
Abstract
The exact mechanism of neuroma-associated pain is not yet fully understood, thus contributing to the substantial challenge faced in managing patients with painful neuromas. We aimed to observe the expression of alpha smooth muscle actin (α-SMA) in the painful traumatic neuroma and to investigate its possible roles in the cause of neuroma-associated pain. Its expression is considered to be a useful phenotypic marker for myofibroblast, and may contribute to its increased contractile activity. We collected peripheral neuroma specimens prospectively and subsequently divided them into two groups: painful (n=21) and non-painful (n=27) based on blinded preoperative visual analogue scale (VAS) pain scores. We also harvested normal nerve specimens from the discarded limbs as a control group (n=8). We performed immunohistological studies to observe the expression of α-SMA in each group, and calculated the expression level by a high-resolution pathological image analysis system. There was no positive staining of α-SMA observed in the control group, slight positive staining in the non-painful group, and obviously positive staining in the painful group. Pearson correlation analysis demonstrated that VAS scores were significantly associated with the expression intensity of α-SMA (R=0.831; p<0.001). Linear regression analysis indicated that the expression intensity of α-SMA was positively related to the scale of VAS (R 2=0.691, p<0.001). These findings suggest that: 1) expression of α-SMA may play certain roles in painful traumatic neuroma, either as a direct cause of neuroma-associated pain or as an indirect marker of local mechanical stimuli, and 2) the presence of α-SMA in the painful group may provide rationale for transpositional procedures in the management of traumatic neuroma. The persistent existence of α-SMA in the painful group and the correlation with VAS scores may provide insight into the development of new therapeutic strategies.
Introduction
P
The exact mechanism of neuroma-associated pain is not yet fully understood, contributing to the challenge of managing patients with painful neuromas. 6 There are several theories proposed to explain the mechanisms or pathophysiology of this situation, including the persistent mechanical or chemical irritation of the axons, persistent stimulation of the axons within the neuroma via spontaneous discharge, and the gate theory. 7 However, none of these theories can explain this phenomenon completely. 5 Nonetheless, mechanical entrapment of regenerated unhealthy nerve fibers by surrounding scar tissues is generally considered to be the most plausible cause of painful neuromas, and serves as the rationale for treatment modalities that transfer the nerve stump into muscles, veins, or bone. 8
In addition to serving as a phenotypic marker of myofibroblast activity, the expression of α-SMA may also contribute to the increased contractile activity of the myofibroblast. 9,10 In view of the self-contraction ability of the myofibroblast and its role in the development of scar contracture in wound healing, 11,12 we hypothesize that scar tissue within the neuroma may also crush the regenerated nerve fibers (internal compression), especially when contractile myofibroblasts are present, leading to the persistent neuroma-associated pain. The aims of this study are to quantify the expression of α-SMA in the painful neuromas and to investigate its possible roles in the mechanism of neuroma-associated pain.
Methods
Clinical data collection
Full approval of the research protocol was obtained from our institution's scientific review committee. All patients provided informed consent prior to neuroma specimen collection.
Neuroma specimens were obtained from patients undergoing secondary management of stumps following limb or digital amputation, including secondary stump plasty, painful neuroma excision, and digital reconstruction. The patients with ongoing neuromatous pain were assigned to the painful group (n=21) and those without pain were assigned to the non-painful group (n=27). The pain status had been scored on a 10 cm visual analogue scale (VAS), ranging from 0 “no pain at all,” to 10 “unbearable pain” 8 before the patients underwent neuroma excision. All assessments of the pain status were made by the same blinded observer. In addition, a sample of normal nerve specimens (control group, n=8) were harvested from the discarded limbs in patients who underwent unavoidable limb amputations as a result of mutilated limb injury or vascular injury. The mean period after the primary surgery was 5.5 months (range, 3–15 months) in the painful group and 6.1 months (range, 3 –19 months) in the non-painful group, respectively. The majority of the specimens (31/56) were collected from the digital nerves (DN, control group n=4, non-painful group n=14, and painful group n=12), and the others (25/56) were harvested from median nerves (MN), ulnar nerves (UN), tibial nerves (TN), and sciatic nerves (SN), respectively. Detailed clinical data for all patients were provided in Tables 1 and 2.
DN, digital nerve; MN, median nerve; UN, ulnar nerve; TN, tibial nerve; SN, sciatic nerve.
Chi-square test showed that no significant differences were observed between the two groups regarding personal history, type of injury, and comorbidities (p>0.05).
