
Abstract
More than 5,000,000 survivors of traumatic brain injury (TBI) live with persistent cognitive deficits, some of which likely derive from hippocampal dysfunction. Oscillatory activity in the hippocampus is critical for normal learning and memory functions, and can be modulated using deep brain stimulation techniques. In this pre-clinical study, we demonstrate that lateral fluid percussion TBI results in the attenuation of hippocampal theta oscillations in the first 6 days after injury, which correlate with deficits in the Barnes maze spatial working memory task. Theta band stimulation of the medial septal nucleus (MSN) results in a transient increase in hippocampal theta activity, and when delivered 1 min prior to training in the Barnes maze, it significantly improves spatial working memory. These results suggest that MSN theta stimulation may be an effective neuromodulatory technique for treatment of persistent learning and memory deficits after TBI.
Introduction
Hippocampal oscillations in the theta frequency range (5–12 Hz) play a critical role in facilitating short-term and episodic memory tasks, 7 spatial working memory and navigation, 8 –10 and arousal and sensorimotor processing. 11,12 Theta oscillations in the hippocampus are generated and maintained through connections with the medial septal nucleus (MSN). 13 Previous studies have demonstrated that hippocampal theta activity can be driven via electrical stimulation of the MSN. 14 Tetracaine inactivation of the MSN results in loss of hippocampal theta activity concomitant with learning and memory deficits in adult rats, and rhythmic electrical stimulation of septal projections restores theta oscillations and dramatically improves cognitive performance. 9 Increased theta power is associated with successful encoding of episodic memory in humans, 15 and recent data demonstrate an increase in hippocampal theta activity prior to successful memory retrieval. 16 Similar findings in animal studies have shown that theta rhythms correlate with memory storage 17 and behavioral learning. 18 These findings underscore the importance of theta activity in normal memory function.
Lateral fluid percussion TBI in rats leads to a chronic decrease in hippocampal theta activity that correlates with significant spatial navigation deficits. 19 Additional research has demonstrated that TBI can disrupt the organized firing patterns in the hippocampus necessary for working memory. 20 Given the abundant evidence for the role of theta oscillations in memory, we hypothesized that theta stimulation of the medial septal input to the hippocampus would increase hippocampal theta activity in rats after experimentally induced TBI. We further hypothesized that theta stimulation would improve functional performance in the Barnes maze, a spatial learning and memory task. The results of our study support both hypotheses, and provide new evidence for the therapeutic potential of DBS to restore cognitive performance following TBI via activation of hippocampal rhythmicity.
Methods
Fifty-six adult male Sprague–Dawley rats (Harlan, Indianapolis, IN; 300–350 g) underwent a lateral fluid percussion TBI (n=30) or sham-TBI (n=26). Animals were housed under a 12 h light and dark cycle and had continuous access to standard rat chow and water. All procedures conformed to the National Institutes of Health guidelines and were approved by the University of California, Davis Institutional Animal Care and Use Committee.
Lateral fluid percussion injury and electrode placement
Animals were anesthetized using 4% isoflurane in O2/N2O (1:2) carrier gas, and were then intubated and mechanically ventilated with 2% isoflurane to maintain surgical anesthesia. A circular 4.8-mm diameter craniectomy was made in the skull over the right hemisphere (leaving the dura intact) at a position of 3 mm lateral to the midline and midway between lambda and bregma (AP ∼−4.5mm). 21,22 A rigid plastic injury tube was placed into the craniectomy site, cemented to the skull using methyl methacrylate and cyanoacrylate adhesive, and filled with sterile saline. Then a calibrated ∼0.10 mL pulse of sterile saline was rapidly injected (18 msec) through the injury tube onto the surface of the dura with a pressure of 2.12–2.15 atm which reliably produces a moderate TBI in rats. 21 After TBI, the intubated rat was then returned to the ventilator with isoflurane at ∼1% for maintenance during surgical placement of a twisted bipolar electrode spanning the CA3-CA1 dorsal hippocampal fissure (AP −3.3 mm from bregma, lateral +2.0 mm from midline, 3.6 mm deep to skull; PlasticsOne MS333/3-B/SP, Roanoke, VA). An additional 2 mm craniotomy (−0.5 mm AP, 1.0 mm L) was made to allow access for implantation of a chronic, indwelling twisted parallel bipolar stimulating electrode into the MSN (−6.8 mm depth from the skull surface at 12.8 degrees to avoid the dural sinus; PlasticsOne MS303/3-B/SP). The locations of the MSN stimulating electrodes are shown in Figure 1a and b, and location of electrodes in the hippocampus used for recording theta activity are shown in Figure 1c and d.
