
Abstract
Spinal cord injury (SCI) is less common in children than in adults, but in children it is generally more severe. Spinal loading conditions (speed and displacement) are also thought to affect SCI severity, but the relationship between these parameters is not well understood. This study aimed to investigate the effects of vertebral speed and displacement on the severity of SCI in infants and adults using a rodent model of vertebral dislocation. Thoracolumbar vertebral dislocation was induced in anaesthetized infant rats (∼30 g, 13–15 days postnatal, n=40) and adult rats (∼250 g, n=57). The 12th thoracic vertebra was secured, whereas the first lumbar vertebra was dislocated laterally. Dislocation speed and magnitude were varied independently and scaled between adults and infants (Adults: 100–250mm/s, 4–10mm; Infants: 40–100mm/s, 1.6–4mm). At 5 h post-injury, rats were euthanized and spinal cords harvested. Spinal cord sections were stained to detect hemorrhage (hematoxylin and eosin) and axonal injury (β-amyloid precursor protein). For each millimeter increase in vertebral displacement, normalized hemorrhage volume increased by 1.9×10−3 mm3 (p=0.028) and normalized area of axonal injury increased by 2.2×10−1mm2 (p<0.001). Normalized hemorrhage volume was 3.3×10−3 mm3 greater for infants than for adults (p<0.001). Magnitude of dislocation was found to have a different effect on the normalized area of axonal injury in adults than in infants (p=0.003). Speed of dislocation was not found to have a significant effect on normalized hemorrhage volume (p=0.427) or normalized area of axonal injury (p=0.726) independent of displacement for the range of speeds tested. The findings of this study suggest that both age and amount of spinal motion are key factors in the severity of acute SCI.
Introduction
Spinal cord injury (SCI) is relatively rare, its incidence ranging globally from 11.5 to 57.8 per 1,000,000 cases per year. 1,2 Pediatric SCI is even rarer, accounting for ∼3% of new SCI cases. 3 Although rare, the consequences of SCI can be devastating and it can result in permanent disability and lifelong economic and social impacts. In 2008, the total cost of SCI in Australia was estimated to be $2.0 billion AUD. 4
In young children, the mechanism of spinal injury leading to cord injury commonly involves vertebral dislocation without fracture, or SCI without radiographic abnormality (SCIWORA). 5 SCI in adults, by contrast, commonly involves vertebral fracture, of which the most common mechanisms are vertebral fracture dislocation followed by burst fracture. 1 Two clinically relevant vertebral dislocation models have been developed, 6,7 and one has been modified for investigating pediatric SCI. 8 Contusion models for experimental SCI are valuable, but vertebral dislocation models offer a unique additional opportunity to study cord injury under loading conditions that more closely mimic the most common mechanisms of SCI in humans.
The mortality rate is higher following SCI in younger children (≤11 years of age) than in older children (>11 years of age), 5 and these injuries are more likely to be neurologically complete in children 0–5 years of age than in adolescents >16 years of age, and adults. 9 This pattern of injury, in which SCI is more severe in children than in adults, has also been confirmed experimentally under controlled conditions using a rodent model of vertebral dislocation. 8 Despite this, children tend to have higher rates of neurological improvement, 10 and this has also been demonstrated experimentally in rats by testing locomotor function following spinal cord contusion injury, 11 and transection injury. 12
The severity of SCI is also thought to depend upon the speed and displacement of spinal motion during injury. Recent studies of motor vehicle collisions found that impacts with fixed roadside objects were more likely to result in a serious rather than a minor spinal injury compared with car to car collisions. 13,14 This suggests that the crash pulse shape resulting from a fast deceleration is more likely to result in a severe injury, and that injury severity increases with increasing speed, because of the higher levels of kinetic energy that must be dispersed following a crash. Experimental studies have also found that, for the same displacement, higher velocity contusion produces greater white matter hemorrhage and axonal damage, 15 and that axonal strain is correlated with neuronal injury severity. 16 In addition, higher severity crashes, which result in a larger deflection of the spine, were found to be associated with a greater risk of severe spinal injury. 14 This indicates that injury severity also increases with increasing displacement. However, it is not known how these two important loading variables interact during the primary or secondary phases of injury; whether they affect adults and infants in different ways, and whether the application of loading to the spinal column, rather than directly to the cord, changes the effect of these variables.
The factors of age, speed, and displacement could interact with each other in determining the severity of SCI. In a study investigating spinal cord biomechanics using in vitro spinal cords of adult and infant rats, Clarke et al., found that strain rate (i.e., rate of deformation) had a greater effect on tissue stress (i.e., force acting over an area of an object) in adult spinal cords than in infant spinal cords. 17 This suggests that adult spinal cords may be more sensitive to changes in impact speed than infant spinal cords.
In summary, it is thought that age, speed, and displacement independently affect SCI severity; however, the relationship between these injury variables is not known, specifically whether the speed and displacement of injury affect adults and infants in different ways. Therefore, the current study aims to investigate which aspects of loading are the greatest risk factors for increased SCI severity in the acute phase of injury, and whether this is different in infants and adults.
Methods
Vertebral dislocation injury
All experiments were performed in accordance with a protocol approved by the University of New South Wales Animal Care and Ethics Committee. Infant Sprague–Dawley rats (∼30 g, 13–15 days postnatal, n=40) and adult male Sprague–Dawley rats (∼250 g, n=57) were anesthetized using an intraperitoneal injection of urethane (adults: 1.4 g/kg, infants: 2 g/kg; Sigma-Aldrich, St. Louis, MO). These ages were chosen to approximately correspond to a 1–2-year-old human child with an immature spine and cord, and an adult with a mature spine and cord.
Once the rats were under deep anesthesia, a dorsal approach was used to expose the laminae and transverse processes of the vertebral column from the level of the 12th thoracic vertebra (T12) to the 1st lumbar vertebra (L1). The supraspinous and interspinous ligaments between T12-T13 and T13-L1 were cut, and facetectomies were also performed at these levels. This ensured consistent realignment of the vertebrae following the dislocation in each case. The ribs of T12 were cut adjacent to the joint capsule to allow custom-made vertebral clamps to be fastened around the vertebral bodies of T12 and L1 to secure the rat into the injury device. The vertebral dislocation device used in this study was first described by Fiford et al., and consists of a stationary beam, a moving beam, and sensors for monitoring displacement and force (Fig. 1). 7 The device was used to hold T12 stationary while translating L1 laterally to the left at a desired speed and displacement. This position was held for 1 sec before L1 was returned to its original position at the same speed.