Immunohistological study
After a minimum of 48 h of fixation in 10% formaldehyde solution, the tissue was processed by standard paraffin-embedding methods. Sections were cut at 4 μm. Standard hematoxylin and eosin (H&E) staining was performed. In order to standardize the site for immunohistochemical staining, the sections were randomly selected at a distance ranging from 400 to 600 μm from the distal ending of the specimens. At least 30 sections were obtained from each sample, but after small sections or poorly cut sections were excluded, ∼20 sections were obtained. Out of the 20 sections, 5 were randomly selected. Monoclonal antibodies against the following proteins were used: the monoclonal mouse anti-human α-SMA (Sigma, USA) and secondary monoclonal goat anti-mouse antibody (Sigma, USA). Streptavidin–peroxidase (SP) immunohistochemical staining (Wuhan Boster Bio-engineering Company Limited, China) was used according to manufacturer instructions. To examine the specificity of immunostaining, phosphate-buffered saline (PBS) was used to replace the primary antibody as the control and the specificity was further verified by the immunohistochemical response of the small vessels in all the sections for the staining of α-SMA. 13 The slides were observed under a light microscope, and brown-yellow color staining was taken as positive expression.
Image analysis
HPIAS-1000, a high-resolution pathological image analysis system 14 was used for the quantitative analysis of α-SMA expression. A blinded investigator made all assessments, and three observation regions were randomly selected in each section. The mean integral gray values reported by HPIAS-1000 were used for subsequent statistical analysis.
Statistical analysis
Descriptive statistics and univariate analyses (one-way analysis of variance for quantitative data and χ2 test for binomial data) comparing the painful, non-painful, and control groups were performed using SPSS 17.0. Correlation analysis between the expression intensity of α-SMA and VAS scores were performed using Pearson's correlation coefficients and linear regression. A threshold of p<0.05 was used to determine statistical significance.
Results
There were no significant differences in demographic data between the painful and non-painful groups (Tables 1 and 2). H&E staining revealed a disorganized tissue structure with haphazardly arranged nerve fascicles mingled with extraneural collagen fibers, represented by plenty of connective tissues in both experimental groups. The regenerated nerve fibers were observed to have different morphologies, including bundles of almost normal myelinated nerve fascicles and some dispersed irregular myelinated and unmyelinated fibers. These fibers were not easy to distinguish and quantify; however, no noteworthy differences between the two experimental groups were revealed grossly (Figs. 1 and 2).
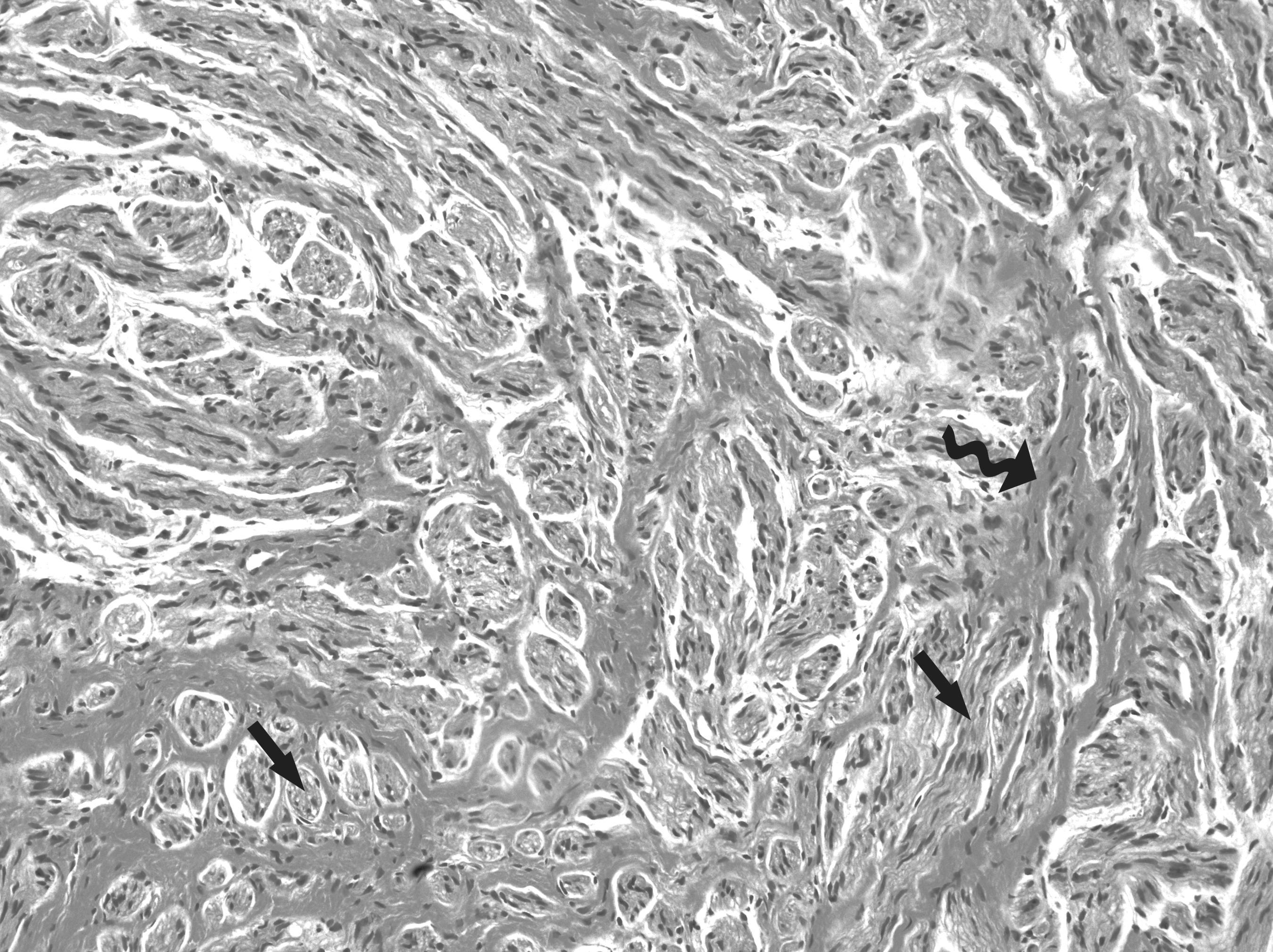
Histological demonstration of non-painful group (×20). Hematoxylin and eosin (H&E) staining show that regenerated nerve fibers are surrounded by extraneural fibrosis (top, curved solid black arrow). The regenerated nerve fibers show different modalities, including bundles of almost normal myelinated nerve fascicles (left, straight solid black arrow) and some dispersed irregular nerve fibers (right, straight solid black arrow).