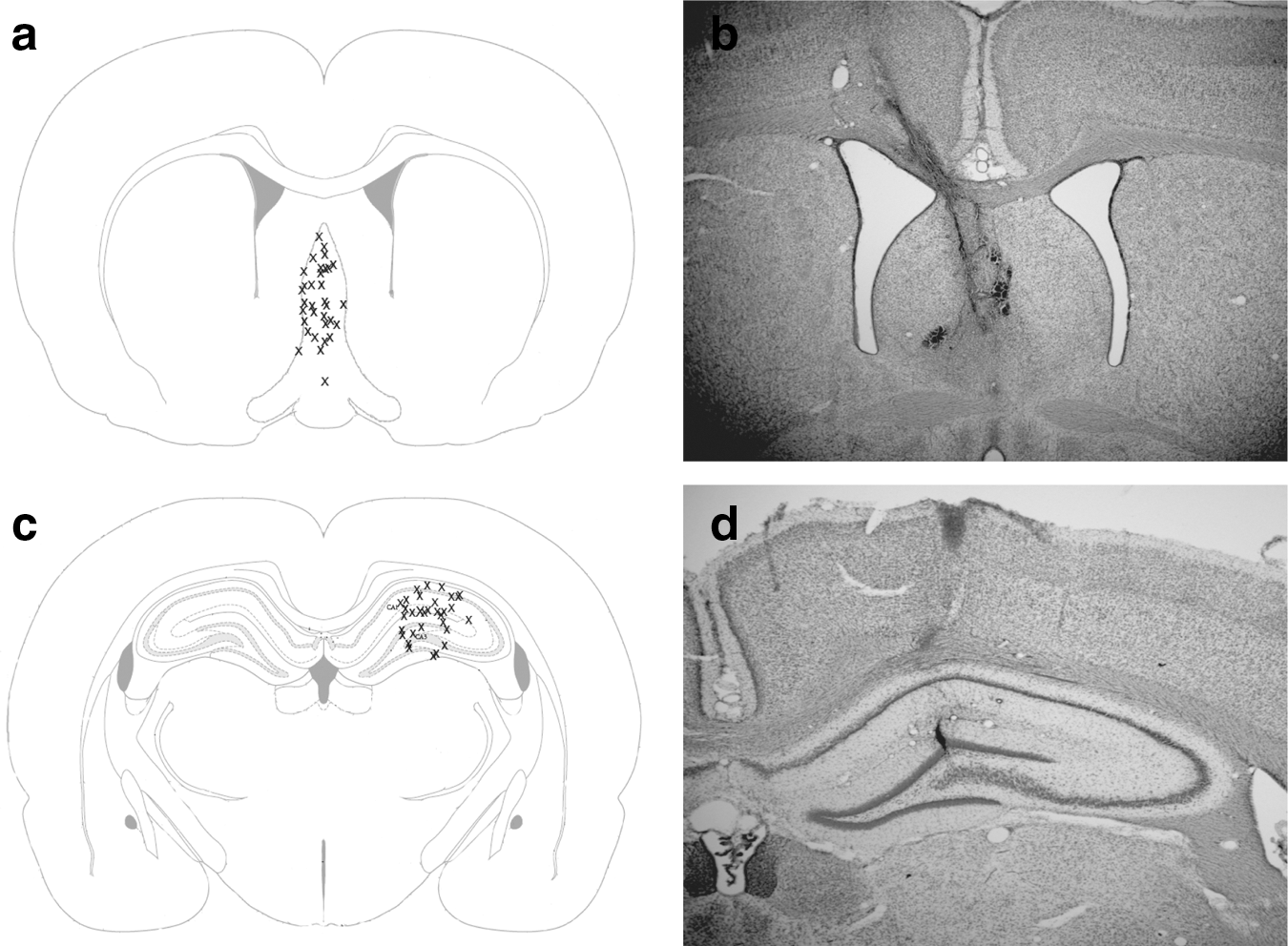
Diagram showing histologically verified placement of
Baseline electroencephalogram (EEG) recordings
On post-TBI days (PID) 1–6, each rat was placed in a video-EEG observation box (enclosed translucent Plexiglas cage, 25×45×50 cm) for 10 min. EEG was recorded with a preamplifier (Grass model 7P5B, Quincy, MA) and polygraph driver amplifier (Grass model 7DAG) and data collected with a computer data acquisition system (Polyview16 v1.1, Astromed, Inc., West Warwick, RI). A fast Fourier transform analysis was used to calculate the daily power spectral frequency analysis within the theta range (6–10Hz). 19 Differences in theta power at each minute post-stimulation were analyzed using a two-tailed Student's t test.
MSN stimulation paradigm
A twisted wire cable connected the implanted stimulating electrode to an isolated pulse stimulator (A-M Systems, model 2100, Sequim, WA) and electrical stimulation of the MSN was delivered using the following stimulation parameters: 7.7Hz frequency, 1-millisecond pulse width and current of 80μA. This current intensity was based on preliminary experiments with several intensities that established 80μA as an effective intensity for improving memory (data not shown).
Effects of stimulation on endogenous theta
A separate group of animals (sham, n=5; TBI, n=5) were examined for the effects of theta stimulation on endogenous theta power. Implanted MSN electrodes were attached to the pulse stimulator device, and the rats were placed in the video-EEG observation box. Video and EEG recordings from the hippocampus were simultaneously collected for 15 min (baseline), during 15 min of MSN theta frequency stimulation (7.7 Hz) and for 15 min after cessation of stimulation on post-injury days 1–6. The overall changes in post-stimulation theta power were determined using a repeated measures ANOVA (2 groups×9 time points) with Bonferroni post-hoc analyses.