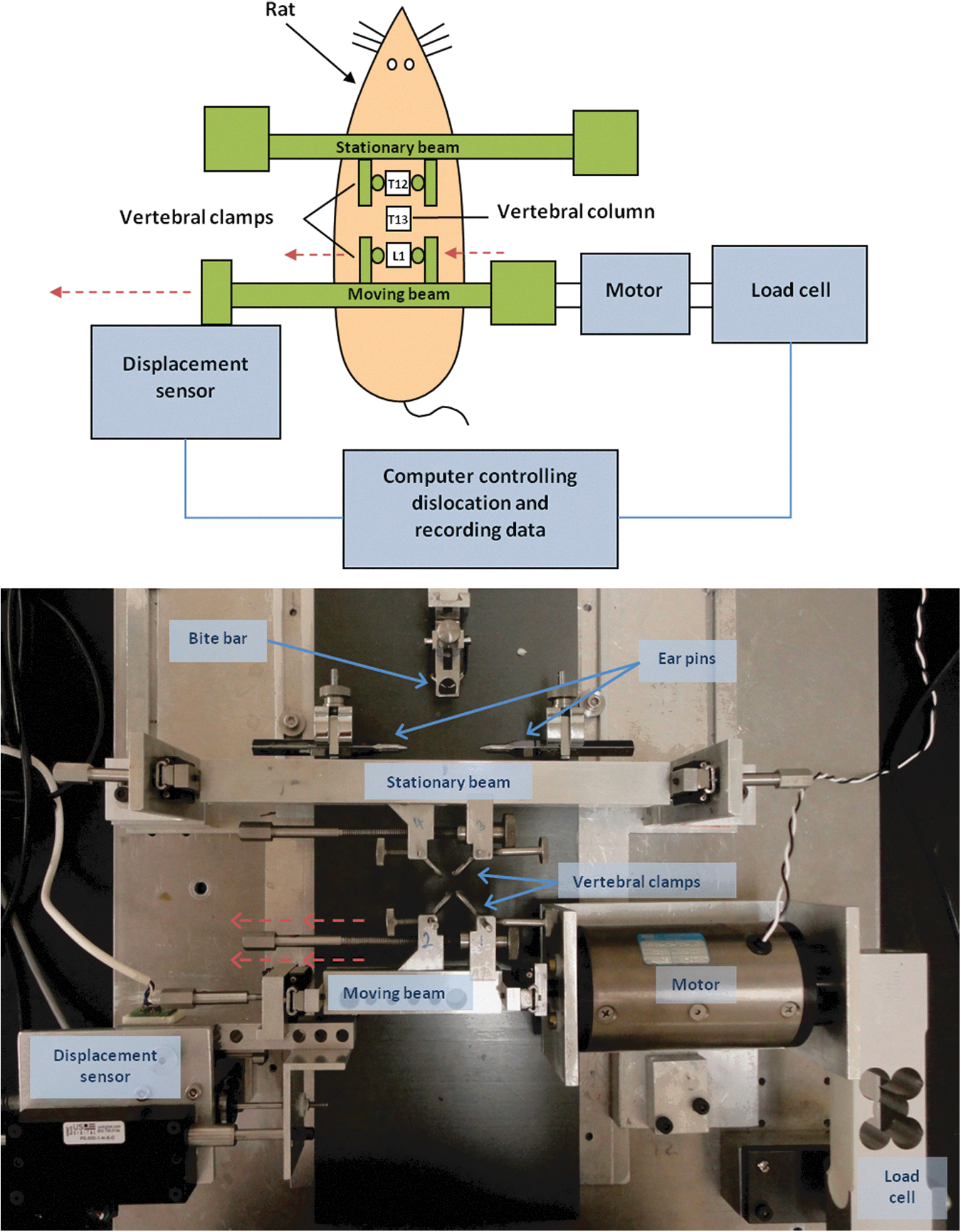
Simplified diagram (top) and photograph (bottom) of the custom-made lateral vertebral dislocation injury device. Vertebral clamps are attached to the vertebral bodies of T12 and L1, and the rat is secured in place using a stereotaxic frame. The computer controls translation of the caudal moving beam with attached vertebral clamps at a target speed and displacement via the motor. The L1 vertebra is translated laterally with the caudal clamp to create the injury. The displacement sensor and load cell record biomechanical data. Color image is available online at
Five different injury groups were studied (Table 1). These parameters were chosen to encompass a wide range of SCI severities and examine the effects of speed and displacement independently, while minimizing animal use. Injury groups 1–3 examined the effects of speed, whereas injury groups 3–5 examined the effects of displacement. In the adult cohort, eight rats were tested using injury conditions intermediate between the established groups, to aid statistical analysis. The injury conditions for the infant cohort were scaled by a factor of 2.5 based on the measured nerve root separation at T12-T13 (a nominal rostrocaudal geometrical measure) of adult and infant spinal cords, with the purpose of applying equivalent strain and strain rate to adult and infant cords during injury. 8 Biomechanical data were collected from the displacement sensor and load cell connected to the moving beam of the injury device.
Injury Conditions for Infant and Adult Cohorts
Following the injury, the vertebral clamps were detached from the rat and the spinal column was stabilized by adhering stainless steel rods (0.8 mm in diameter) to the laminae of T12-L1 using a tissue adhesive (Vetbond tissue adhesive; 3M, St. Paul, MN). This allowed careful handling of the animal following injury without applying additional loading to the spinal cord and inducing further damage. The wound was sealed with tissue adhesive and the rats monitored for 5 h post-injury. After this survival period, all animals were euthanized with an overdose of Euthal (Pentobarbitone Sodium, 200mg/kg, i.p.; Delvet, Seven Hills, NSW, Australia). Animals were then immediately injected with Heparin (0.1 mL, IC; Heparin Sodium Injection, Pfizer, NY) and 1% sodium nitrite solution (0.1 mL, IC) before intracardial perfusion through the ascending aorta with phosphate-buffered isotonic saline solution (PBS, pH 7.4) followed by a solution of 4% paraformaldehyde in phosphate buffer (pH 7.4).
Tissue preparation
The entire spinal cord was isolated, and the meninges and nerve roots were removed and post-fixed for 12 h in 4% paraformaldehyde solution. It was then stored in a 30% sucrose solution with 0.1% sodium azide for at least 24 h. Following removal of the spinal cord, examination of the vertebral bodies, end plates, intervertebral discs, and posterior longitudinal ligaments was performed using a surgical microscope and spinal manipulation (axial, dorsoventral, and lateral) to identify any damage.