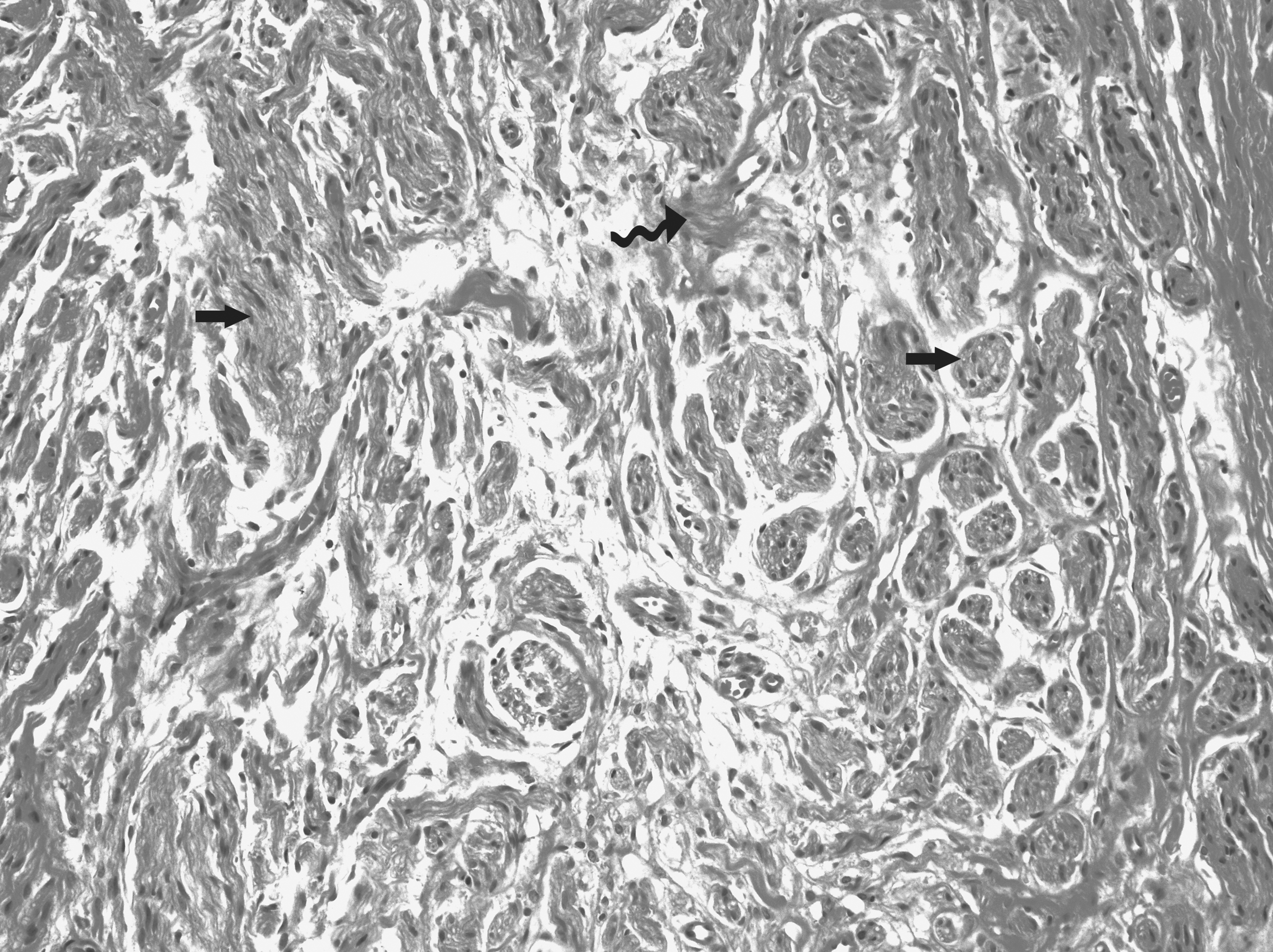
Histological demonstration of painful group (×20). Hematoxylin and eosin (H&E) staining show that regenerated nerve fibers are surrounded by extraneural fibrosis (middle, curved solid black arrow). The regenerated nerve fibers show different modalities, including bundles of almost normal myelinated nerve fascicles (right, straight solid black arrow) and some dispersed irregular nerve fibers (left, straight solid black arrow).
