
Abstract
Electrospinning is the process by which a scaffold containing micrometer and nanometer diameter fibers are drawn from a polymer solution or melt using a large voltage gradient between a polymer emitting source and a grounded collector. Ramakrishna and colleagues first investigated electrospun fibers for neural applications in 2004. After this initial study, electrospun fibers are increasingly investigated for neural tissue engineering applications. Electrospun fibers robustly support axonal regeneration within in vivo rodent models of spinal cord injury. These findings suggest the possibility of their eventual use within patients. Indeed, both spinal cord and peripheral nervous system regeneration research over the last several years shows that physical guidance cues induce recovery of limb, respiration, or bladder control in rodent models. Electrospun fibers may be an alternative to the peripheral nerve graft (PNG), because PNG autografts injure the patient and are limited in supply, and allografts risk host rejection. In addition, electrospun fibers can be engineered easily to confront new therapeutic challenges. Fibers can be modified to release therapies locally or can be physically modified to direct neural stem cell differentiation. This review summarizes the major findings and trends in the last decade of research, with a particular focus on spinal cord injury. This review also demonstrates how electrospun fibers can be used to study the central nervous system in vitro.
Spinal Cord Injury and Guidance Strategies for Regeneration
M
With an estimated 12,000 new cases of SCI added each year and at an individual cost of more than $1.5 million over the lifetime of a patient, 4 there is incentive to find a treatment option that restores function and reduces chronic injury. New methods provide regenerating neurons with both factors and guidance cues to overcome the barriers to regeneration.
Many cell- and drug-based therapies have been introduced for the purpose of restoring lost function. There are three primary approaches to SCI: (1) transplanting cells to repopulate the region, (2) reducing the physical and chemical barriers for regenerating cells, or (3) promoting growth through a series of factors. Transplantation of neural stem cells has received the most attention within the decade, and in the United States, there is one completed clinical trial and four ongoing clinical trials that use neural stem cells to repair SCI.
A recent alternative to stem cell-based therapies, however, uses a peripheral nerve graft (PNG), and this approach has produced evidence of functional recovery equivalent to pre-injury levels in animal models of SCI. PNGs use sections of nerve from the peripheral nervous system and can either be autografts or allografts. The first study that used a PNG in a SCI model showed partial recovery of limb function. 5 Studies from Houle and associates 6 and Alilain and colleagues 7 present an effective synergistic approach where a PNG is used with the glycosaminoglycan degrading enzyme chondroitinase ABC (ChABC). The synergistic approach partially restored limb function 6 and completely restored respiratory function. 7
Recently, an intercostal nerve graft in combination with ChABC and acidic fibroblast growth factor (aFGF) restored the complex set of systems involved in bladder control. 8 Strategies that promote functional recovery using a nerve graft to guide regenerating axons are an important divergence from past approaches that focused on the application of cells or biologically active agents alone.
The application of a biologically active agent alone has failed to restore lost function to the same extent as when a nerve graft is used, and this important concept was validated by Alilain and colleagues 7 in a rat model of SCI. ChABC (in the study by Alilain and colleagues 7 ) use within rodent injury models over the last decade was recently reviewed. 9 The appeal of ChABC stems from the ability of the enzyme to degrade glycosaminoglycans attached to the protein core of chondroitin sulfate proteoglycans (CSPGs). Glycosaminoglycans inhibit axonal extension through protein tyrosine phosphatase sigma (PTPσ) signaling. 10
Importantly, the study by Alilain and colleagues 7 demonstrated that the use of ChABC alone did not restore respiratory function to the same extent as when ChABC was combined with a PNG. 7 Thus, the inclusion of a guidance strategy is imperative when the experimental approach is focused on restoring lost function.
Electrospinning (presented in several recent reviews 11 –13 and briefly described later in this review) is a technique used to generate fibers with nano to micro scale diameters. Electrospun fibers mimic the scale and high surface area to volume ratio found in the extracellular matrix and are currently studied as synthetic nerve guides. Synthetic guides may be a desirable alternative to natural nerve guides because allografts may induce an inflammatory response and autografts create loss of function or sensation at the harvest site. Further, electrospinning of nano or microfibers can be performed using many types of FDA-approved polymers such as polylactic acid (PLA), polyglycolic acid (PGA), and polycaprolactone (PCL).
Pioneering work from the group at the National University of Singapore introduced electrospun fibers as a promising scaffold for neural regeneration a decade ago. 14,15 Since then, many studies have attempted to engineer electrospun fibers to better guide the extension of axons.
More recently, electrospun fibers have been used to create in vitro culture systems that recapitulate the topographical features observed in the native spinal cord. These new topographical tools are used to understand cell behavior on topography known to exist in the white matter tracts. For example, new in vitro culture systems have been created using patterned electrospun fibers and smooth films to study how astrocytes respond to local changes in surface topography 16,17 that may be similar to topographical changes after SCI. In subsequent sections of this review, electrospun fiber research over the past decade in the context of SCI is presented, both as a material to promote spinal cord regeneration and as tools to better understand how topography affects the response of cells known to exist in the central nervous system (CNS).
Fabrication of Fibers Using Electrospinning
Creation of polymer fibers from electrospinning is accomplished by applying a high voltage to a small bead of polymer solution residing on the tip of a needle. The high voltage propels a solvent/polymer jet toward a grounded collector. Over several milliseconds, the jet elongates to around 10,000 times its original length and solidifies (by solvent evaporation) as it whips unstably through the air. 18 Rapid elongation of the jet and the high charge density in the polymer cause the jet to bend into a series of coils. 19,20 As a consequence, fibers fall randomly onto the collection surface without any specific orientation. These fibers can be aligned, however, if collected on an oscillating collection plate, 21 a rotating disc, 22 or between two grounded plates. 23 By tuning these collection parameters, fiber orientation can range from random to highly aligned. 22,24 –26
The versatility of the electrospinning process allows researchers to observe the effects of both fiber alignment and fiber diameter on neural cell models. Electrospun fibers can be generated with diameters ranging from tens of microns to tens of nanometers, with the smallest diameter fibers containing fewer than 10 elongated polymer chains. 18 The simplest method of altering fiber diameter is to change the concentration of polymer in the electrospinning solution, where a higher concentration of polymer leads to a larger fiber diameter 27 because of an increased solution viscosity. 28,29 Other factors, however, influence fiber diameter such as the dielectric constant of the electrospinning solvent, 30 the distance between the electrospinning tip and the collector, 31 and vapor pressure of the solvent. 32 These methods of controlling fiber diameter make electrospinning a versatile tool for scientific research.