Spinal cords were embedded in Tissue-Tek OCT Compound (Sakura Finetek USA Inc, CA) and flash-frozen on an aluminum chuck. The spinal cord was sectioned dorsal to ventral using a cryostat at −16°C in the coronal plane at a thickness of 20 μm, and the cut sections were mounted on gelatinized microscope slides.
Histology
Slides were stained with hematoxylin and eosin to detect hemorrhage and to inspect gross morphology. Consecutive segments of each section were imaged using a digital camera (Pixelink, PL_A662; Scitech, Melbourne, Australia) mounted on a light microscope with a 4x objective lens (Olympus BH-2, Tokyo, Japan). The Cavalieri method of unbiased stereology, 18 as described by Howard and Reed, 19 was used to estimate the volume of hemorrhage within each spinal cord. This involved a manual grid-crossing method, and has been used previously to estimate the volume of hemorrhage in experimental studies of SCI. 6,8 To enable comparisons between the adult and infant spinal cords with different dimensions, the estimated volume of hemorrhage was normalized by dividing the raw volume of hemorrhage by the calculated volume of an infant or adult spinal cord segment between nerve roots at T12-13. 8 This was approximated by an elliptical cylinder (π×spinal cord radius 2 ×nerve root separation).
Immunohistochemistry
Slides were immunostained for beta-amyloid precursor protein (β-APP) to detect axonal injury. All incubations were performed at room temperature unless otherwise stated, and a negative control with the primary antibody omitted was used to confirm specific staining in all staining runs.
Slides were washed in xylene, rehydrated through graded alcohol (100%, 100%, 95%, 70%) and water and incubated in phosphate-buffered saline with Triton X-100 (PBST, pH 7.4) for 10 min. Endogenous hydrogen peroxidase was removed with a 30 min incubation in PBST with 3% hydrogen peroxide. Slides were rinsed through three changes of PBST for 10 min, and incubated in normal goat serum (1:20, Sigma-Aldrich) for 30 min and then incubated in rabbit polyclonal antibody for β-APP (1:100, Invitrogen Australia Pty Ltd, Melbourne, Australia) at 4°C for 18 h and rinsed through three changes of PBST. This was followed by incubation in biotinylated goat anti-rabbit secondary antibody (1:200, Vector Laboratories, Burlington, CA) for 1 h, and rinsing in three changes of PBST. Antibodies were labeled by incubating in avidin-biotin peroxidase complex (VECTASTAIN Elite Standard ABC kit; Vector Laboratories) for 1 h, and slides were then rinsed sequentially in PBST, Tris-buffered saline (TBS) and Nickel-TBS (NiTBS) for 10 min each. A 3,3-diaminobenzidine stain was prepared by dissolving a 3,3-diaminobenzidine tetrahydrochloride pellet (10mg, Sigma-Aldrich) in 20 mL of NiTBS, filtering the solution and adding 3 μL of hydrogen peroxide. Each slide was stained with this filtrate for 3 min, and placed in PBST for 10 min to stop the reaction. Slides were then rinsed in water, dehydrated through alcohol (70%, 95%, 100%, 100%), cleared through xylene, and cover-slipped using DPX mounting media (BDH, Poole, UK).
Images were captured from the dorsal, ventral, left lateral, and right lateral funiculi of the spinal cord using a digital camera (Pixelink, PL_A662; Scitech) mounted on a light microscope with a 20x objective lens (Olympus BH-2). For each funiculus, five images were captured:±3 mm,±1 mm, and 0 mm rostral and caudal from the lesion epicenter for adults, and±1.2 mm,±0.4 mm and 0 mm rostral and caudal from the lesion epicenter for infants. Rostral-caudal distance for imaging was scaled by a factor of 2.5 for infants as with the injury conditions, to account for their smaller spinal cord geometry. 8 These distances were chosen to capture an objective sample of injured and uninjured axons, based on previous studies estimating axonal injury. 8,20 Images were captured from three spinal cord sections for each funiculus (spaced 140 μm apart). To estimate the amount of axonal injury from these images, ImageJ (National Institutes of Health, Bethesda, MD) was used to set an intensity threshold and determine the area of the image staining positive for β-APP. Regions of interest within these images were manually selected to minimize false positives and negatives. The area of axonal injury was normalized to density by dividing the raw area of β-APP staining by the calculated coronal area of an infant or adult spinal cord segment between nerve roots at T12-13 (spinal cord width×nerve root separation). 8
Statistical analysis
Normality was assessed using the Shapiro–Wilk test for normality and inspection of skewness and histograms. Speed, displacement, and normalized volume of hemorrhage were normally distributed. Data for the area of β-APP staining were not normally distributed and, therefore, required a square root transformation to permit use of parametric statistics. The normalized area of positive β-APP staining for each white matter funiculus was analyzed in combination to demonstrate overall trends and separately to demonstrate differences among funiculi. Primary statistical analysis was conducted using linear regression modeling with analysis of covariance (ANCOVA). For hemorrhage, normalized volume of hemorrhage was used as the dependent variable and speed, displacement, and age were used as the independent variables. Age was coded with a dummy variable of 0 for infants and 1 for adults. For total axonal injury, the sum of positive β-APP staining from all four white matter funiculi was calculated. This was normalized and used as the dependent variable with speed, displacement, age, and the age-displacement interaction as independent variables. The differences among white matter funiculi were analyzed using repeated measures analysis of variance (ANOVA). This compared the effects of age, speed, displacement, and rostro-caudal distance from the injury epicenter in the four white matter funiculi. The cross-sectional pattern of axonal injury within the spinal cord was investigated by comparing ratios among funiculi. That is, linear regression modeling with ANCOVA was used to examine the effects of age, speed, and displacement on the ratio of axonal injury between dorsal and lateral, and ventral and lateral funiculi. The value for the lateral funiculus was calculated by taking the average of right and left lateral funiculi. Overall, these statistical tests aimed to quantify the effects of different injury conditions on adults and infants. Relationships were considered significant where p<0.05.
Results
Spinal column and SCI
Vertebral dislocation injury was induced in 57 adult and 40 infant rats. In adults, the injury resulted in fracture of the vertebral column for all rats in injury groups 3, 4, and 5 (100 mm/s, 10mm; 200 mm/s, 10 mm; 250 mm/s, 10 mm). Fracture was evident in 1/6 for injury group 1 (100 mm/s, 4 mm) and 9/10 for injury group 2 (100 mm/s, 7 mm), giving a total of 51/57 with evident vertebral fracture after injury. This was always a transverse fracture through the caudal endplate of the T13 vertebra. All fractures were complete with separation of T13 from L1, thus compromising the spinal canal during loading. Returning the displaced vertebra to the original position after injury and prior to stabilization restored the canal acutely. The posterior longitudinal ligament at this level was also ruptured in all cases of fracture. In infants, the injury resulted in fracture only in 1/40 rats with a diagonal fracture through the growth plate of the vertebral body. All others had intact vertebral bodies, end plates, posterior longitudinal ligaments, and vertebral discs on macroscopic examination and manual manipulation of the thoracolumbar junction.