Barnes maze
The Barnes Maze was performed as previously described. 19 The Barnes maze consists of a white Plexiglas circular table with a diameter of 122 cm. There are 18 holes (7.5 cm diameter) spaced equally around the periphery of the table with a dark escape box under one of the holes. White curtains surround the table with four distinct visual cues spaced equally around the curtains. On PID 4, 1 day prior to testing in the Barnes maze, each animal was connected to the EEG recording system and placed in the escape box under ambient lighting and allowed 5 min to habituate to the conditions. On PID 5, the table was rotated 135 degrees from the initial position used for habituation and two bright overhead LED lights (500 W each) were illuminated during testing. Animals were randomly assigned to receive either stimulation (sham, n=11 TBI, n=8) or no stimulation (sham, n=14; TBI, n=13) and were tested twice daily with video-EEG recordings. Prior to each trial, the rat was connected to the EEG recording device and stimulating device. For each trial, the animal was first placed in a dark box at the center of the table. While in the box, animals received either 1 min of theta stimulation or no stimulation. After 1 min, stimulation was stopped and the box was raised with an overhead drawstring from outside of the curtain. Animals were allowed 5 min to find the escape hole and enter the box below. If they did not find the box they were manually guided to the escape box. After each trial, animals were kept in the escape box for 1 min prior to being placed in a cage for a 5 min inter-trial interval (after the 4th min the animals were placed back in the dark box at the center of the table and, for the stimulation group, theta stimulation reinstated for 1 min, so that the second trial would start precisely at 5 min). Dependent variables included latency to find the escape box, an analysis of search strategy pattern, and locomotor speed during each trial. Locomotor speed (distance traveled/escape latency in seconds) was calculated by determining path length during each Barnes maze trial. Data for the Barnes maze were analyzed using a 4×3 ANOVA with a Bonferroni post-hoc analysis, with escape latency over the 3 trial days used as the repeated measure within groups variable.
Search strategies were categorized as spatial, peripheral, or random. 23 The spatial search strategy was operationally defined as the rodent using direct pathways toward the escape hole, indicating the use of a spatial working memory strategy. The peripheral search strategy was operationally defined as circling the edge of the platform prior to finding the escape hole, implying the use of a non-spatial strategy to solve the task. The random search strategy was operationally defined as searching non-consecutive holes prior to the escape hole without a discernible search pattern. Differences in search strategy patterns were analyzed using a χ2 analysis collapsing data across trials.
Histology
Rats were euthanized 8–14 days after TBI by anesthesia (Euthasol, Virbac Corp., Fort Worth, TX) and transcardially perfused with 100 mL of 0.1M sodium phosphate buffer saline (PBS, pH-7.4) followed by 50 mL of 4% paraformaldehyde (pH 7.4). Brains were extracted and stored overnight in 4% paraformaldehyde at 4°C. Brains were transferred first to 10% sucrose for 4–6 h and then cryoprotected in 30% sucrose for a minimum of 24 h. Brains were then rapidly frozen in dry ice and stored at −80°C. Serial coronal sections were cut at 45 μm thickness with a microtome (model #869; American Optical Corporation, Buffalo, NY) starting at +1.0 mm bregma and ending at −4.80 mm bregma in order to capture all tissue spanning between the MSN and the caudal aspects of the hippocampus. Tissue was saved in 24-well cell culture plates for histological staining. Sections in the vicinity of electrodes were mounted onto 0.1% gelatin-subbed slides and stained with cresyl violet to identify precise electrode placement.
Results
Hippocampal theta power is attenuated after TBI
Spontaneous hippocampal oscillations recorded during the 10 min spent in the Plexiglas observation box demonstrated that lateral fluid percussion TBI significantly decreased theta power in the ipsilateral hippocampus as compared with sham control animals during the first 6 days after TBI (TBI: 0.952±0.062 root mean squared voltage [VRMS] and sham: 1.572±0.128 VRMS, respectively, F (1,30)=15.51, p<0.05; Fig. 2a). Similarly, log theta power was attenuated in TBI rats relative to sham rats (t[10]=12.55, p<0.001; Fig. 2b). Consistent with previous studies, hippocampal oscillations were predominantly in the theta frequency range in both sham and TBI rats (Fig. 2c and d).