The following sections present research that has investigated how neurite extension was affected by fiber geometry (fiber alignment and diameter), the material from which the fiber was made (including surface chemistry modification), and drug and protein delivery from the fibers. Fiber geometry is presented first because fiber geometry is easily modified and generally results in a drastic change in neurite extension. Material and chemical composition of the fibers is presented next because attempts to improve neurite guidance by changing materials or chemical modification tend to produce marginal differences in neurite extension compared with traditional materials alone (polycaprolactone or polylactic acid). Finally, drug and protein delivery are presented last because of the complexity of drug release and other considerations such as protein denaturation.
The importance of electrospun fiber geometry in directing the extension of neurites
Electrospun fiber alignment, neurite extension, and axonal guidance
The first study to combine electrospun fibers with nerve cells was conducted in 2004. Neural stem cells (multipotent neural cell line, C17.2
33
) were placed on randomly oriented poly-
The ability of aligned electrospun fibers to direct the extension of neural processes is now routinely reported, using both primary neurons and neural cell lines. Dorsal root ganglia (DRG) cultures are useful in vitro models for the study of a biomaterial's ability to direct neurite extension. 37 DRG extend longer neurites than neurites extending from cell line cultures. Thus, if the electrospun fiber approach is meant to induce longer neurite extension, then a DRG model may be more appropriate than a cell line.
DRG placement onto electrospun fibers routinely occurs within studies demonstrating the ability of aligned electrospun fibers to direct neurite extension. The correlation between fiber alignment and neurite extension from embryonic rat DRG was demonstrated in pioneering experiments by Corey and coworkers 38 using aligned PLLA electrospun fibers. In general, electrospun fiber scaffolds with greater fiber alignment result in a more elongated DRG body, with longer neurites that align to the orientation of fibers compared with results where DRG were cultured on randomly organized fibers. The study by Corey and coworkers 38 also included the use of neuroblastoma cell lines (neuroblastoma cell lines SH-EP 39 and SH-SY5Y 40 ), and these cells extended neurites along the electrospun fibers.
In an article published by Chow and colleagues, 41 DRG from embryonic day 16 rats were cultured on both aligned and randomly oriented polydioxanone (PDS) fibers. This study was unique because the DRG were cultured on top of electrospun fibers with adherent astrocytes. The presence of astrocytes significantly increased the length of neurites from DRG cultured on the electrospun fibers.
Schnell and associates 42 cultured embryonic day 10 DRG from chicken on aligned electrospun fibers composed of PCL or a blend of collagen type I and PCL (mixed 1:3 [w/w]) to study the ability of collagen to promote long extension of neurites. Interestingly, Schnell and colleagues 42 collected fibers on a surface coated with star-poly(ethylene glycol), which was used to adhere fibers to the collection surface, but also prevented cell adhesion to the collection surface. There was no comparison of neurite lengths when DRG were cultured on aligned or randomly oriented fibers, but neurites from whole and dissociated DRG traveled along the length of both PCL and PCL/blended fibers. The DRG experiments from these initial studies confirm the ability of electrospun fibers to guide the extension of neurites, with aligned fibers inducing the longest neurite extension.
From these initial studies, one can easily conclude that neurites extend parallel to fiber orientation and extend longer processes when the electrospun fibers are aligned. Since these initial studies, several publications document the ability of aligned, electrospun fibers to induce long neurite extension from DRG. Several subsequent studies document similar results stating that different cells from different animal sources all extend neurites along aligned fibers. DRG from postnatal day 1, 43 day 4, and day 16 48 rat pups, 44,45 DRG from day 8 chicken, 46,47 PC12 cells (rat adrenal pheochromocytoma cells 49 ), 50,51 and C17.2 cells 52,53 all increased neurite extension along aligned, electrospun fibers compared with neurite extension observed on randomly oriented fiber controls.
Also, different types of neurons respond to aligned fibers in unique ways. In a study comparing DRG neurite outgrowth and hippocampal neurite outgrowth, DRG neurites extended along the length of aligned fibers while hippocampal neurites extended both parallel and perpendicular to the aligned fibers. 54 Recently, the need for fibers to be aligned to direct neurite extension was challenged in a study revealing neurite extension can occur perpendicular to the direction of fiber alignment when the fiber density was high (3000 fibers/mm). 55 The study, however, did not quantify fiber alignment, so it is difficult to determine whether fiber alignment played a role in their results.
From the above studies, neurites extend to a greater extent on aligned fibers compared with neurite extension on randomly oriented fibers. The literature presents two explanations for why aligned fibers more efficiently direct neurite extension. The first explanation is supported by observations in cell lines. C17.2 and PC12 derived neurons are more viable when cultured on aligned fibers compared with randomly oriented fibers, 24,56 –58 and increased viability was also confirmed when primary neural progenitor cells from rats were cultured on fibers. 59
Most of these studies assess cell viability with either a metabolic assay (without normalizing to cell count) or by a simple cell count, meaning an increase in cellular viability could merely represent an increase in the rate of cellular proliferation. If fibers enable neural cells to attach and cells in culture possess greater viability, then it is reasonable to assume that cells on fibers will extend longer processes than cells having difficulty attaching to the substrate.