The injury tended to disrupt the spinal cord parenchyma in a diagonal pattern. Lesions consistently extended from the left rostral area to the right caudal area, centered on the T13 vertebral body, and formed an angle of ∼45 degrees to the direction of dislocation. The spatial distribution of injury, and the differences in lesion pattern between adult and infant animals was very similar to that described in a previous study using this model. 8 Sham-operated controls (infant and adult rats that underwent the same surgical exposure and placement in the device but without application of dislocation), have been performed previously using the same custom-made injury device, and no spinal cord pathology was found. 8
Hemorrhage
The amount of hemorrhage evident in sections of spinal cord varied from no visible hemorrhage to extensive hemorrhage. In both adult and infant spinal cords, hemorrhage was often concentrated around the mid-coronal plane, and tended to form a diagonal band centered at the level of T13 and angled at ∼45 degrees to the direction of dislocation (Fig. 2). It also affected both gray and white matter areas of the spinal cord (Fig. 2). This diagonal pattern is consistent with a shear injury. The neural tissue around areas of hemorrhage also often featured a loss of normal architecture.

Representative images of infant
For both infant and adult cohorts, the normalized volume of hemorrhage increased with increasing displacement (Fig. 3A). As such, magnitude of dislocation (displacement) had a significant effect on the normalized volume of hemorrhage (Table 2; p=0.028). For each 1 mm increase in injury displacement (scaled up for infants by 2.5), there was an increase in normalized volume of hemorrhage by 1.9×10−3 mm3. However, there was no obvious pattern found between normalized volume of hemorrhage and speed (Fig. 3B; Table 2; p=0.427). Infants appeared to have a higher normalized volume of hemorrhage than adults across all injury conditions, suggesting that age had a significant effect on the volume of hemorrhage (Fig. 3; Table 2; p<0.001). The normalized volume of hemorrhage was 3.3×10−3 mm3 greater for infants than for adults. There were no interactions found between age and displacement.
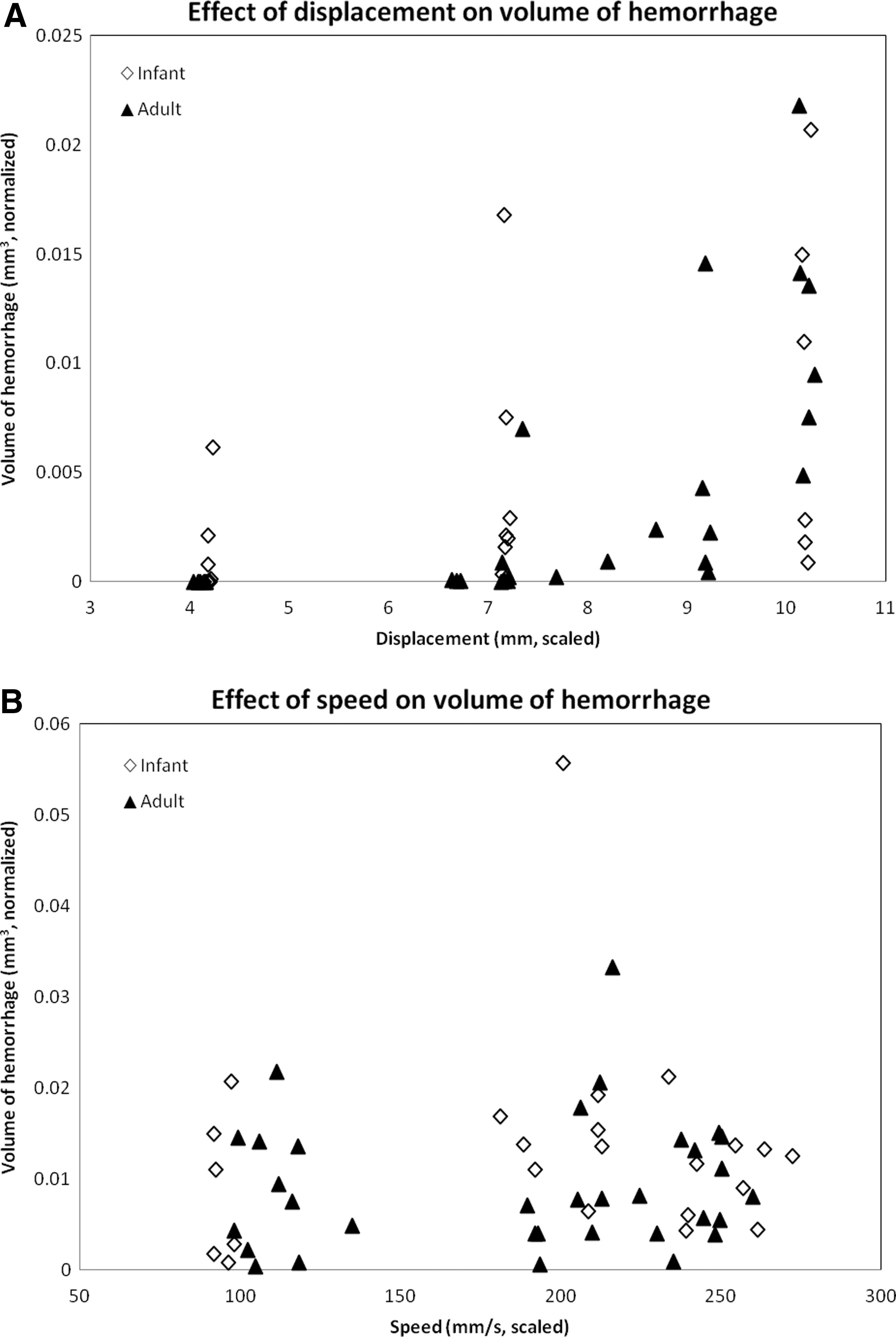
Variation in normalized hemorrhage volume with displacement
Summary of Statistical Outputs for the Effects of Age, Speed, and Displacement on Volume of Hemorrhage
p<0.05
Axonal injury
Injured axons staining positive for β-APP were observed in varying quantities for both adult and infant spinal cord sections. This ranged from no positive staining to extensive staining throughout white matter around the lesion. No injured axons were observed in the gray matter of the spinal cord. Injured axons typically formed retraction balls with a bulb-like appearance caused by cytoskeleton disruption (Fig. 4i-iv).