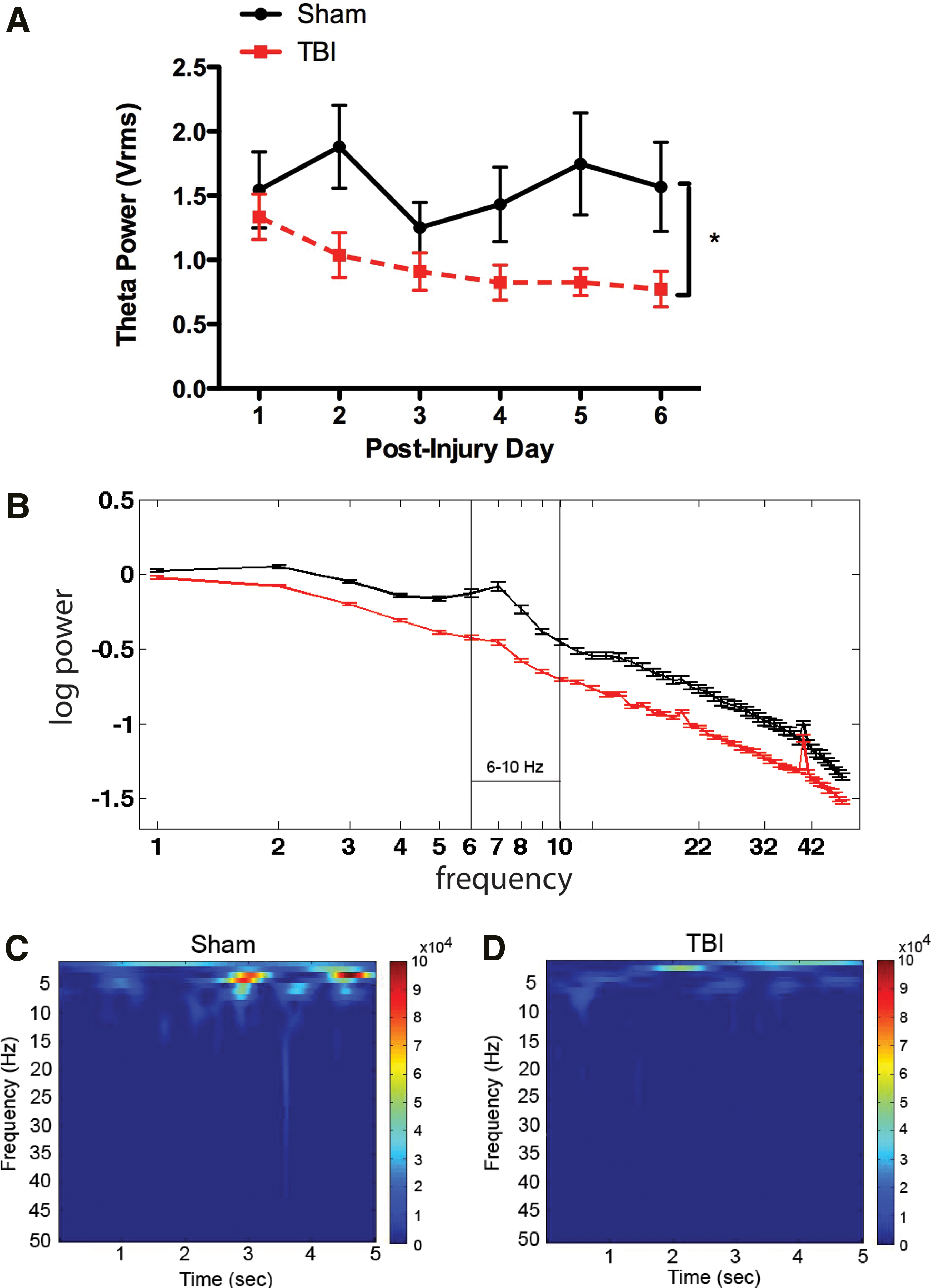
Effects of traumatic brain injury (TBI) on hippocampal theta activity.
Hippocampal theta power is increased during MSN stimulation
Theta power was increased above baseline levels during MSN stimulation in both TBI and sham animals (197.4±24.4% [n=5] and 218.3±18.4% of baseline [n=5], respectively; Fig. 3a and b). Hippocampal theta power was stably elevated throughout the duration of stimulation and did not markedly fluctuate in magnitude.
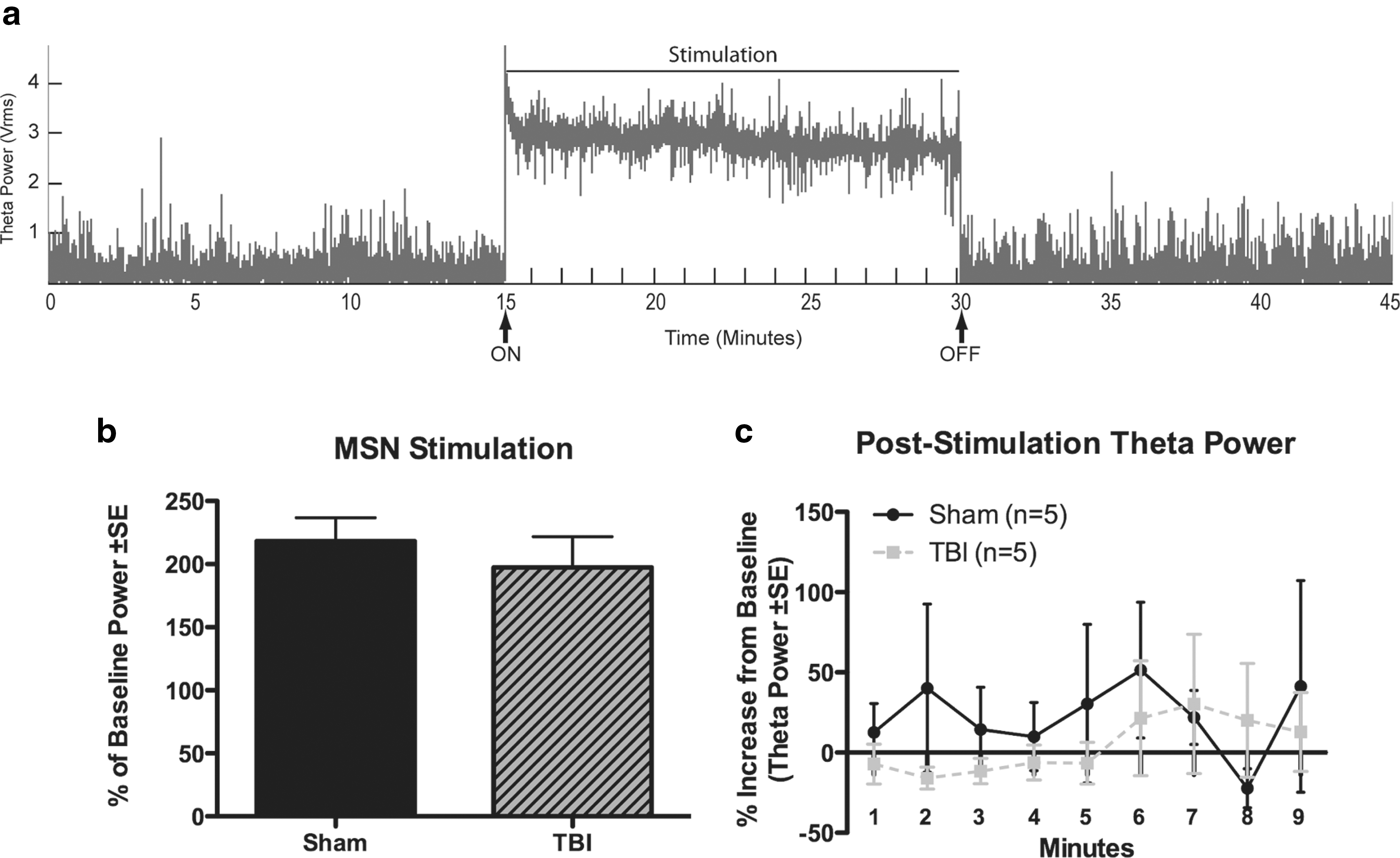
Effects of medial septal nucleus stimulation on hippocampal theta power.
To determine if stimulation had persisting effects, EEG was recorded for 15 min following the termination of stimulation. There were no persisting effects of stimulation on theta power (Fig. 3a and c). Specifically, there were no significant differences in theta power between baseline recordings and the first 15 min after termination of stimulation for TBI or sham animals (F (3,14)=0.35, p=0.78), or when each time point was analyzed