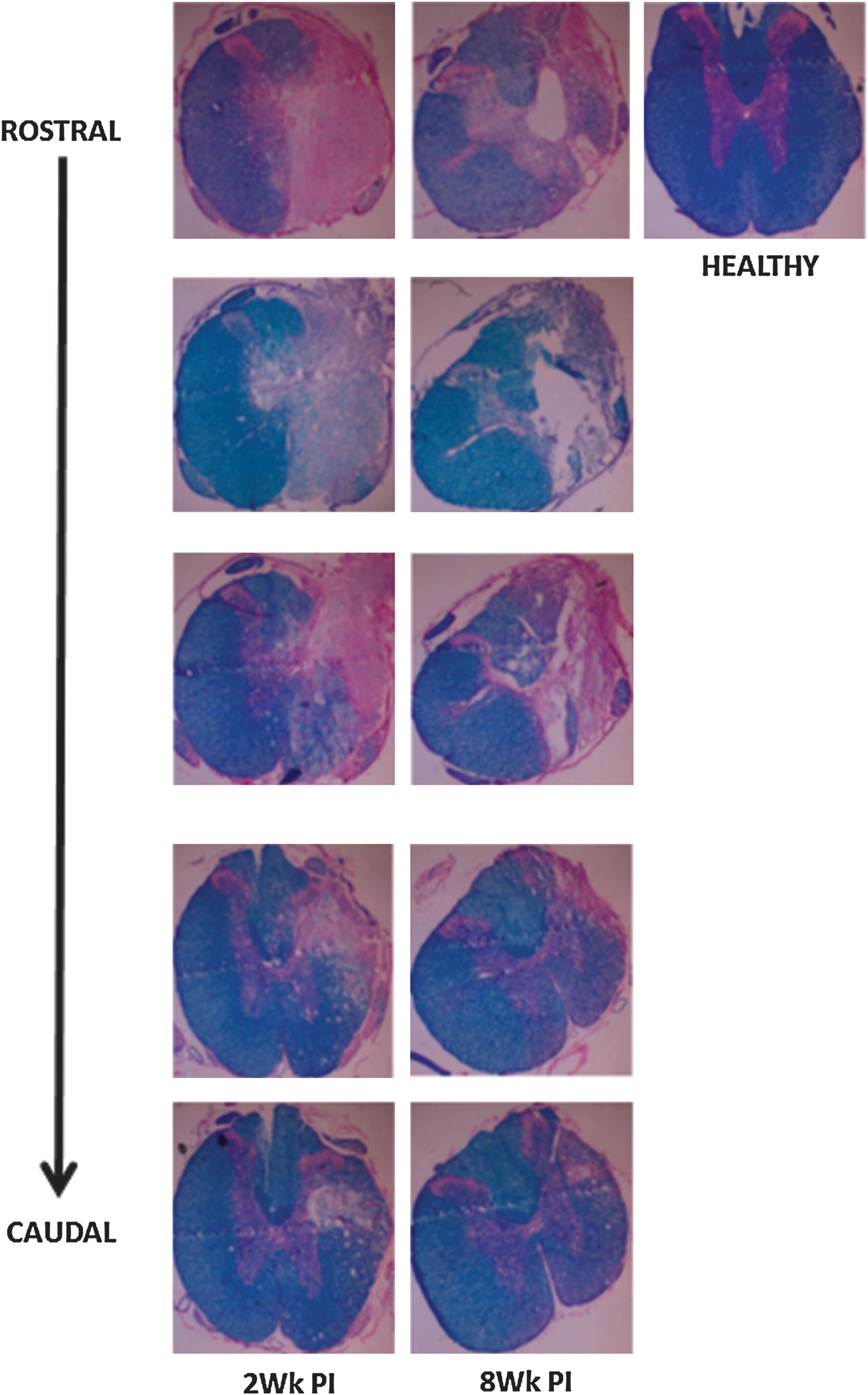
Representative Luxol fast blue, eosin, cresyl violet triple staining to show progression of spinal cord injury (SCI) in 2-week (2Wk PI, left column) and 8-week (8Wk PI, center column) injured cords. A section of healthy cord (right column) is shown as control. Sections are 500 μm apart. Top is rostral and bottom is caudal. Moving caudally, remyelination is observed and tissue integrity is restored with a decrease in connective tissue formation. Cyst formation and tissue loss is observed at 8 weeks post-injury.