Representative images of infant
In both adult and infant cohorts, total area of normalized positive β-APP staining increased with increasing displacement (Fig. 5A; Table 3; p<0.001). An increase in displacement by 1 mm increased the area of axonal injury by 2.2×10−1mm2. Speed did not have a significant effect on the area of axonal injury (Fig. 5B; p=0.726). Individually, age also did not have a significant effect on the area of axonal injury (Fig. 5; p=0.463); however, displacement had a different effect on infants than on adults (Table 3; age–displacement interaction p=0.003). This indicates that the interaction between age and displacement has a greater effect on axonal injury severity than age alone.
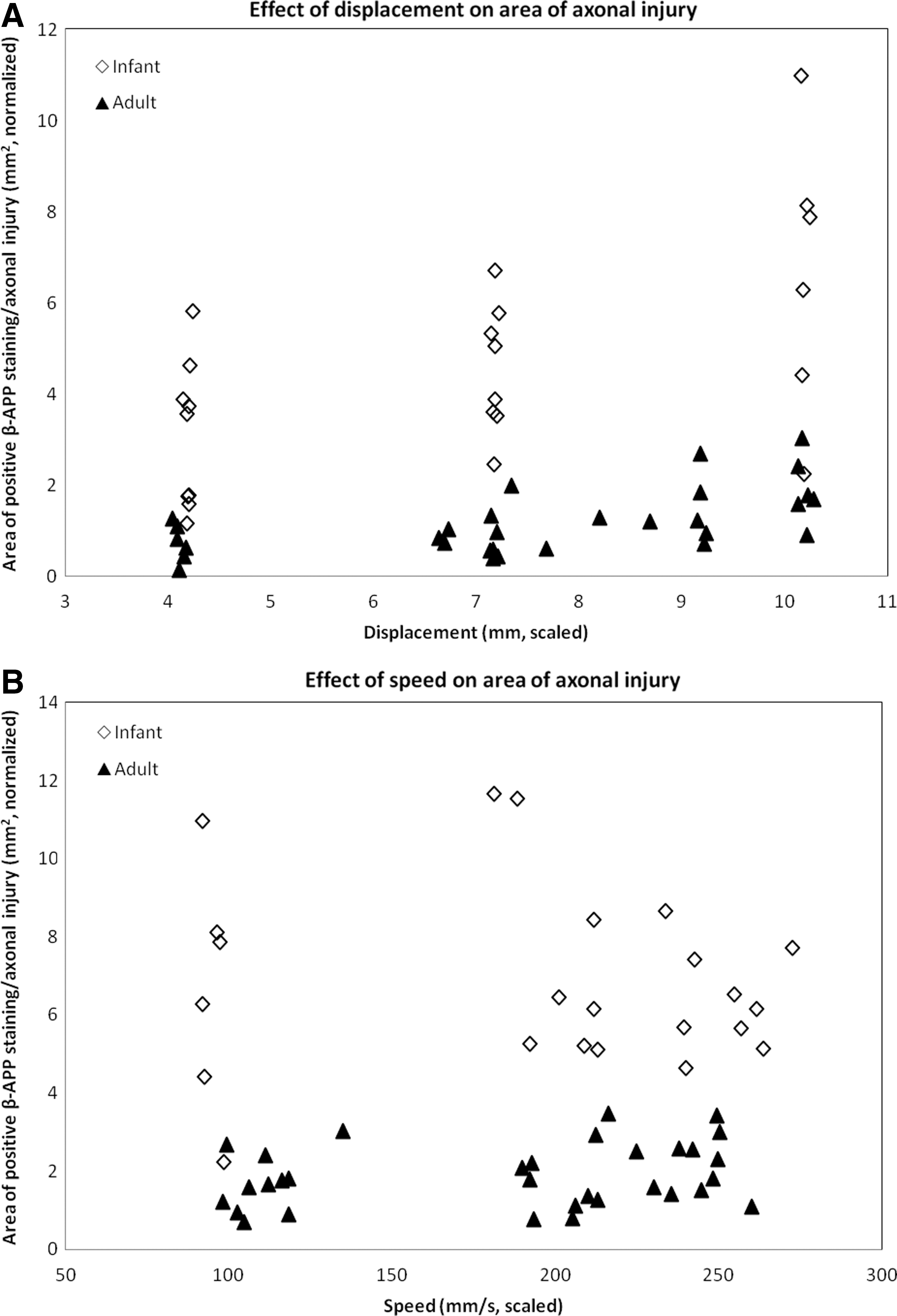
Effects of displacement
Summary of Statistical Outputs for the Effects of Age, Speed, and Displacement on the Total Area of Axonal Injury
p<0.05
The highest densities of injured axons were evident in the white matter along a band ∼45 degrees to the direction of dislocation in both adults and infants, and this pattern of injury was consistent with a shear injury. Most injured axons were found in the areas rostral to the lesion epicenter in the left lateral funiculus (Fig. 4, images i, iii), caudal to the lesion epicenter in the right lateral funiculus (Fig. 4, images ii, iv) and at the lesion epicenter in the dorsal and ventral funiculi. This cross-sectional pattern was significant in the dorsal and left lateral funiculi but not the ventral or right lateral funiculi (Table 4; dorsal: p=0.013, ventral: p=0.452, left lateral: p=0.002, right lateral: p=0.549).
Summary of Statistical Outputs for the Effects of Age, Speed and Displacement on the Pattern of Axonal Injury by Funiculi (p-Values)
p<0.05
Within all white matter funiculi, the area of axonal injury increased with increasing displacement and was greater in infants than in adults (Fig. 6; Table 4; all funiculi: p<0.001). Speed, however, did not have an effect on the area of axonal injury in any of the funiculi (Fig. 7; Table 4; dorsal: p=0.501, ventral: p=0.075, left lateral: p=0.454, right lateral: p=0.819).