The immunostaining results showed no positive staining of α-SMA, except that blood vessels displayed a highly stained circular distribution in the control group. In the non-painful group, only slight positive staining of α-SMA was noted. Obvious positive staining of α-SMA was present in the painful group. The area of staining was located in the cytoplasm and distributed diffusely in the proliferative myofibroblasts among regenerated nerve fibers (Figs. 3 –5).
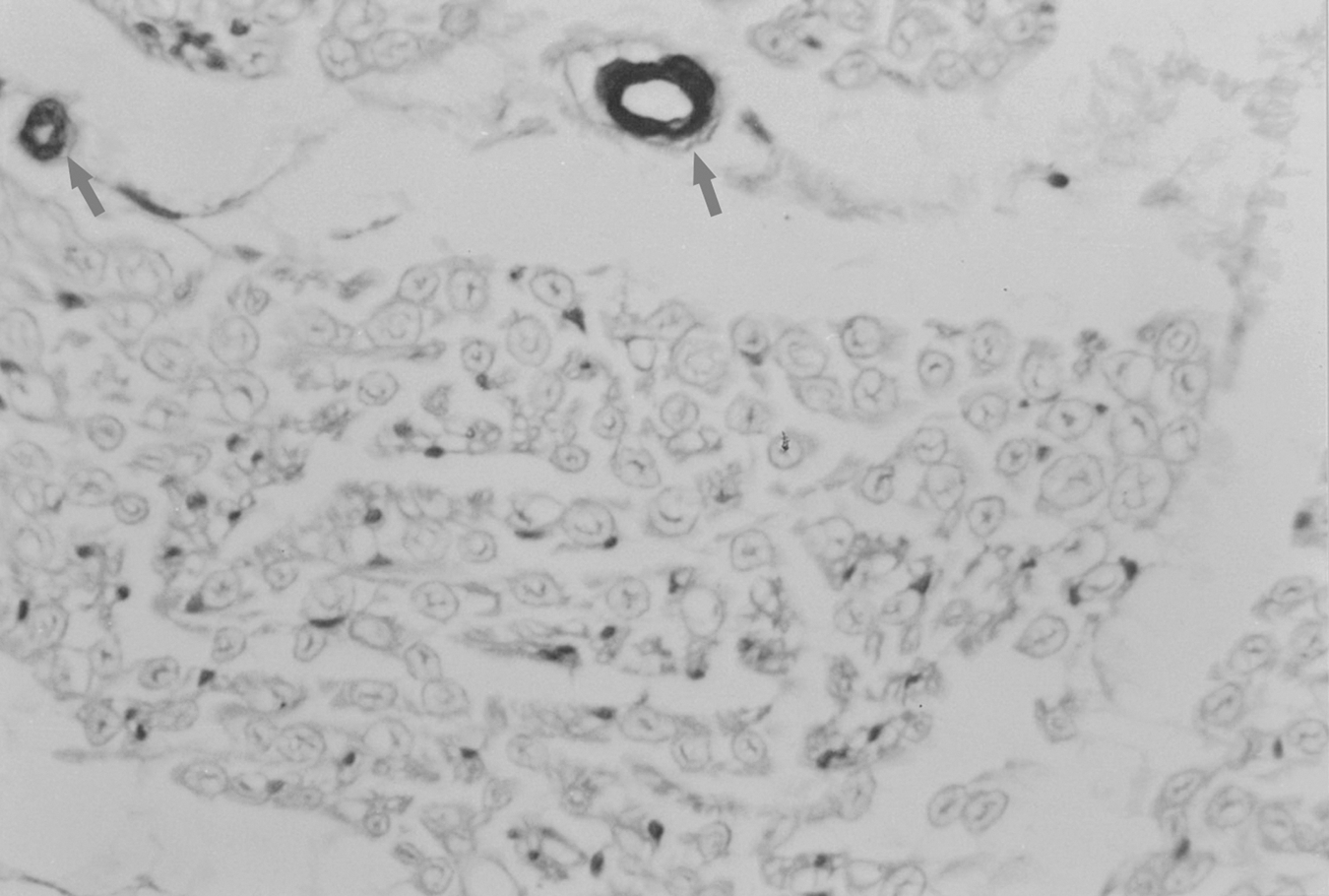
Immunostaining of alpha smooth muscle actin (α-SMA) in the control group (×40). Positive staining simply presented within vascular walls. The arrows show the highly stained blood vessels.