separately by two-tailed t tests (Fig. 3c). No evidence of epileptic after-discharges, convulsive seizures, or stereotypical behaviors was noted during or after MSN stimulation.
MSN stimulation improves spatial working memory following TBI
There was a significant difference in escape latency among treatment groups on the Barnes maze (F (2,39)=4.554, p<0.05). Specifically, as shown in Figure 4a, TBI rats had significantly longer escape latencies across trials on PIDs 5–7 than did sham controls (p<0.05). In contrast, TBI rats receiving MSN theta stimulation had significantly shorter escape latencies than did TBI rats without stimulation (p=0.05). There was no significant difference between TBI stimulated and sham rats. Stimulation of sham rats did not alter average escape latency compared to unstimulated sham-injured rats (F (1,21)=1.127, p=0.30; Fig. 4b). Additionally, there was no difference in locomotor speed during the Barnes maze among the four groups (p=0.46).
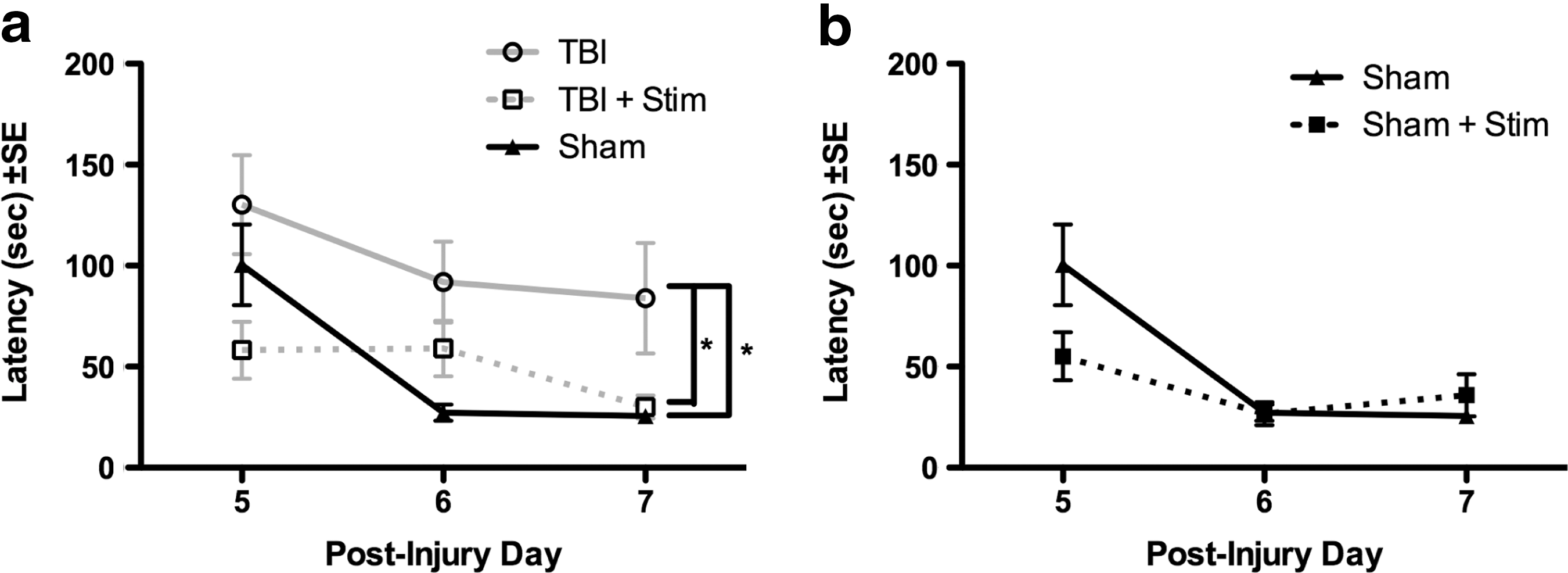
Barnes maze performance.