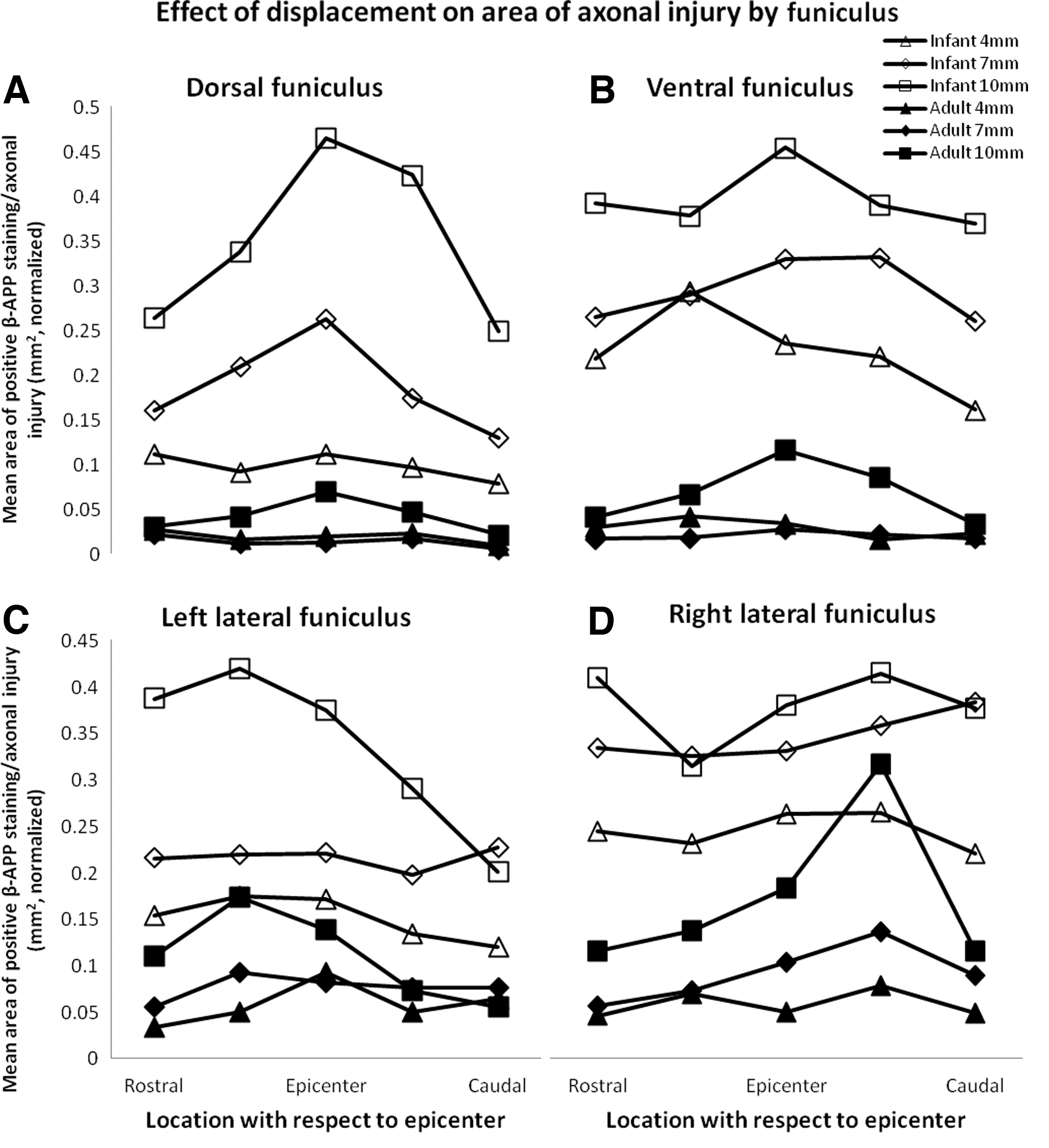
Effect of displacement on the area of β-amyloid precursor protein (β-APP) for adults and infants. Normalized means of area of β-APP staining are demonstrated for dorsal
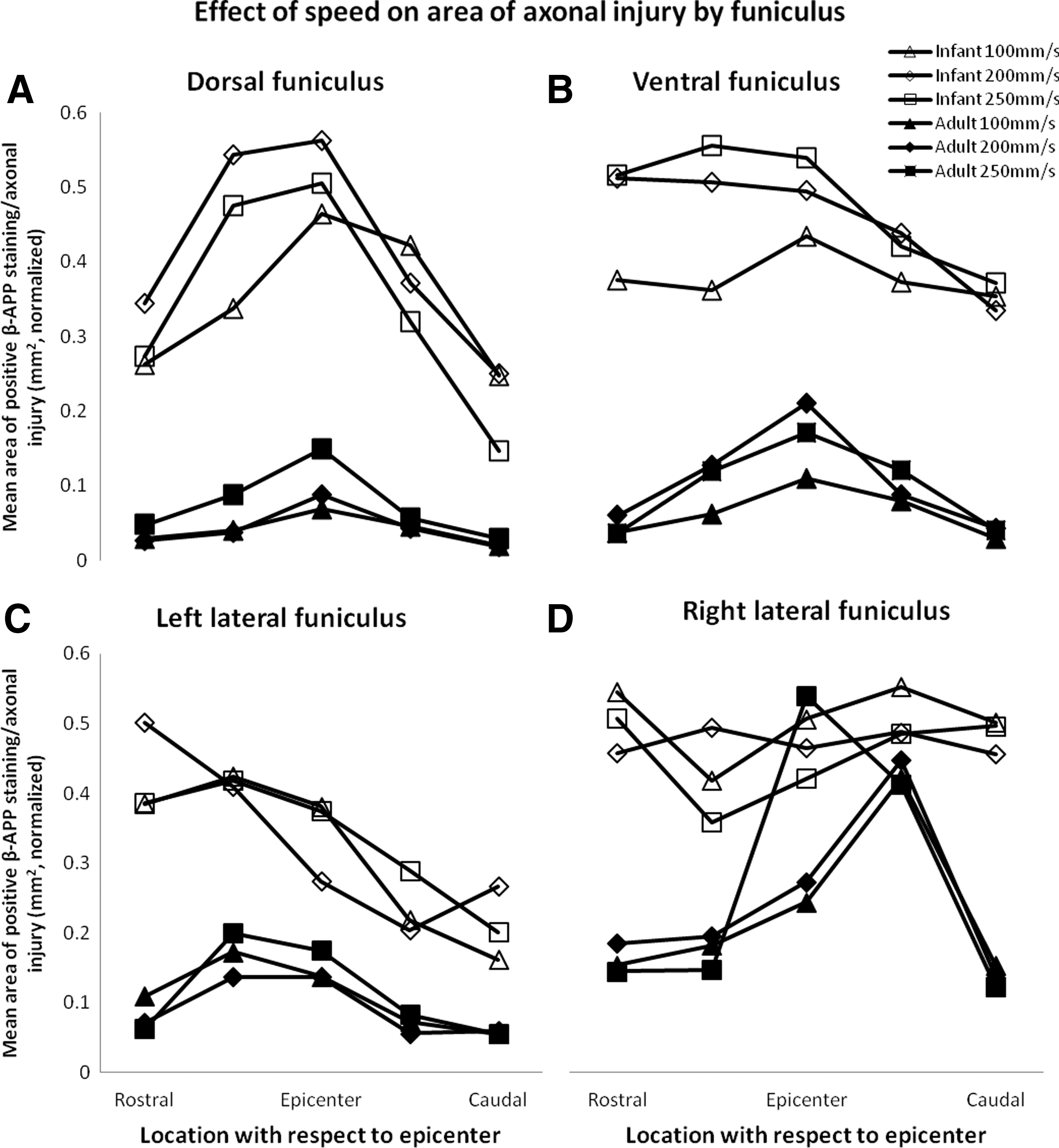
Effect of speed on the area of β-amyloid precursor protein (β-APP) for adults and infants. Normalized means of area of β-APP staining are demonstrated for dorsal
Animals in the infant cohort had an increased amount of positive β-APP staining in ventral and dorsal funiculi than in lateral funiculi when compared with the adult cohort. As such, age had a significant effect on the dorsal:lateral funiculus area of axonal injury ratio and the ventral:lateral funiculus area of axonal injury ratio such that infants had higher dorsal:lateral and ventral:lateral area of axonal injury ratios than adults (Table 5; p<0.001, p<0.001). This suggests that infant spinal cords were injured preferentially in dorsal and ventral funiculi rather than in lateral funiculi when compared with adult spinal cords.