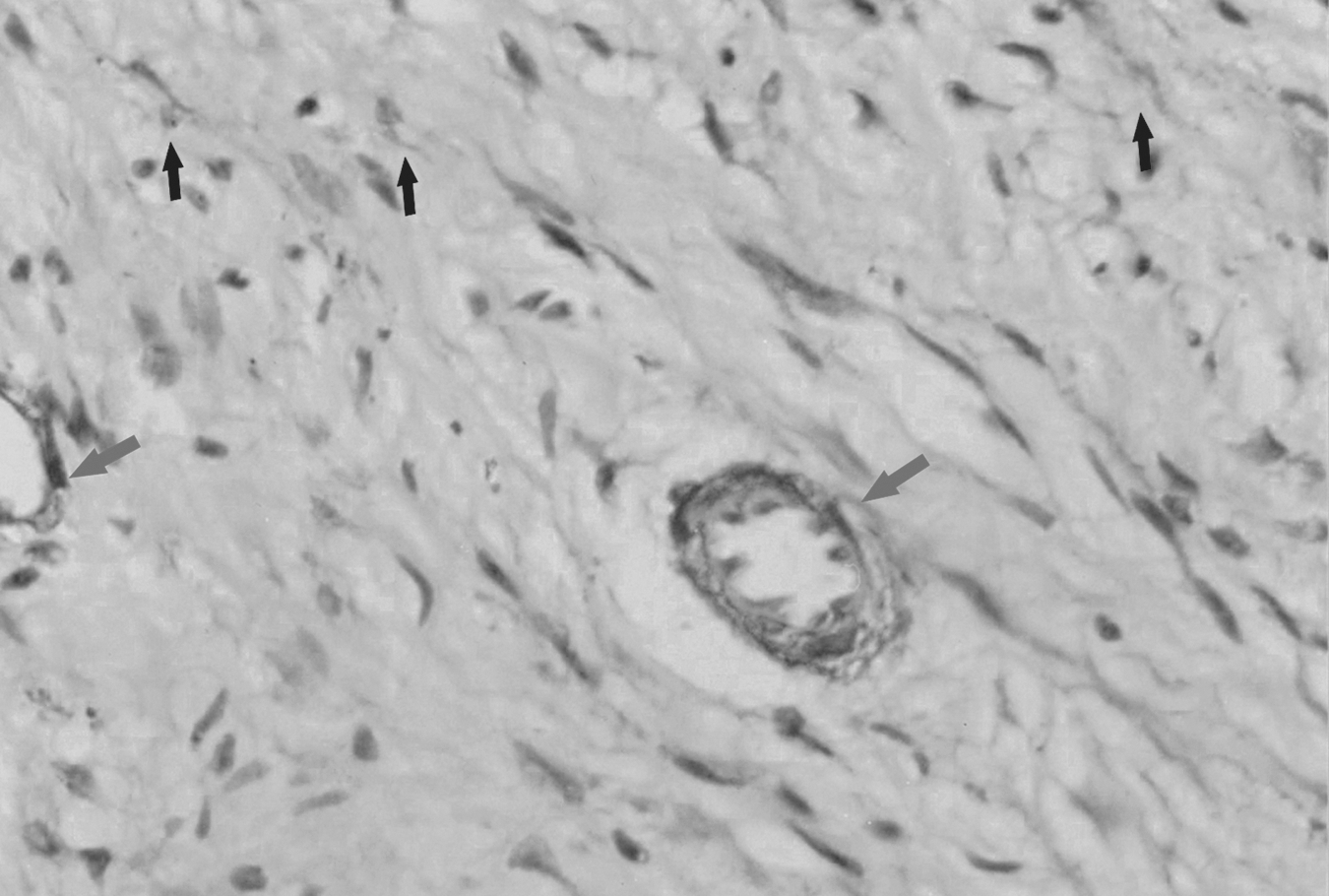
Immunostaining of alpha smooth muscle actin (α-SMA) in the non-painful group (×40). Only slight expression of α-SMA is observed. The arrows show the highly stained blood vessels and the black arrows show the dispersed positive staining of α-SMA.