Complex stiffness determination
The results from the numerical Laplace transformation of the relaxation data are shown in Figure 7. Figure 7a is a plot of the real stiffness (k′) versus the frequency, and Figure 7b is a plot of the complex stiffness (k″) versus the frequency for all three treatment groups. Figure 7c is a plot of the phase angle theta (θ) versus the frequency for all three treatment groups. It is clear that the injured cords have a larger phase angle than uninjured cords, indicating that the viscous nature of tissue increased upon injury.
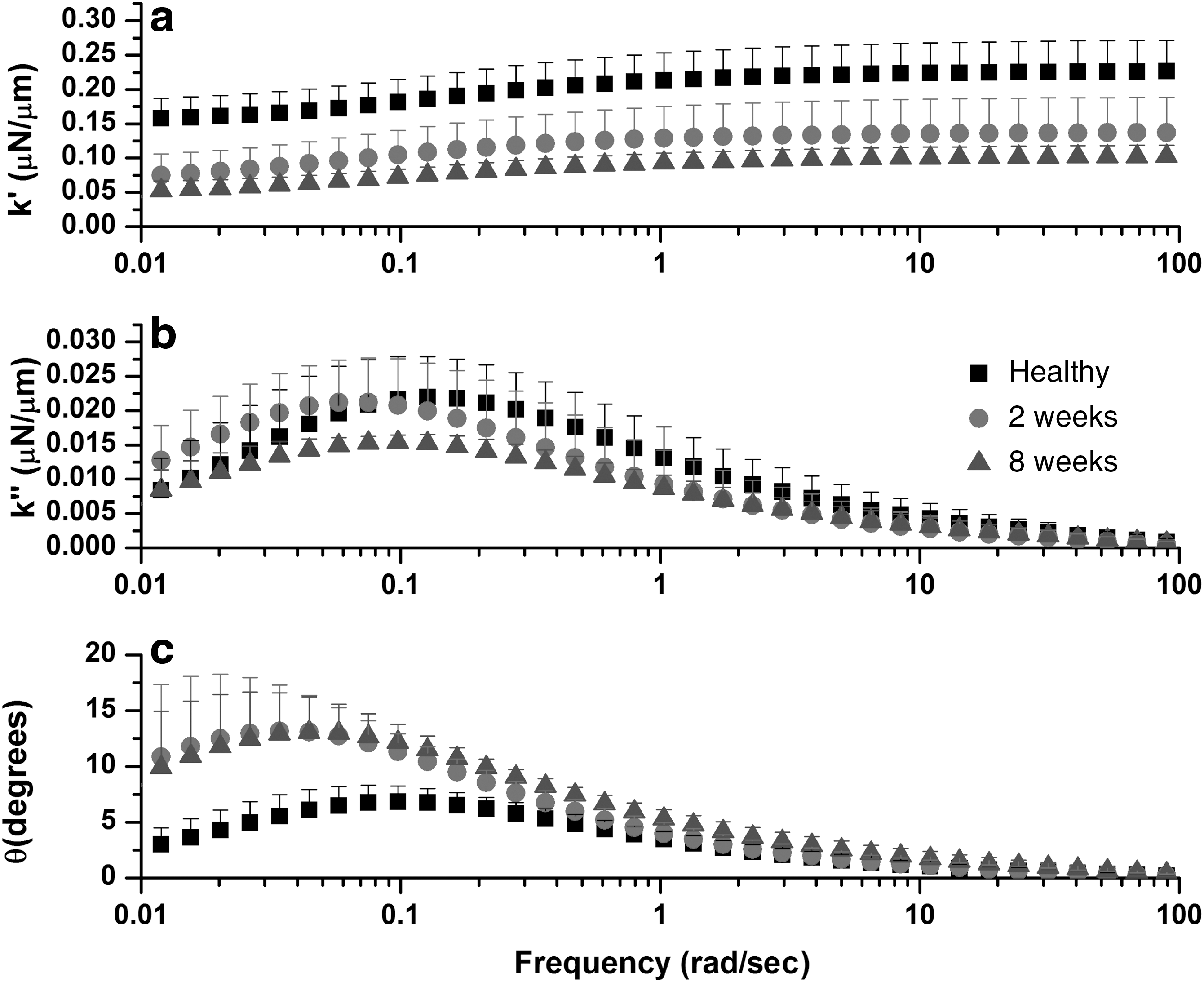
Results from the numerical Laplace transform of time domain data of the treatment groups. All plots are graphed on a logarithmic x-axis representing the frequency. All data are plotted as mean±one standard deviation for clarity. (
Discussion
The major finding of this study is that injured spinal cord tissue has significantly different material properties than uninjured tissue. This study is the first to report material properties of the injured spinal cord, in terms of the stiffness, unrelaxed elastic modulus, viscoelasticity, and frequency domain behavior. While the experiments conducted in this study do not allow direct correlation of these measured and calculated mechanical properties with discrete, spatially resolved, changes in the cellular and extracellular matrix composition of the injury site, there is clear evidence of altered mechanical properties of the spinal cord post-injury, along with structural changes in the tissue. The rather simple microindentation techniques utilized in this study, combined with the computationally facile application of the modified standard linear model (mSLM) and the Laplace transformation, resulted in a rich set of mechanical property data to describe the relaxation behavior of healthy and injured spinal cord tissue.