Summary of Statistical Outputs for the Effects of Age, Speed and Displacement on the Ratio of Axonal Injury Between Funiculi
p<0.05
Discussion
Effects of speed and displacement on the severity of SCI in infants and adults
The current study found that infants have more severe SCI than do adults, across a wide range of injury conditions, using a vertebral dislocation model. Further, infant rats were found to have a different distribution of injury within the spinal cord than adults. This is the first study to investigate the effects of different spinal cord loading conditions on infant rats with a vertebral dislocation model of SCI, and the first to quantify the effects of age and displacement in a vertebral dislocation injury.
Previous studies in this area are limited. Vertebral dislocation has only been used as a model for SCI on a few occasions, 6 –8,20 and experimental research in SCI has rarely looked at mechanisms in children. Our finding that infants had more severe SCI than adults is consistent with epidemiological and experimental studies. 5,8,9 Specifically, Clarke and Bilston found, using a vertebral dislocation model and a single (scaled) injury condition, that normalized axonal injury density (as a marker of acute SCI severity) was significantly higher in infants than in adults. 8
Age has rarely been studied in experimental SCI studies, and the reasons why infants sustain more severe SCI than adults do is not known. It is possible that a combination of more severe primary tissue damage coupled with differences in pathophysiology of secondary SCI cause the clinical differences seen in adults and children with SCI. This is consistent with a recent study of contusion SCI, which observed a longer period of secondary ventral horn motor neuron loss in infants than in adults. 11 Further, it has been speculated that spinal cord tissue composition and spine and spinal cord biomechanical properties may contribute to the observed differences in primary injury severity. 8
Similarly, the effects of speed and displacement on severity of SCI in vivo have rarely been studied. Previously, Fiford et al., using a vertebral dislocation model, demonstrated that increasing the injury parameters of speed and displacement together resulted in increased acute SCI severity. 7 They did not, however, vary these parameters independently.
Our finding that displacement increased SCI severity is consistent with studies that used contusion models of SCI. Ghasemlou et al., using mice, found that spinal cord displacement correlated strongly with motoneuron loss. 21 Similarly, Kearney et al., using a ferret model of contusion SCI, demonstrated that depth of compression (displacement) was a significant factor in the severity of functional and anatomical spinal cord damage, independent of speed. 22 In our model, it is likely that a greater vertebral dislocation resulted in a more severe SCI, because the spinal cord had been deformed to a greater degree.
In this study using a vertebral dislocation model, speed of vertebral dislocation did not have a significant effect on SCI severity. This is somewhat inconsistent with the literature, in that a few studies investigating the effects of speed found significant effects on SCI severity. Kearney et al. demonstrated in ferrets that rate of compression (speed) had a significant effect on functional and anatomical measures of SCI severity, independent of displacement. 22 Similarly, Sparrey et al. found that the volume of hemorrhage in white matter was dependent upon velocity, using a rodent contusion model of SCI. 15 In vitro studies of guinea pig spinal cord white matter and giant squid axons have also demonstrated that strain rate (speed) has a significant effect on functional and structural damage. 16,23 However, using an in vitro hippocampal slice model of traumatic brain injury, Cater et al., found that cell death was dependent upon tissue strain but not tissue strain rate. 24 This is consistent with the findings of the current study. Further, using an impactor model of contusion SCI in mice, Kim et al. found that impact speed was not related to severity of SCI. 25 Regardless, there have not been any previous studies on the primary effects of speed on the severity of SCI using a vertebral dislocation model.
There are several possible reasons for the inconsistency between our findings about the effect of speed on SCI severity and the literature. First, the vertebral dislocation model may “dampen” the effects of speed on SCI severity, caused by space or “slack” between the spinal column and the spinal cord. Existing experimental studies of the effects of speed on injury severity use either in vitro uniaxial loading or contusion models of SCI. 15,16,22,23 In these situations, when force is applied to the spinal cord or axon directly, the speed at which the injury is induced can be precisely controlled. A model of vertebral dislocation injury, in which the injury is created by movement of the vertebral column rather than directly loading the spinal cord, introduces the additional complexity of the vertebral column and, therefore, may be unable to detect an effect of speed on SCI severity. Notably, as human SCI tends to be caused by motion of the vertebral column, our findings suggest that although precisely controlled experiments demonstrate an effect of speed on injury severity, the speed of vertebral dislocation may not be an important factor in real world human SCI.
It is also possible that the range of speeds studied were not broad enough to capture differences in SCI severity. For example, in this study, the speeds for the adult group ranged from 100 to 250 mm/s, whereas the study by Sparrey et al. that demonstrated an effect of speed on SCI severity used 3–300 mm/s. 15 It is also possible that there is a speed threshold for SCI, just as there appears to be a displacement threshold for adult rats in the current study (Figs. 3A and 5A). Performing an investigation of injury biomechanical parameters using a wider range of speeds may assist in determining injury thresholds and help to define SCI criteria.