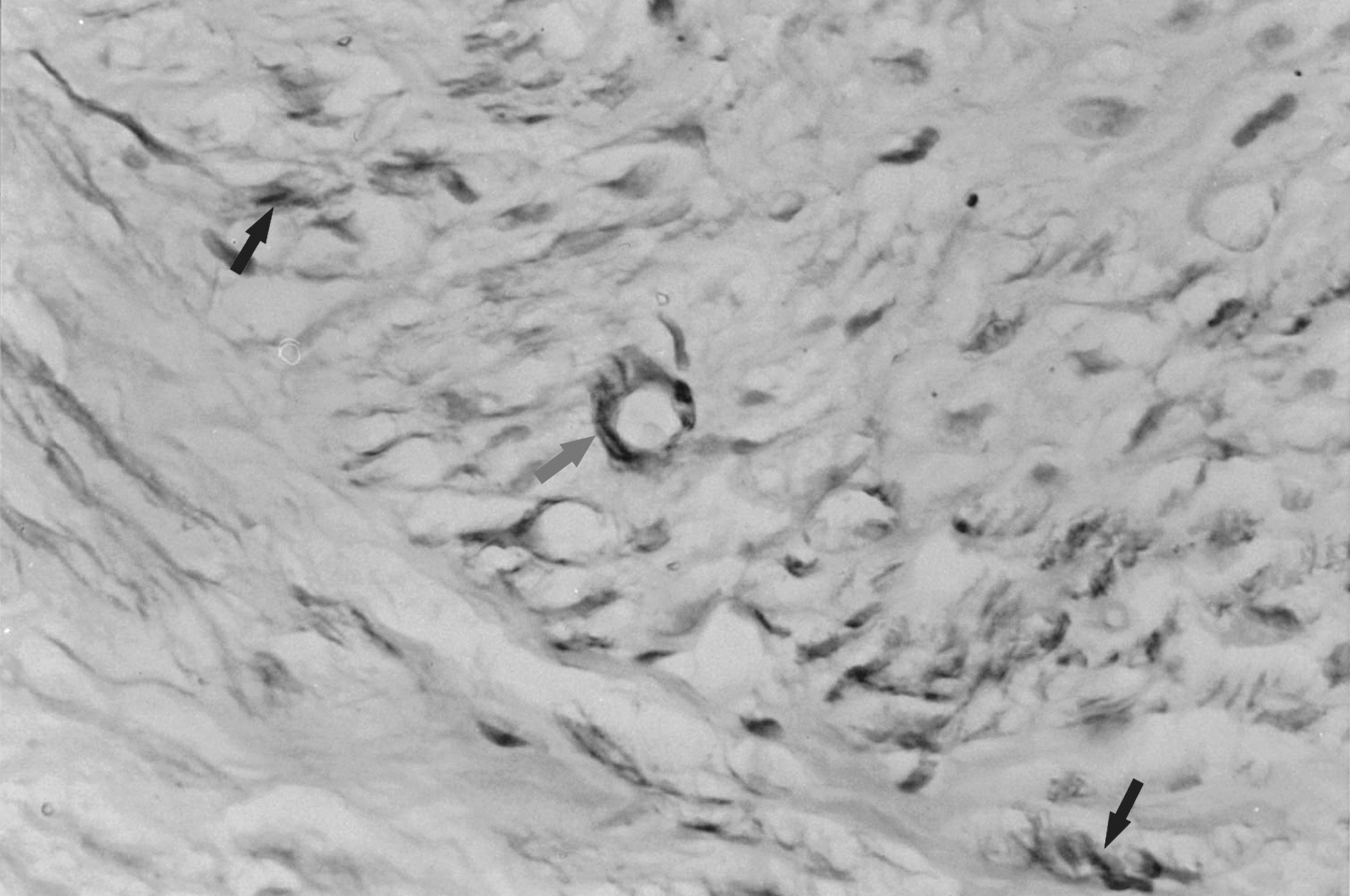
Immunostaining of alpha smooth muscle actin (α-SMA) in the painful group (×40). Obvious high expression of α-SMA is noted. The arrow shows the highly stained blood vessels and the black arrows show the highly stained myofibroblasts.
According to the results of HPIAS-1000, there was a significant difference in the mean gray values among the three groups (p<0.001). Post-hoc pairwise comparisons showed significantly greater expression of α-SMA in the painful group than in the control group (p<0.001) and the non-painful group (p<0.001), whereas there was no statistical difference between the control group and the non-painful group (p=0.470) (Table 3).
Painful group versus control group p<0.001; painful group versus non-painful group p<0.001; control group versus non-painful group p=0.470.
VAS, visual analogue scale.
Pearson correlation analysis demonstrated a significant correlation between VAS scores and the expression intensity of α-SMA, which was expressed as 1/gray value of α-SMA, (R=0.831; p<0.001). Linear regression indicated that the expression intensity of α-SMA was positively related to the scale of VAS (R 2=0.691, p<0.001) (Fig. 6).