The second explanation revolves on the premise that aligned fibers do not have as many barriers to extension as do randomly organized fibers. Studies analyzing the effect of fiber crossing inhibiting neurite extension reveal the ability of crossed fibers to stop neurite extension. As stated previously, a study from Corey and coworkers 38 demonstrated that increased neurite extension correlated with increased fiber alignment. In addition, a study by Wang and colleagues 25 developed a method for generating highly aligned electrospun fibers. This study also introduced fiber scaffolds with two fiber layers of highly aligned PLLA fibers oriented at 45-degree angles to each other. Interestingly, when DRG from day 9 chickens were cultured on these scaffolds, neurites extended along the top layer of fibers until they came into contact with the underlying fibers oriented at a 45-degree angle, at which point the neurites did not extend. The results from these two studies demonstrate that aligned fibers improve total neurite extension and guide neurites along the length of the fibers, while randomly oriented fibers restrict neurite extension.
The importance of fiber alignment within in vivo models of SCI was also demonstrated by Hurtado and associates. 45 To apply the fibers within a three-dimensional in vivo SCI environment, an innovative approach was used to place fibers within a conduit construct (Fig. 1). Specifically, in this study, Hurtado and associates 45 placed aligned or randomly oriented fibers in such a conduit structure within a complete transection rat model (T9–T10, 3 mm gap) of acute SCI. 45 In this study, Hurtado and associates 45 verified that aligned PLLA microfibers permitted the greatest extension of pioneering axons (2055 μm) 4 weeks after implantation. The result was statistically different from axonal extension in rats with randomly oriented PLLA fibers (1162 μm) or a PLLA film (413 μm).
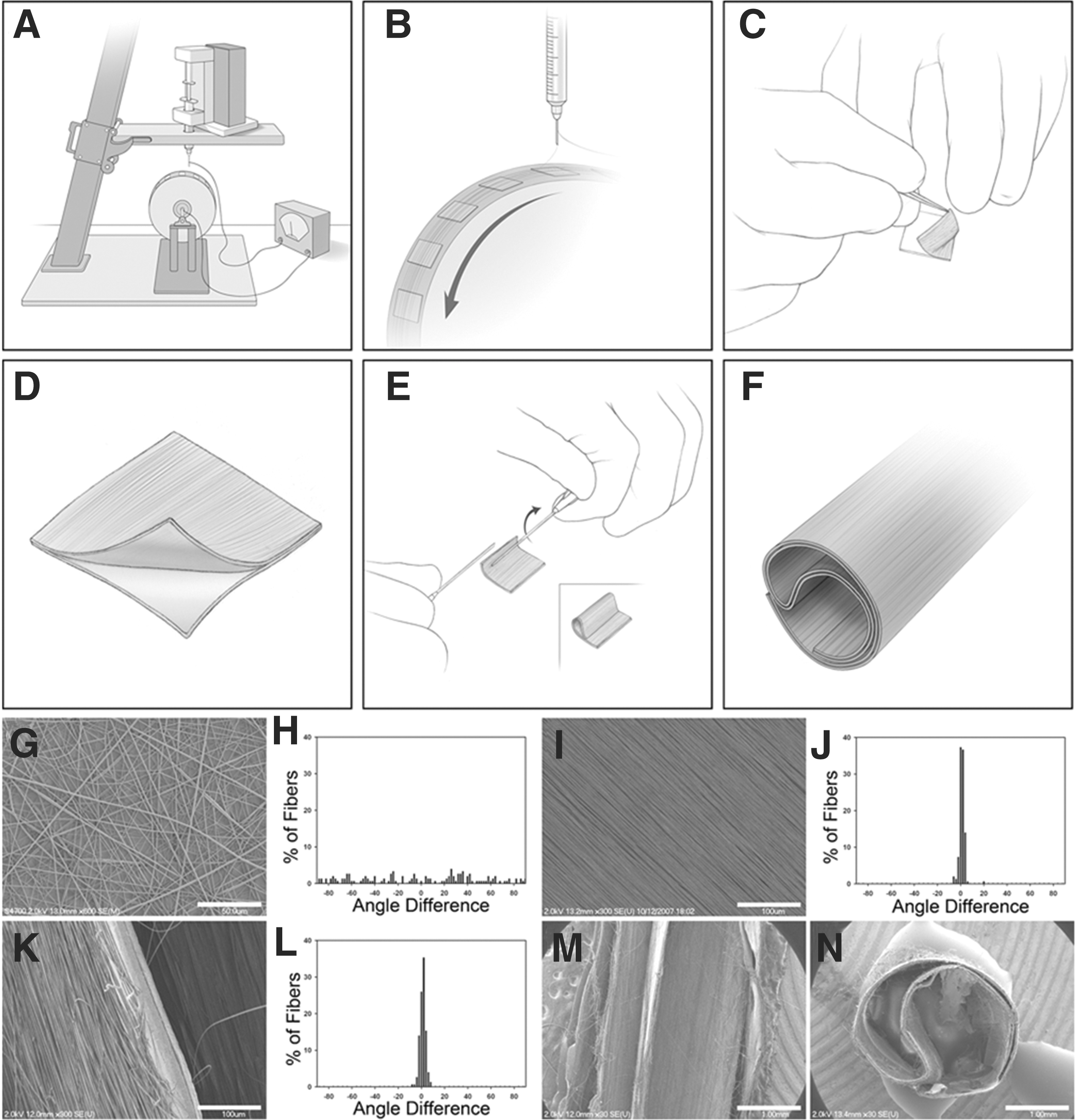
The panel A–F provides a schematic representation of conduit fabrication. PLLA fibers are electrospun onto coverslips coated with PLLA films on a rotating mandrel (
In contrast to the study from Hurtado and associates, 45 Liu and coworkers 60 found that rats with a complete spinal cord transection (C3, 2.5 mm gap) had little neurite sprouting into the lesion regardless of scaffold alignment. The differences between these two studies could be the result of differences in fiber diameter, because Hurtado and associates 45 used microscale fiber diameters while Liu and coworkers 60 used nanoscale fiber diameters (nearly an order of magnitude difference). Material selection may have also been a factor, because the study from Hurtado and associates 45 used PLLA, while the study from Liu and coworkers 60 used collagen.