Another factor to consider is that in a viscoelastic tissue such as the spinal cord, there is an interaction between displacement and speed when determining SCI severity. Kearney et al. found a significant interaction between rate and depth of compression using a ferret contusion model of SCI. 22 This suggested that at low speed, maximum compression depth was the best predictor of SCI severity, whereas with increasing speed, a lower compression depth was required to achieve an equivalent injury. 22 Similarly, a biomechanical study of in vitro adult and infant rat spinal cords found that modulus (a measure of tissue stiffness) increased with increasing strain rate. 17 This reinforces that the spinal cord responds differently to displacement depending upon the rate of the displacement. These properties of the spinal cord may result in a “threshold” effect, in which injuries with a displacement above a certain level are so severe that the effects of speed are negligible. To detect the effects of speed on SCI severity and identify injury thresholds using this vertebral dislocation model, a lower displacement may be required. However, the study by Kim et al. found, using a low displacement impactor model of SCI, that speed did not determine severity of SCI. 25 Future studies could investigate this further, and potentially identify the precise displacement at which speed becomes an important factor in the severity of SCI or the precise speed at which displacement becomes significant. This would not only increase our knowledge of the biomechanics of the spine and spinal cord, but also provide valuable information for the future development of injury criteria that could link spinal kinematics to cord injury.
Implications
Using a rodent model, the current study has quantified the severity of SCI in terms of the injury biomechanics. This has the potential to contribute to the development of clinical policies for the management of SCI and the development of motor vehicle safety devices. By knowing which components of injury are most critical to severity of injury, we can develop algorithms for identifying patients at highest risk of severe SCI, or motor vehicle safety devices that are more effective at preventing SCI and minimizing SCI severity. Further, the finding that infants have both more severe and different patterns of injuries than adults supports the need for child-specific interventions. As such, our findings suggest that pediatric SCI may need to be treated differently than adult SCI, and that children may require child-specific motor vehicle safety devices for adequate protection against SCI.
Limitations
This study has several limitations that need to be considered when interpreting its findings. The infant cohort was defined as rats 13–15 days postnatal. This age was chosen to approximately correspond to a 1–2-year-old human child, based on the appearance of secondary ossification centers in the humerus and femur. 26 It is acknowledged that the development of human and rat bones do not directly correspond, which makes determining the equivalent human age of a 13–15-day-old rat difficult. However, it is widely accepted that the human spine becomes functionally equivalent to that of an adult at the age of ∼8 years. 27 -29 Therefore, the ages for our adult and infant rats were chosen to ensure infants were equivalently <8 human years, and that adults were equivalently >8 human years.
A model of vertebral dislocation SCI was used in this study. This is a closed column SCI model in which the SCI is induced by relative motion of the vertebrae, as commonly occurs in human SCI. However, closed column models have the limitation that the spinal cord loading conditions are not known or directly controlled. There may, therefore, be variations in how vertebral displacements are transmitted to the spinal cord among animals, based on individual anatomical variation. To ensure as much consistency as possible, the injury device was computer controlled, surgery for all animals in the study was conducted by one researcher, and the injury groups were randomized so that any variation was expected to be distributed evenly among injury groups.
In this study, injury was induced at T12-L1 in both adult and infant animals. Because of the different growth rate of the spinal cord and vertebrae during development, inducing the injury at the same vertebral level in adult and infant animals may have resulted in the dislocation being applied at different neurological levels, and, therefore, have had a small effect on the severity of injury. However, it was necessary to match the vertebral level of injury in this way to ensure equivalent vertebral loading mechanics.
To account for the difference in size between adult and infant rats, injury conditions were scaled by a factor of 2.5. This number was derived from the ratio between the mean measured nerve root separation of adult and infant spinal cords of the same age and weight at T12-13 in the coronal plane. This had the aim of matching the injury conditions based on strain and strain rate. It is recognized, however, that the biomechanical properties of the spinal cord and the interaction between the spine and spinal cord in vivo are complex and three dimensional. As such, it is impossible to be certain that the scaling factor of 2.5 was sufficient to precisely match strain and strain rate in this experiment. Also, to facilitate comparison between adult and infant data, volume of hemorrhage and area of axonal injury were normalized to a nominal volume or area (the estimated volume of the spinal cord at T12-13, and the estimated cross-sectional area of the spinal cord in the coronal plane at T12-13). For axonal injury, however, this makes the assumption that the total number of axons in the spinal cord was the same for adults and infants. This is unknown; therefore, it is important to consider these normalizations as approximations of the severity of injury.
Analysis of axonal injury used a manual region of interest selection and automatic thresholding technique to calculate the percentage area of the images that exceeded a specific threshold. This aimed to minimize false positives and negatives. However, background staining and variation between staining runs are recognized problems in immunohistochemistry; therefore, the order of staining was randomized to spread any variation evenly among all injury groups and the threshold level was adjusted for each staining batch. To ensure consistency among these batches, sections from areas in which no axonal injury was expected were randomly chosen, and the results of automatic thresholding were compared using a Student's t test. This demonstrated no difference in the thresholded level of background staining, and, therefore, of consistency in the threshold levels chosen for each batch.
Finally, the current study only examined SCI at 5 h post-injury, and therefore, is limited to the acute phase of SCI. The pathophysiology of SCI has been extensively studied and is accepted to involve a biphasic process. 1,30 –32 The primary phase of injury lasts for several hours and features hemorrhage, traumatic axonal necrosis, and edema. 31,33,34 The secondary phase continues for weeks after injury and features free radical formation, excitotoxicity, apoptosis, and inflammation. 1,30,32 The survival time of 5 h used in this study has been shown in pilot studies with this injury device to be sufficient to characterize hemorrhage and acute axonal injury, but to better understand the differences between adults and infants in the secondary phase of injury and subsequent recovery, a longer survival period would be required. An appropriate method of permanent spinal stabilization is currently being trialed to facilitate such studies.
Conclusion
In conclusion, we found, using a rodent model of vertebral dislocation SCI, that magnitude of vertebral dislocation (displacement) and age have significant and quantifiable effects on acute SCI severity in the form of volume of hemorrhage and area of axonal injury. However, we also found that speed of dislocation did not have a significant effect on the severity of SCI. Whereas this could be a genuine lack of effect, it could also be the result of the interaction between the spine and spinal cord during the dislocation, a displacement thresholding effect on speed, or the range of speeds chosen in this study. This study is the first to demonstrate quantifiable and predictable effects of displacement and age on the severity of vertebral dislocation SCI. This greatly increases our understanding of the biomechanics of the spine and spinal cord, and may have significant implications in the field of SCI prevention by demonstrating that age and magnitude of spinal motion affect SCI severity.
Footnotes
Acknowledgments
This research was funded by the SpineCare Foundation. Drs. Clarke and Bilston are supported by National Health and Medical Research Council of Australia (NHMRC) research fellowships.
Author Disclosure Statement
No competing financial interests exist.