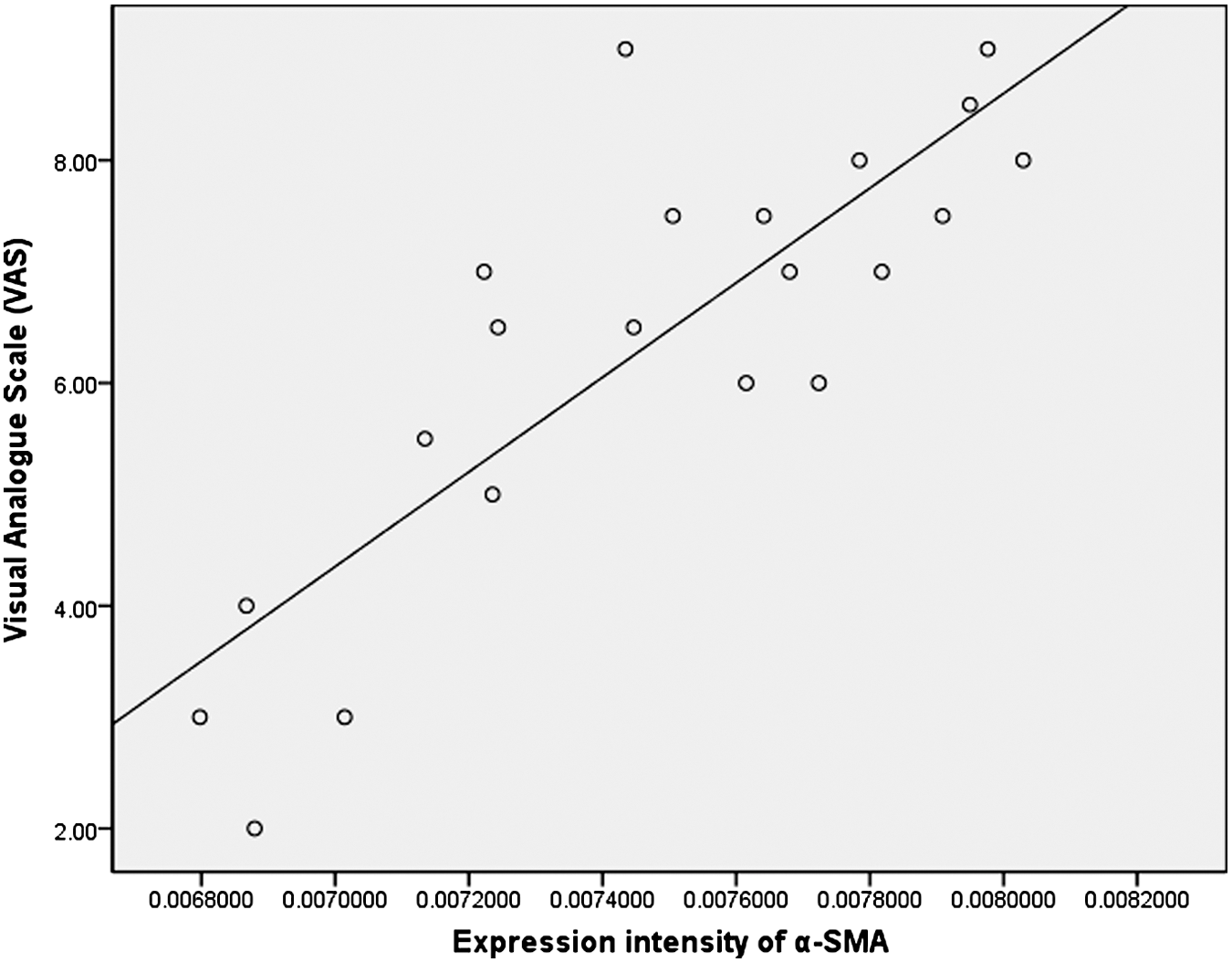
The scatter diagram demonstrates a linear correlation between the expression intensity of alpha smooth muscle actin (α-SMA) and the visual analogue scale (VAS) scores (R 2=0.691, p=000).
Discussion
The painful neuroma is a debilitating consequence of peripheral nerve injury. 7 There is no consensus on the optimal management of painful neuromas, with up to 150 various recommendations cited to control neuroma formation. 8,15 –20 This is likely attributed a lack of understanding of the pathophysiology of persistent neuroma-associated pain. 2,21 –23 ).
Despite the lack of consensus on the optimal clinical management of painful neuromas, many authors have reported success after freeing the traumatic neuroma from surrounding scar tissues and transferring it into surrounding muscle, vein, or bone. 17 –20 The rationale for this treatment is based on the belief that external mechanical compression of the neuroma from surrounding the scar tissues leads to persistent neuroma-associated pain. The lack of consistent results from this technique 8 suggests that additional sources of neuroma-associated pain exist.