Overall distribution of search strategies
There were significant differences in search strategy patterns used by the treatment groups in finding the Barnes maze escape hole (Fig. 5a–d). Specifically, search patterns for the TBI group were significantly different from both the sham group (χ2[2]=6.17, p<0.05; Fig. 5b vs. 5a) and the TBI + Stim group (χ2[2]=11.37, p<0.01; Fig. 5b vs. 5d). Importantly, after theta stimulation, the search strategy for the TBI + Stim group did not differ from the sham group (χ2[2]=2.737, p=0.25; Fig. 5d vs. 5a). The Sham + Stim did not differ significantly from the sham group (χ2[2]=1.335, p=0.51; Fig. 5a vs. 5c).
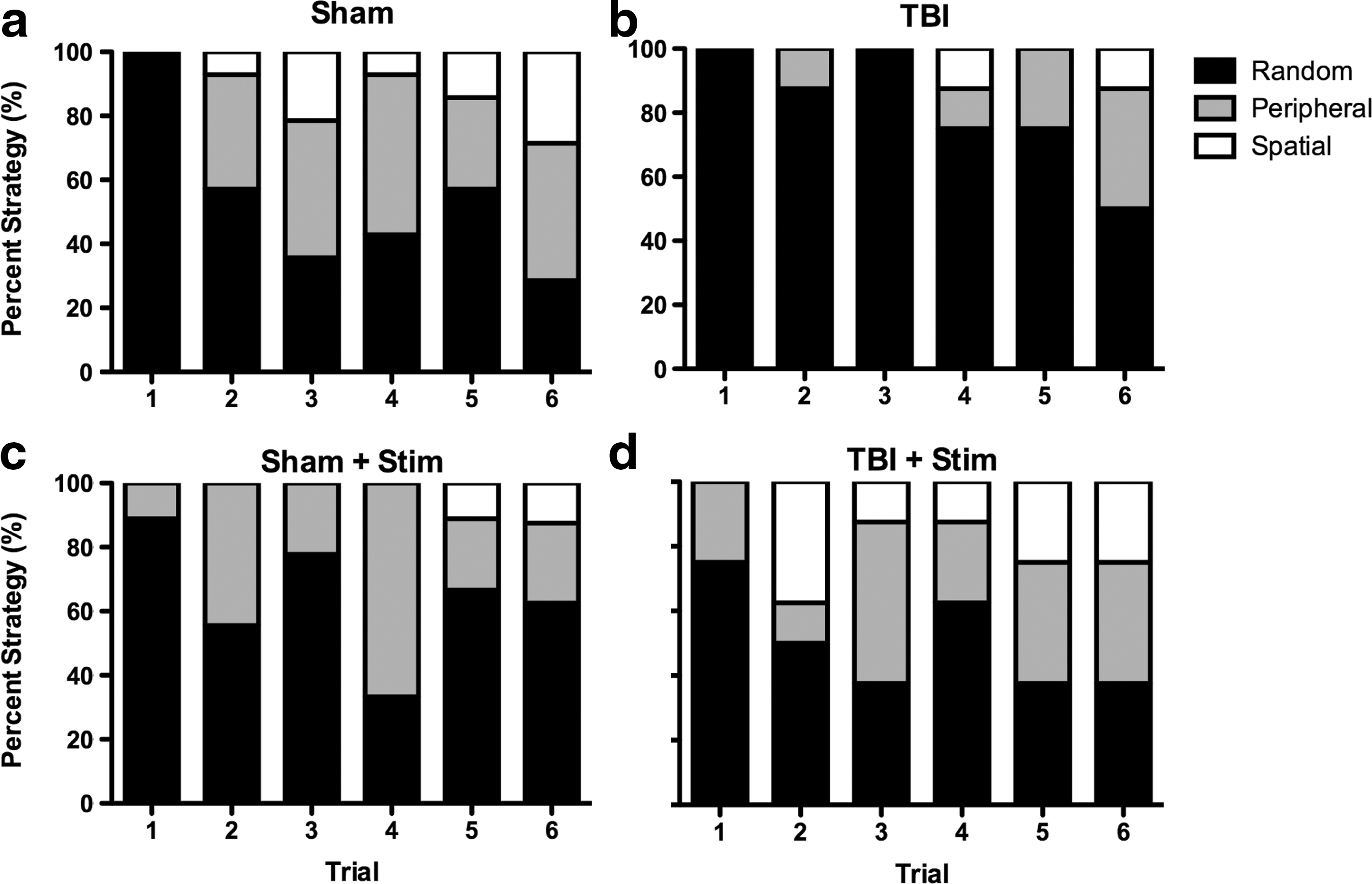
Percent time using spatial, peripheral, or random search strategies averaged across trials.