The results of our study indicate that the unrelaxed elastic modulus (Eu) and stiffness (k1) of injured spinal cord tissue were significantly lower in comparison to uninjured tissue. This result is consistent with the results of a recent study by Shafieian and associates (Shafieian et al., 2009), who observed a significant reduction in the instantaneous elastic force in injured brain tissue, which was also studied using indentation-based stress relaxation tests. There are distinct changes in the ECM composition at the injury site, that likely account for this decrease in stiffness and modulus. There was an increase in CSPG expression at the injury site at both 2 weeks and 8 weeks. CSPGs are known to bind water molecules tightly (Gray et al., 1999), thus a higher concentration of water at the injury site may contribute to the lower stiffness and modulus measured in our study. Indeed, Nesic and colleagues have reported that water content remains high even in chronically injured spinal cords (Nesic et al., 2006). Another factor that affects the elastic modulus of spinal cord tissue is white matter integrity. We observed robust demyelination at the lesion site at both 2 weeks and 8 weeks in our histological analysis. Shreiber and co-workers (Shreiber et al., 2009) reported that both demyelinated spinal cords and spinal cords with depleted glial cell populations demonstrated significantly lower stiffness and ultimate tensile stress than myelinated spinal cords. Thus, chronic demyelination could be a significant factor in the decreased stiffness and modulus measured in this study.
Changes in the cellular composition of the injured tissue may also have an effect on the stiffness and modulus. Miller and associates (2009) observed that the elastic modulus of cultured reactive astrocytes was significantly lower than naïve, uninjured astrocytes, and that this softening was not limited to the injury region, but also spread to the penumbra. Further, other researchers have shown that there is a widespread astrocytic reaction, as assessed by the upregulation of GFAP, which may extend for up to 1 cm from the injury in rats (Fawcett and Asher, 1999). In accord with these results, we noted that as the tissue was indented across the lesion, moving rostral to caudal, there was a progressive increase in the stiffness of the tissue (Figs. 4 and 5). However, the maximum stiffness of the regions surrounding the wound site, which visually appeared to be uninjured, was lower than that measured in control tissue (Figs. 4 and 5), indicating some degree of softening beyond the core of the injury site.
Rather than being a physical barrier to regeneration, as classically hypothesized, localized softening of the spinal cord tissue at the injury site, as measured in this study, may actually present a mechanically permissive substrate for nerve regeneration. It has been demonstrated in several previous in vitro studies that neurons prefer to grow on softer substrates, exhibiting significantly more neurite extension and branching (Balgude et al., 2001; Discher et al., 2005; Flanagan et al., 2002; Georges et al., 2006; Lu et al., 2006; Yu and Bellamkonda, 2001). Additionally, Miller and colleagues (2009) hypothesized that reactive astrocytes could be a mechanically favorable substrate for neuronal regrowth and repair in the absence of other inhibitory factors (Pekny and Nilsson, 2005; Pekny et al.,1999). However, different subsets of neurons have a different regenerative response to injury. Therefore, factors other than stiffness must be critical for aborting axonal regrowth (Fitch and Silver, 2008).
The spinal cord tissue tested in this study displayed complex viscoelastic behavior, as determined from the mSLM parameters and the frequency domain behavior of tissues. We observed a significant increase in the viscosity (ηE), the coefficient of viscosity (η), and the relaxation time (τ) of injured spinal cord tissue at 2W PI. We also observed a larger phase angle from the Laplace transform analysis in the injured groups. We hypothesize that these increases are due to the increased expression of proteoglycans and collagen IV at the injury site. It was observed that CSPG expression was maximal at 2W PI, which correlates with results of other published studies (Jones et al., 2003), and continued to be higher than control levels even at 8W PI. Large proteoglycans and hyaluronic acid (HA) increase tissue viscosity (Bothner and Wik, 1987; Gray et al., 1999). Also, the binding of HA with other CSPGs has been shown to increase the viscosity of HA-CSPG solutions (Tanaka et al., 2003). Roitbak and Syková (1999) reported that after injury to the cortex, in regions of glial scarring, increased tortuosity was a characteristic feature of injured brain tissue. They attributed this to the diffusion barriers interposed between the cells due to astrocytic hypertrophy, and a variety of ECM macromolecules such as laminin, collagen IV, fibronectin, and CSPGs produced by reactive glial cells that could form additional diffusion barriers and result in increased tissue viscosity (Roitbak and Syková, 1999; Syková et al., 1999).