We hypothesize that another source of compression within the neuroma may exist, and that the myofibroblast activity may be responsible. The myofibroblast has been widely accepted to actively promote dermal wound contraction 24 and possibly prevent axon elongation in the process of peripheral nerve regeneration along a tubulated gap. 25 Its activity is beneficial for dermal wound closure and for restoring the mechanical stability of injured organs against rupture. However, deregulated and chronic myofibroblast activity generates tissue deformation by contracture and impedes organ function. 26 In addition to being a marker of the myofibroblastic phenotype, 9 α -SMA contributes to the additional generation of contractile forces in non-muscle cells 27 and can upregulate the contractile activity of fibroblasts. 28 This may partially explain the clinical finding of positive correlation between increasing α-SMA expression and VAS pain scores.
In the present study, we found that persistent high expression of α-SMA was observed in the painful group in comparison with only slight expression in the non-painful group and no obvious expression in the normal nerve tissue. Based on these findings, we speculated that the contractile myofibroblast may entrap the regenerated small nerve fibers within the neuroma, leading to persistent mechanical stimulation of the regenerated nerve fibers and the clinical presentation of spontaneous neuroma-associated pain. Our hypothesis that internal compression contributes to neuroma-associated pain may be in agreement with the previously described “micro”- double crush syndrome. 29
Studies have demonstrated that exogenously applied and endogenously generated mechanical tension can regulate α-SMA expression, and its expression responds to the mechanical tension positively. 30 Clinically, the mechanism of the widely adopted approach for the treatment of painful neuroma with transferring the nerve ends into muscles, veins, or bones can also be partially explained in this perspective. It is conceivable that by capping the nerve ends with these methods and away from the scar tissues, the external mechanical stimuli are reduced, and, accordingly, the expression of α-SMA in the neuroma will be terminated and decreased, contributing to the relief of pain and discomfort. It is reported that smooth muscle cell contraction is Ca2+ dependent and is reversible physically, whereas tension production by the myofibroblast is not reversible pathologically, leading to persistent expression of α-SMA and is regulated by a ρ/ρ kinase-mediated inhibition of myosin phosphatase. 31,32 Based on these facts, a new approach for the treatment of painful neuroma may be developed by gene therapy to inhibit the proliferation of myofibroblasts and the expression of α-SMA in traumatic neuromas.
In physiological remodeling, such as during dermal wound healing, the contractile activity of myofibroblasts is terminated when the tissue is repaired, and the α-SMA expression then decreases and myofibroblasts disappear by apoptosis. 33 In pathological wound healing, however, myofibroblast activity persists and leads to tissue deformation, which is particularly evident in hypertrophic scars. 12 The exact reason why the α-SMA expression is persistently present in the painful group is unclear. Myofibroblasts are of great interest for the study of mechanobiology. 26 It is mechanically active, contributing to alterations in overall tissue mechanics; most importantly, it is mechanosensitive and mechanically inducible. In addition, α-SMA is also suggested as not only a mediator of traction forces in contractile myofibroblasts, but also as a component of the global mechanotransduction system. 26 Hence, it is conceivable that the persistent expression of α-SMA may be a result of the persistent local mechanical stresses, which are produced by friction from adjacent structures, compression from surrounding scar tissues, and irritation from topographically rough elements in the amputated stump. The expression intensity may be correlated to the magnitude of mechanical stimuli, indicating that the expression level of α-SMA may be a corresponding response to the topical mechanical stimuli. Therefore, the linear correlation between the expression intensity of α-SMA and pain scale of VAS value can also be interpreted as a linear correlation between the local mechanical stimuli and the pain scale of VAS value. From this perspective, the existence of α-SMA may be simply interpreted as a result of external mechanical stimuli, not as a cause of neuromatous pain.
Histological examination is important for understanding the pathophysiology of traumatic neuromas, especially the status of the regenerated myelinated and unmyelinated fibers and the fiberosis situation. Previous investigations have been widely conducted using transmission electron microscope (TEM) and other. 34,35 In view of the main purpose of our investigation, the quantitative analyses regarding these parameters were not conducted, but require further study.
Conclusion
In summary, the expression of α-SMA may contribute to neuroma-associated pain either as a direct cause of pain or an indirect marker of existence of local mechanical stimuli. The presence of α-SMA in the painful group may be partially taken as the rationale for the transpositional procedures performed to “bury” a traumatic painful neuroma in surrounding muscle, vein, or bone. The attempts to inhibit its expression may benefit patients with intractable neuromatous pain, and provide insight into the development of new therapeutic strategy. The exact role of α-SMA in the painful neuroma and exact mechanism of neuromatous pain are both subjects for further investigation. It is unlikely that traumatic neuroma pain is generated and maintained by one mechanism alone. It is more likely a product of multiple factors.
Footnotes
Acknowledgments
We are indebted to Dr. Christopher J. Dy for his suggestions and revisions of our manuscript. We are also grateful to the assistant researcher Xiaolei Zhang for his technical instruction and literature retrieval. This study was supported by funding from the National Natural Science Foundation of China (81171477) and the Department of Education of Zhejiang Province (20051185).
Author Disclosure Statement
No competing financial interests exist.