Because of the discrepancy between the results of these two studies, it may be beneficial to compare with trends seen in the peripheral nervous system. In a rat tibial nerve model, aligned poly(acrylonitrile-co-methylacrylate) fibers improved axonal extension compared with extension on random electrospun fibers. 43 Using the same model, aligned PCL fibers yielded increased axonal density compared with randomly aligned PCL scaffolds. 61 Therefore, it may be possible to attribute results of the study by Liu and coworkers 60 in the spinal cord to the use of collagen and small diameter fibers (<400 nm), two factors that could potentially reduce axonal guidance and extension (described later in this review).
Few in vivo studies, however, have been performed using electrospun scaffolds in the spinal cord. While evidence supports electrospun fiber alignment as a strong factor in improving axonal guidance, further work is required to understand how fiber alignment aids in axonal guidance and extension in vivo.
Neurite response to electrospun fiber scaffolds of varying diameter
Although there was significant focus on developing fibers with nanoscale diameters when electrospun fibers were first being investigated as neuron regeneration scaffolds, studies focusing on the effects of fiber diameter suggest a unique response of neurons and neural stem cells to fibers on the nanoscale versus the microscale. In 2005, Yang and colleagues 34 placed C17.2 cells on aligned and randomly oriented electrospun PLLA fibers with a mean diameter of 300 nm (nanofibers) and on fibers with a mean diameter of 1.5 μm (microfibers). From this study, aligned nanofibers induced longer neurite outgrowth than fibers with diameters on the microscale.
In addition to this finding, a large portion of the C17.2 cells preferentially differentiated into neurons on nanofibers (∼80%) compared with microfibers (∼40%) regardless of fiber orientation. 34 This initial study examining the role of fiber diameter in inducing neurite extension also suggests fiber diameter is as important as alignment in directing the extension of neural processes.
The results from C17.2 cells cultured on fibers of varying diameter initially proposed the idea of nanoscale diameter fibers being superior to microscale diameters for neural regeneration applications. Nanoscale fiber diameters, however, do not always provide optimum results when directing the extension of neurites. He and associates 52 created unique electrospun PLLA scaffolds (both aligned and randomly oriented) with diameters ranging from 300 nm to 900 nm in 200 nm increments. C17.2 cells cultured on scaffolds with a mean fiber diameter of 500 nm induced the longest neurite extension and the greatest amount of neuronal differentiation. 52
Unlike their previous study, 34 C17.2 differentiation was highly dependent on fiber alignment as more cells differentiated into a neuronal lineage on aligned fibers regardless of fiber diameter. This result led the authors to conclude that fibers with a 500 nm diameter were optimal for neurite guidance. The authors did acknowledge, however, that their observation might be a result of the high degree of neuronal differentiation of the C17.2 cells because neurite extension was longest on scaffolds with the greatest amount of neuronal differentiation.
Similarly, electrospun silk fibers with diameters on the nanoscale (400 nm) improved neurite extension in a coculture of neurons and astrocytes from postnatal day 1 rat pups versus neurons and astrocytes cultured on microscale diameter fibers (1200 nm). 62 Yao and colleagues 51 performed a similar study using PC12 cells cultured on a range of larger diameter PCL fibers (800 nm to 8800 nm). In this study, PC12 cells generated the longest neurites on aligned fibers with mean diameters of 3.7 μm and 5.9 μm. A similar result was observed when ND7/23 sensory neuronal cells (neuroblastoma cell line 63 ) were cultured onto PLLA fibers, where neuronal cells cultured on fibers with mean diameter of 750 nm had shorter neurites than neuronal cells cultured on fibers with diameters of 5–6 μm. 64
Daud and coworkers 66 cultured NG108-15 cells (neuroblastoma cell line 65 ) or adult rat DRG onto PCL fibers with diameters similar to those reported by Yao and colleagues 51 (1 μm to 8 μm). The neuroblastoma cells cultured on 8 μm fibers had longer neurites than neuroblastoma cell lines cultured on fibers with smaller diameters. When neuroblastoma cells were cocultured with Schwann cells, however, neurite extension was the same on every scaffold regardless of fiber diameter, and neurite extension was similar to the neuroblastoma cells cultured without Schwann cells on the 8 μm fibers. Interestingly, neurite extension from rat DRG was greatest when cultured on fibers with a 1 μm diameter. These studies suggest a micron scale fiber diameter is more suitable for neurite extension (>750 nm).
It is difficult to compare the studies from Yang and associates 34 and He and colleagues, 52 however, with the studies from Daud and coworkers 66 and Yao and colleagues 51 because there was no overlap in the diameter of the fibers used between the studies. Wang and coworkers 67 used a range of fiber diameters that sufficiently overlap the other studies (diameters ranging from 293 nm to 1325 nm); they cultured embryonic day 9 chicken DRG on PLLA fibers with diameters that were large (1325 nm), intermediate (759 nm), or small (293 nm). DRG cultured on electrospun fibers with large and intermediate diameters had significantly longer neurites than DRG cultured on small diameter fibers.
The study by Wang and coworkers 67 helps to bridge the gap of many studies that propose a fiber diameter between 750 nm and 5 μm may be most suitable for instigating the greatest neurite extension. It is possible that discrepancies are merely a result of cell source (cell line vs. primary cells), and this is supported by the study from Daud and coworkers. 66 Unfortunately, there is no study that investigates how fiber diameter alters neurite extension in vivo in the central or peripheral nervous system. Regardless, a consistent theme throughout all of the in vitro studies is that fiber diameter significantly alters neurite extension, but more research must be performed to examine the significance of fiber diameter in animal models of SCI.