We hypothesize that an increase in the viscosity of spinal cord tissue following injury likely correlates with a reduction in tissue permeability at the injury site. While we did not measure permeability of the tissue directly, this hypothesis is in accord with results reported by Roitbak and Syková (1999), and Darcy's law, which states that permeability is inversely related to viscosity (Cowin and Doty, 2007). Decreased tissue permeability at the site of spinal cord injury is physiologically advantageous in the acute phase of injury, limiting the diffusion of neurotoxic agents out of the lesion site, thus providing neuroprotection (Fawcett and Asher, 1999; Renault-Mihara et al., 2008; Rolls et al., 2009; Silver and Miller, 2004). However, this decrease in permeability may also limit the diffusion of growth-promoting factors at the lesion site, thus acting as a secondary inhibitory factor in axonal regeneration.
In this study we utilized a microindentation-based stress-relaxation test to measure the viscoelastic properties of injured and uninjured rat spinal cord tissue. We observed that the indent depths for injured cords were on the order of 220 μm, and that those of uninjured controls were on the order of 140 μm. We recognize that this represents a high-strain situation for injured cords. However, the modulus values obtained for uninjured cords are in agreement with a previously published study (Shafieian et al., 2009), and thus it is reasonable to use this technique to make relative comparisons between healthy and injured tissue in this study. Another limitation of this study is that spinal cords were tested in the dorsal-ventral plane, as opposed to the rostral-caudal orientation situation in vivo. Stress-relaxation testing of the rat spinal cord has traditionally been performed in tension, by orienting the spinal cord along its longitudinal axis (Clarke et al., 2009; Fiford and Bilston, 2005). The spinal cord is in tension in vivo, and also has perfusion pressure exerted due to the cerebrospinal fluid and blood circulation. While we tested the spinal cords completely submerged in PBS, the lack of perfusion pressure could lead to material properties that are distinctly different from in vivo conditions. While the elucidated properties may differ from in vivo properties, this should not have affected the relative behavior of injured and uninjured spinal tissue.
A hemisection injury model was used in this study; however, this type of injury accounts for a relatively small proportion of spinal cord injuries in humans, where contusion injuries are much more common (Bunge et al., 1997; Iannotti et al., 2006). The hemisection model was chosen due to the focal scar formation and relatively low incidence of cyst formation at short times, even though we did see cyst formation in some animals at the chronic stage (8W PI) of SCI (Siegenthaler et al., 2007). Additionally, the hemisection model has been well characterized by other investigators, and our immunohistological findings are similar to those of several other studies reported in the literature (Busch and Silver, 2007; Fawcett and Asher, 1999; Hermanns et al., 2001; Klapka and Müller, 2006; Klapka et al., 2005; Siegenthaler et al., 2007; Silver and Miller, 2004; Stichel et al.,1999). Further, the glial scar formed following hemisection consists of both connective tissue and glial elements. Thus, it was impossible to discern changes in central nervous system tissue exclusively, due to glial activity from contributions by connective tissue formation. Future studies will focus on a contusion model of SCI, which is more clinically relevant, and has a more severe glial reaction, but could lead to difficulty in mechanical testing, since it also leads to widespread tissue damage and cyst formation (Siegenthaler et al., 2007). Finally, the size scale of testing could also have influenced the results (Ebenstein and Pruitt, 2004). The diameter of the indentation probe was 336 μm. Perhaps at smaller size scales, for example at those on the order of individual cells, the individual effects of various wound remodeling elements on the observed mechanical properties could be assessed more accurately.
Conclusions
In summary, we found that the spinal cord tissue at the site of injury has a lower stiffness and modulus than uninjured tissue, but that it displayed increased viscosity compared to adjacent uninjured tissue using a lateral microindentation test method. The stretched exponential form of the mSLM provides a facile means of characterizing the viscoelastic behavior of these tissues in time-dependent relaxation experiments. The results of this study indicate that the healing wound site is perhaps a physical barrier to regeneration due to its viscoelastic nature, which likely poses a diffusion barrier. It is not simply a stiffness barrier to regeneration.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.