Random search strategy
As shown in Figure 5, on the first trial, all groups essentially used a random search strategy. However, across trials, the majority of rats in the sham group no longer used this strategy (Fig. 5a), whereas TBI rats continued to use a random search strategy (Fig. 5b). Importantly, the use of a random search strategy for the TBI+Stim group (Fig. 5d) was similar to that of the sham group (Fig. 5a) and the Sham+Stim group (Fig. 5c). Group differences in search strategies were also apparent when expressed as the percent of all trials in which the random strategy was used. Viewed this way, the sham group used a random strategy on 61.1% of trials, the Sham+Stim group used a random strategy on 66% of trials, and the TBI+Stim group used a random strategy on 52.1% of trials, whereas the TBI group used the random strategy on 83.3% of trials.
Peripheral search strategy
Use of the peripheral search strategy across trials was qualitatively similar for the sham (29.6%), Sham + Stim (30.2%) and TBI + Stim (27.1%) groups, whereas rats in the TBI group showed markedly less use of this strategy over trials (15.2%).
Spatial search strategy
No rats in any group showed a spatial search strategy on the first trial. However, across trials, the sham group used a spatial strategy in 9.3% of trials, whereas the TBI rats group exhibited a spatial strategy in only 4.2% of trials (Fig. 5a vs. 5b). Interestingly, the rats in the TBI+Stim group used a spatial strategy on 20.8% of trials, and the Sham + Stim group used a spatial strategy on 3.7% of trials.
Discussion
This study utilized the rat lateral fluid percussion injury model to study the effects of TBI on hippocampal theta oscillations, as well as the effects of MSN theta stimulation on spatial working memory. We report two novel findings: 1) fluid percussion injury resulted in depression in hippocampal theta in the first 6 days following injury, and 2) MSN theta frequency stimulation delivered immediately before training trials in the Barnes maze significantly improved spatial working memory. Importantly, the benefits of pre-stimulation outlasted the period of actual theta stimulation. These findings illustrate for the first time that hippocampal-dependent spatial working memory can be improved by theta stimulation of the medial septal input.
Hippocampal theta activity is reduced acutely after lateral fluid percussion injury
Previous research from our laboratory demonstrated that hippocampal theta frequency is chronically depressed 6–8 weeks after TBI. 19 In the present study, a reduction in hippocampal theta power and log theta power were similarly observed during EEG recording within an enclosed Plexiglas observation chamber during the first 6 days after injury. One explanation for decreased hippocampal theta activity may be damage to the generator of this oscillatory rhythm, the MSN. 24,25 Another possibility is that hippocampal cell death and dysfunction 26,27 underlies the decrease in theta activity, compromising ability of the hippocampal circuitry to receive or maintain oscillatory activity. It is also possible that damage to other brain regions that interact with the hippocampus and are known to demonstrate theta band oscillations, such as the thalamus, 28 supramamillary/mammillary bodies, 29 and/or cingulate cortex, 30 may play a role in attenuating theta. Whereas previous research demonstrates that TBI results in hippocampal cell loss, 26,27,31,32 the reduction in theta activity may also be caused by other physiological perturbations, such as a reduction in neurotransmitter levels 33,34 or receptor expression, integrity of axonal connections, or robustness of hippocampal circuitry. 35 Although the underlying pathophysiology is not known, the current study demonstrates that TBI results in a substantial attenuation of hippocampal theta activity that is likely to contribute to deficits in working memory tasks.
Experimental stimulation of theta frequency results in benefits that outlast the stimulation period
Hippocampal theta activity has been correlated with exploration in both humans and rats. 12,36 Several lines of evidence support the theory that theta oscillations are necessary for effective navigation and spatial memory by generating a temporal organization of neuronal activity. 37,38 Additionally, retention and encoding of items are associated with increases in theta and gamma power. 15,39 –41 Our study demonstrates that MSN stimulation-induced increases in hippocampal theta power do not persist beyond the period of stimulation; however, this treatment paradigm still results in significant improvements in the Barnes maze test of learning and memory.
One possible explanation for this observation is that MSN stimulation synchronizes activity across a circuit that includes the hippocampus, and, therefore, may act as a priming mechanism for spatial working memory tasks. This hypothesis suggests that memory retrieval may be based upon “phase coding” through theta activity. 42 –44 This may explain why irregular stimulation of hippocampal theta, rather than fixed rhythmic stimulation, does not restore learning and memory deficits. 9 Previous research also demonstrates that theta activity immediately preceding training in a spatial memory task is crucial for successful recall in both rodent models 10 and in human epilepsy patients. 16 Specifically, Addante et al. demonstrated that increased theta activity prior to a task or event is correlated with successful episodic memory retrieval, whereas decreased theta activity is associated with poor episodic memory retrieval. These findings in human subjects are consistent with our finding that stimulation prior to engaging in a spatial learning task improves performance, possibly by enhancing memory encoding.