An illustration summarizing the general results of several independent studies presented above reveals the ability of fibers with specific fiber diameters and fiber alignment to direct the extension of neuritis from DRG (Fig. 2). Specifically, highly aligned fibers (with diameters larger than 750 nm) strongly direct neurite extension (Fig. 2A), while aligned fibers with fiber diameters less than 750 nm direct neurite elongation to a lesser degree (Fig. 2B). Randomly organized fibers (with diameters greater than 750 nm) effectively direct neurite outgrowth, but the outgrowth is randomly distributed from the neural explant (Fig. 2C).
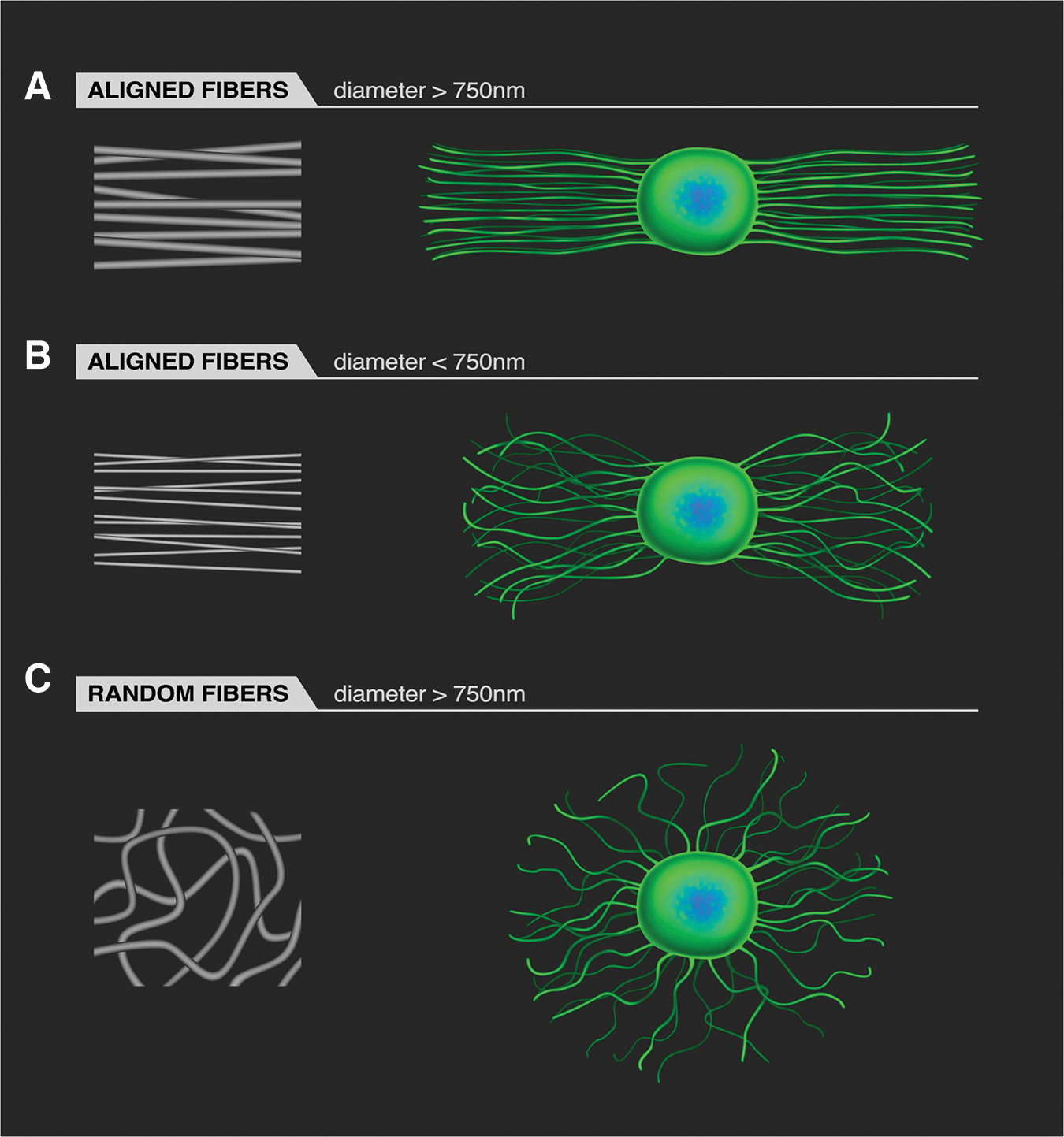
An illustration of how fiber geometry affects neurite extension from dorsal root ganglia (DRG) on electrospun fibers. The blue/green objects in the illustrations on the right side of the figure indicate the DRG body. When electrospun fibers are aligned and the fiber diameter is above 750 nm, neurites extend along the fibers (
Polymer material selection/surface modification
In addition to the geometric properties of fiber orientation and diameter, material selection may also influence neurite extension. Synthetic, degradable polymers have been used to electrospin fibers for neural applications including PLLA, poly (
Biodegradable, synthetic polyesters can be mixed with proteins (such as laminin, fibronectin, or L1) or peptide sequences (such as YIGSR, RGD, or IKVAV) before electrospinning to incorporate these biological molecules within the fibers. The above molecules can also be placed onto the surface of fibers modified through covalent coupling of proteins or coupling of integrin binding peptides to support the extension of neurites. 69 –71
Synthetic polymers can be mixed with proteins in the electrospinning solution and subsequently electrospun, but there is some evidence that proteins such as collagen denature during the process of electrospinning. 72 Further, addition of a protein to the electrospinning solution can significantly alter fiber morphology (e.g., fiber alignment and diameter) because proteins are charge carriers. 73 In addition, there is no guarantee that the added protein or peptide will remain functional or in a high enough concentration at the surface to significantly improve extension of neurites.
An alternative method of adding proteins to fibers is through covalent linkage or physical adsorption. To promote protein adsorption to hydrophobic polymers, surface treatments are frequently made to the fibers before exposure to proteins or peptides. Surface treatment affects the hydrophobicity and surface free energy, which alters the type of protein adsorbing to the fibers. 74 Decreasing the hydrophobicity of the surface tends to increase protein adsorption, 75 but intermediate values of surface hydrophobicity are optimal for cell adhesion. 76
A comparison of blending, covalent linkage, and physical adsorption of laminin on electrospun fibers was performed by Koh and associates 77 to determine which method improved neurite extension, and electrospun fibers blended with laminin were most likely to induce long neurite extension. Thus, specific concentrations of proteins or peptides are necessary to improve the rate of neurite extension compared with unmodified fiber controls. 78
A protein commonly incorporated into electrospun fibers is collagen. Polymer and collagen blends or collagen alone improves neuron adhesion/viability. Generally, collagen inclusion within polymers reduces neurite extension when neurite extension is compared between polymer fiber only controls and collagen containing fibers. A PLLA-PCL blend electrospun with collagen I and III created fibers with diameters ranging from 196–288 nm. The inclusion of collagen improved C17.2 neural stem cell viability, but it was unclear how collagen blends of PCL affected neurite extension. 79 Schnell and colleagues 42 observed neurites from DRG explants were shorter on collagen/PCL scaffolds when compared with neurite outgrowth on PCL alone. 42
The reasons why collagen inclusion does not promote long neurite extension are unknown. Collagen is denatured by electrospinning, however. 72 Thus, it is possible that denatured collagen may be incapable of promoting the extension of neurites. A study by Liu and coworkers 60 placed collagen I electrospun fibers within an in vivo rat model of SCI. Unexpectedly, the fibrous scaffolds did not induce axonal extension into or onto the electrospun collagen scaffolds, which is a different result to the significant amount of axonal extension observed when electrospun PLLA fibers were placed into the rat spinal cord. 45 While it is difficult to directly compare these studies because of the major differences in fiber diameter in addition to the different materials, the lack of axonal extension into the scaffold may be because of the selection of collagen. 80
Another protein incorporated into electrospun fibers is the basement membrane glycoprotein laminin. As mentioned previously, Koh and colleagues 77 analyzed neurite extension on fibers where laminin was adsorbed to fibers, electrospun into, or covalently bound to the fiber surface. The study by Koh and colleagues 77 proved laminin inclusion in the electrospinning solution induced the longest extension of neurites.
In another study, 81 investigators electrospun pure laminin into fibers and cultured neurons on them. The length of neurites on these fibers, however, was not compared with neurite extension on other types of fibers. In a follow-up study, 61 blends of PCL and laminin (10% weight laminin) had the same effect on maximum neurite extension from mouse DRG cultured on electrospun fibers with a higher percentage of laminin (range 10–100% laminin). 61
Another method to modify the surface chemistry of fibers is through plasma treatment. Oxygen plasma treatment adds oxygen-containing molecules to the fiber surface, which decreases hydrophobicity. Corey and coworkers 82 sought to improve adhesion of primary rat sensory DRG or motor neurons through plasma treatment of aligned 2 μm diameter PLLA fibers. Interestingly, plasma treatment did not improve motor neuron survival when compared with motor neurons cultured onto a nonfibrous control. Further, motor neuron survival was statistically higher on untreated electrospun fibers. From these findings, Corey and colleagues 82 concluded that plasma treatment may restrict neuronal adhesion possibly because of electrostatic repulsion between the cell membrane and the negatively charged surface imparted by plasma treatment.
Similar findings were recently reported from our laboratory when chick DRG extended shorter neurites when cultured on oxygen plasma treated electrospun PLLA fibers. 83 In our study, different surface chemistries were covalently linked to the surface of the electrospun fibers to determine how different chemistries changed the hydrophilicity and ability to guide neurite extension. Although we found that all surface modifications resulted in a similar degree of hydrophilic character of the scaffolds, all of the treatments resulted in a decrease in neurite extension except the group that was modified with RGD, the only group with a biologically functional component. In these studies, neurons were cultured in media free of significant amounts of serum. Thus, the reduced neurite extension observed on charged surfaces is likely because of the presence of surface chemistry and not from differential adhesion of protein.
While plasma treatment may not directly improve neural adhesion or neurite extension by itself, the presence of oxygen species can provide a binding site for proteins. PLLA fibers with diameters ranging from 100 to 500 nm were plasma treated and functionalized with laminin and bFGF using a di-amino-poly(ethylene glycol) (di-NH2-PEG) linker covalently attached to carboxyl groups on the surface of the fibers using EDC-NHS chemistry. The tethered growth factors significantly improved the length and density of neurites extending from rat DRG. 84
Tethering the neural adhesion ligand L1 to the fiber surface enhanced both functionality in primary neuronal cells and differentiation of human embryonic stem cell derived neural progenitor cells (hESC-NPC) into GABAergic neural lineages. 85 L1 presents an RGD sequence that binds preferentially to neurons instead of astrocytes and oligodendrocytes. The selection of the L1 molecule is appropriate for SCI applications because L1 binds to α5β1 and αvβ3 integrins on the neural growth cone and improves functional recovery and axonal regeneration after SCI. Cherry and associates 85 electrospun a 1.25 μm tyrosine derived polycarbonate polymer and functionalized the fibers with L1. L1 ligand orientation on the fiber surface was improved using a tether that was comprised of the Fc receptor and protein A, and this combined method improved neurite extension.
One material aspect that has not received a significant amount of attention is how different surface modifications of electrospun fibers might degrade over time. This is an especially important topic considering the work from the group at Case Western Reserve with PNGs, where months of recovery time are required before functional recovery is observed. 7,8
Recent work from our laboratory suggests that some types of surface modifications may be very transient, lasting less than a few days. 83 The only electrospun fiber group modified with a peptide (RGD), however, appeared to be stable with no noticeable signs of degradation, so peptides and proteins that are linked to the surface may not have as transient a presence at the surface of electrospun fibers. Still, further work must be performed to better understand how surfaces can be modified to improve neurite extension, and an analysis of the stability of the surface modification should be included considering the potential length of time required to grow neurites across a scaffold in vivo.