Improved performance on the Barnes maze is the result of enhanced spatial working memory
Data presented here suggest that spatial working memory can be enhanced by stimulation of the MSN in the theta frequency band. It is important to note that hippocampal theta activity prior to the Barnes maze task resulted in improved spatial working memory, but that Sham+Stim animals did not benefit from stimulation. Rats in all groups traveled at the same speed (distance traveled/escape latency) during the Barnes maze task. These data suggest that the benefits of MSN stimulation are not due to a nonspecific, hyperarousal effect. Moreover, theta stimulation of TBI rats also altered the spatial search strategy so that it appeared to be more similar to that of sham rats than to that of TBI rats, suggesting that enhanced spatial learning is produced by MSN theta stimulation.
In the Barnes maze, all treatment groups initially used the random search strategy. However, across trials, animals in the sham group progressively utilized more “peripheral” or “spatial” search strategies, whereas TBI rats did not show this change in strategy. A critical finding of this study was that MSN stimulation significantly increased the use of non-random strategies in animals that had experienced TBI (i.e., peripheral and spatial). This suggests that not only does electrical stimulation of the MSN enhance theta activity, but that it also effectively modulates a medial septal–hippocampal connection that in turn improves more complex aspects of cognitive performance. Hippocampal theta activity is coupled to gamma “bursts” in various brain regions that may function in a concerted effort to guide performance. 45 As theta–gamma coupling plays a critical role in temporal segmentation and memory, it is possible that MSN stimulation restores synchrony or coupling of brain networks that are necessary for normal spatial working memory activities.
MSN stimulation in sham rats did not improve spatial working memory in the Barnes maze. The data suggest that the sham-stimulation group may have performed worse than the sham without stimulation group. Whereas sham rats, with or without stimulation, did not differ in Barnes maze latency throughout the six trials, Sham animals without stimulation used the optimal spatial strategy more often (Fig. 5a and c). This further suggests the importance of restoring normal activity in the medial septal–hippocampal circuitry after brain injury via theta frequency stimulation, rather than applying a nonspecific electrical input to the circuit. Stimulating an intact circuit, on the other hand, may disrupt normal spatial–temporal firing of hippocampal neurons in a way that may disrupt spatial working memory. Alternatively, the lack of benefit from stimulation of a sham rat may be caused by stimulation at a suboptimal frequency.
Potential mechanisms for cognitive improvement after MSN stimulation
One potential explanation for the beneficial effects of electrical stimulation on cognitive performance is that it decreases neuronal cell loss in the hippocampus. However, previous research has demonstrated that the majority of cell death occurs within the first 24 h after injury, a time when stimulation was not given in the present study. 32,46 In addition, memory impairment can occur after TBI even in the absence of hippocampal cell death. 47 Another possible mechanism is that stimulation is merely increasing metabolic cellular activity, resulting in increased neuronal activity and, thereby, improved performance. Animal and rodent studies have demonstrated a reduction in frontal lobe, temporal lobe, hippocampal, and thalamic metabolism following TBI. 48 –51 Whereas decreased brain metabolism may persist up to 3 months following lateral fluid percussion injury in rats, hypometabolism is most severe 2 days after injury. 52 As the stimulation paradigm used in this study begins after this time point, this explanation also seems unlikely. It is more likely that stimulating the MSN replaces the lost theta input, allowing the injured hippocampus to function more normally. Further studies are needed to determine if stimulation in different frequency ranges, such as gamma or beta, or an irregular stimulation paradigm, such as theta burst stimulation, has similar effects on spatial working memory.
Clinical implications
DBS is an adjustable therapy that is highly effective in the treatment of a variety of neurological disorders, but its role in treatment of TBI has not been extensively explored. Efforts to treat disorders of consciousness following TBI were explored in the past, with limited benefit, 53,54 but a recent successful case report 55 has renewed interest in its role for treating coma. The persistent comatose state represents <1% of all patients surviving TBI, whereas the majority of severe TBI survivors have persistent cognitive deficits that dramatically impact their productivity and quality of life. 56 Therefore, targeting post-traumatic cognitive deficits rather than disorders of consciousness after TBI may have a significantly larger clinical impact. Recent clinical studies suggest that DBS may improve memory function in patients with Alzheimer's disease or epilepsy, 5,6 and in the present study we show that theta frequency-specific stimulation of the medial septal–hippocampal pathway reduces TBI-induced cognitive deficits. To date, there have been no successful therapeutic interventions to improve the quality of life for TBI patients with persistent cognitive deficits. These data demonstrate that neuromodulation in the form of DBS has the potential to improve hippocampal circuit function and cognitive performance following TBI, and, therefore, represents a novel and exciting potential treatment strategy for millions of patients with chronic disability.
Footnotes
Acknowledgments
This project was supported by the Bronte Epilepsy Research Foundation, and a “Vision Grant” from the UC Davis Health System National Board of Advisors. We thank Angela Echeverri, Mikhail Melnik, Rebecca Wong, and Heather McConnell for their expert technical assistance.
Author Disclosure Statement
No competing financial interests exist.