Drug and protein delivery
While the previous sections have detailed the importance of fiber geometry (alignment and diameter) or surface chemistry with regard to neurite guidance, electrospun fibers are also capable of delivering therapeutic agents to stimulate neural regeneration or reduce the secondary injury response. The simplest method of adding a therapeutic agent to electrospun fibers is by direct addition of the agent to a solution before electrospinning. The small fiber diameters generate a high surface area to volume ratio, allowing for a sustained release profile via diffusion in slowly degrading polymers such as high molecular weight PLLA. 86 –88 The ability to release therapies from electrospun fibers locally enables the sustained release of therapies that otherwise might have difficulty reaching the injury site if the therapy were delivered systemically.
The first article to incorporate a therapeutic agent into electrospun fibers for nerve regeneration was a study presented by Chew and colleagues 73 in 2005. Chew and colleagues 73 loaded human β-nerve growth factor (NGF) into electrospun fibers. NGF was incorporated into an electrospinning solution containing a copolymer of PCL and poly(ethyl ethylene phosphate) (PCLEEP), and the release rate of NGF was sustained over a 90-day period. Chew tested the bioactivity of the NGF released from PCLEEP fibers by adding the released NGF to PC12 cells. The NGF released from the fibers lost some of its ability to induce neurite extension from PC12 cells. To improve NGF bioactivity, polymer and NGF were mixed with a surfactant. The inclusion of the surfactant improved NGF bioactivity in the presence of PC12 qualitatively, but the neurite extension results were not quantified. 89
A later study by Valmikinathan and colleagues 90 included bovine serum albumin (BSA) in an electrospinning solution containing NGF and PCL. BSA inclusion increased the total amount of NGF released from fibers and improved the bioactivity of the released NGF as shown when released NGF was cultured with PC12 cells inducing them to form neurites. Another article by Chew and coworkers 91 demonstrated the ability to release glial derived neurotrophic factor (GDNF) from PCLEEP. Although no bioactivity experiment was conducted, fibers loaded with GDNF induced recovery in animals with peripheral nerve injury compared with the controls with no GDNF. These studies demonstrate the ability of electrospun fibers to release proteins, but these studies also highlight the importance of protecting proteins from the large electrical fields required for the electrospinning process.
Other studies present strategies for delivery of proteins from electrospun fibers. Proteins (such as collagen) are denatured by the electrospinning process, 72 potentially because the organic solvents used for electrospinning can cause the protein to denature. In addition, fibers undergo intense stretching because of violent motion of the electrospinning jet, and the intense stretching may denature proteins.
One potential solution to maintain protein activity may be to covalently bind proteins to the fiber surface. Chew and colleagues 92 used microbial transglutaminase to covalently couple either ChABC or neurotrophin-3 (NT3) to the surface of electrospun collagen fibers. Collagen in the supernatant linked to NT3 was less capable than soluble NT3 at promoting neurite extension from embryonic day 15 rat DRG. When DRG were placed on fibers with covalently attached NT3, however, there were no differences in the length of neurite extension when comparing neurite outgrowth from DRG placed on plain collagen scaffolds with the addition of soluble NT3. In addition, collagen fibers covalently coupled to heparin or ChABC were more apt to maintain ChABC's ability to digest decorin over a 32-day period compared with soluble heparin.
Thus, post-fabrication surface inclusion of protein may maintain a protein's function better than mixing the protein in with the polymer and solvent before electrospinning. Very few studies have investigated the release of proteins from fibers for nerve regeneration applications, so more work is needed to develop new approaches to protect protein and better understand the mechanisms by which proteins diffuse out of the fibers.
Rather than delivering large proteins, some groups have delivered small organic molecules to improve neurite extension. Delivery of small organic molecules has a number of advantages over the release of protein. There is less of a concern that organic molecules will lose their bioactivity as a result of the electrospinning process. Further, the size of small organic molecules permits them to diffuse out of slowly degrading polymers at a much higher rate than proteins. This attribute is important, because slowly degrading polymers are of greater interest in nerve guidance because of the long periods required for functional regeneration in animal models of SCI (3–6 months). 6 –8
One of the first small organic molecules released from electrospun fibers for spinal cord repair was the antimetabolite 6-aminonicotinamide (6AN). 6AN is known to inhibit astrocyte metabolism at low levels while having a lesser effect on neurons. 93 –95 Schaub and Gilbert 96 released 6AN from electrospun PLLA, and the released 6AN was found to inhibit astrocyte viability without interfering with neurite extension at a concentration of 10% of the weight of the polymer. The intention was to try to inhibit astrocyte viability to reduce the negative effects from astrocyte reactivity after SCI, but no in vivo work was performed.
A more in-depth study was performed using rolipram by Downing and associates, 97 where rolipram was released from electrospun PLLA and placed in a C5 hemisection rat model of SCI. The study by Downing and associates 97 found that animals that received electrospun fibers that released a low dose of rolipram (∼3 μg/cm2 over 12 days) had significantly improved functional recovery after injury over a control group with injury alone. Interestingly, the treatment group that received electrospun fibers that released a low dose of rolipram had significantly improved motor function over another group that included electrospun fibers that released a large amount of rolipram (∼60 μg/cm2 over 12 days). This study demonstrates the ability to easily tune the amount of drug released from electrospun fibers and the ability to release therapeutic levels of drug.
In addition to the concern of maintaining protein bioactivity, if the protein or organic molecule is incorporated within the electrospun fiber, it is also important to note that protein incorporation or inclusion of small molecular weight therapeutic agents alters fiber geometry. Because fiber diameter is known to alter cellular function, fiber diameter must be maintained when performing cell culture experiments on fibers that release therapeutic agents. In general, inclusion of protein in an electrospinning solution results in electrospun fibers with drastically smaller fiber diameters compared with electrospinning solutions without protein. 73,89,98 The decreased fiber diameter may be attributed to the large charge character of the protein.
Schaub and Gilbert, 97 however, found a decreased fiber diameter even when using 6AN, a small organic molecule. To compensate, Schaub and Gilbert 96 reduced the concentration of PLLA fibers with no 6AN to obtain fibers with similar fiber diameter. The change in fiber diameter when protein or therapeutic agent is added to an electrospinning solution can interfere with a proper interpretation of the results if the fiber diameter is not compensated for, because either a small fiber diameter or the release of a therapeutic agent may be the source of any observed changes in neurite extension.
Electrospun Fibers as a Tool to Understand the CNS and SCI
Although the focus of this review thus far has detailed the use of electrospun fibers to guide axons after SCI, a new area of biological research is emerging that uses electrospun fibers to understand the behavior of CNS cells in the presence of topography. As stated in the introduction, dissociated DRG placed in the corpus callosum extended neurites along the length of white matter tracts. 99
Neurite extension, however, could be attributed to any number of factors, including trophic or structural cues. Electrospun fibers provide the opportunity to reduce the complexity of the system by generation of synthetic structures of similar dimension to axons because aligned fibers can be fabricated with diameters similar to those of axons found in the CNS (myelinated axons range in diameter from 0.2 μm to more than 16 μm). 100
In the absence of serum, which contains a complex assortment of proteins, neurites extend along the length of electrospun PLLA fibers. 82 Neurite guidance by electrospun fibers in the absence of other cells or protein gradients is a powerful indication that the physical, isotropic organization of white matter tracts plays a significant role in guiding axonal extension. In addition, simple experiments on electrospun fibers can help elucidate how biological factors, such as fibronectin, contribute to axonal guidance on these three dimensional structures. 101
A very recent discovery from the laboratories of the University of California, San Francisco, and the University of Michigan demonstrated that oligodendrocytes myelinate electrospun fibers in a diameter dependent manner, and that oligodendrocyte progenitors begin electrospun fiber ensheathment before differentiation. 102,103 This discovery effectively decoupled the molecular characteristics from the physical characteristics of an axon to demonstrate that the physical dimension of an axon is a critical component to myelination.
Myelination is a critical component in the action potential conduction velocity of neurons and is an important factor in restoring full functional recovery. In addition to being a useful tool for understanding oligodendrocyte myelination, it also gives further credence to a previous study that found primary neural stem cells differentiate into oligodendrocytes or neurons in response to electrospun fiber diameter. 104 These discoveries demonstrate how electrospun fibers can be used to better understand basic biological processes involved in the CNS, but also may prove useful for SCI repair because neural stem cells are currently being pursued as a treatment option. 105
In addition to oligodendrocytes, astrocytes have also been found to respond uniquely to electrospun fibers versus a flat surface. Zuidema and coworkers 106 demonstrated how astrocytes cultured on electrospun fibers produce more GLT-1 (a glutamate transport protein) when cultured on aligned or randomly oriented electrospun PLLA fibers compared with astrocytes cultured on a flat surface of PLLA. The significance of this finding is that GLT-1 is found to be down regulated after SCI 107 and contributes to neuronal excitotoxicity. 108
The results of the study from Zuidema and associates 106 suggest that the microscale, axon-like structure of the fibers contributes to the expression of GLT-1. Taking the results of the astrocyte study and the oligodendrocyte data from the preceding paragraph, the axon-like structure of electrospun fibers may be a more suitable surface for in vitro culture of glial cells because these cell types appear to behave more like glial cells in native tissue. Therefore, electrospun fibers may not simply be an approach to repairing SCI, but also a tool to better study glial cells in vitro.
In addition to better understanding the biology of neurons and other cell types in the CNS, electrospun fibers may also prove useful in understanding the mechanisms responsible for failure of axons to regenerate into a SCI lesion. Although the glial scar is believed to be a chemical and physical barrier to axonal extension, 109 –111 the extent of glial scarring in humans is marginal compared with rodents, 112 and some researchers suggest the mesenchymal scar plays a larger role in the failure of regeneration after SCI. 80,113
One study hypothesized that the orientation of collagen fibrils in the mesenchymal scar may contribute to the scar's inhibitory nature, 114 while other studies have demonstrated how certain collagens inhibit axonal growth. 80,115 Electrospun fibers have supported these ideas by demonstrating that neurites extend along electrospun collagen fibers, 57,116,117 but fibers containing collagen type I tend to have shorter neurites than neurites from neurons grown on biologically benign synthetic polymers. 42 These results suggest that collagen fibril organization within the fibrous mesenchymal scar may act as a barrier to nerve growth across the site of injury because the collagen fibrils are oriented parallel to the edge of the scar, which is perpendicular to the direction needed for nerve regeneration. Thus, electrospun fibers may aid in understanding whether components in the injured region are biologically or structurally inhibitory.
Conclusions
Although electrospun fibers have only been applied to neural applications for a decade, they have contributed to the understanding of how geometric structure may guide axonal extension. The initial in vitro and in vivo studies show that optimization of electrospun fibers can improve axonal extension and demonstrate the promise that electrospun fibers can be used as synthetic nerve guidance scaffolds. In addition to the fibers acting as a physical guide, further utility is found in the ability to use a wide range of materials and inclusion of therapeutic agents that can be released over time.
There is a great need for additional work to optimize fiber diameter, material properties, and release characteristics specifically for SCI, however. In addition to using electrospun fibers as a repair strategy, electrospun fibers may also prove a useful tool in understanding biological processes in the healthy and injured CNS.
Footnotes
Acknowledgments
The authors acknowledge grant support to RJG provided by the National Science Foundation (CAREER grant 11050125), National Institutes of Health (R01 NS092754), and the New York Spinal Cord Injury Research Trust (Contract # C030239). Nicholas Schaub was supported by the Ajit Prabhu '98 Fellowship.
Author Disclosure Statement
No competing financial interests exist.