
Abstract
The utility of in vitro models of traumatic brain injury (TBI) depends on their ability to recapitulate the in vivo TBI cascade. In this study, we used a genome-wide approach to compare changes in gene expression at several time points post-injury in both an in vitro model and an in vivo model of TBI. We found a total of 2073 differentially expressed genes in our in vitro model and 877 differentially expressed genes in our in vivo model when compared to noninjured controls. We found a strong correlation in gene expression changes between the two models (r = 0.69), providing confidence that the in vitro model represented at least part of the in vivo injury cascade. From these data, we searched for genes with significant changes in expression over time (analysis of covariance) and identified sorting protein-related receptor with A-type repeats (SORLA). SORLA directs amyloid precursor protein to the recycling pathway by direct binding and away from amyloid-beta producing enzymes. Mutations of SORLA have been linked to Alzheimer's disease (AD). We confirmed downregulation of SORLA expression in organotypic hippocampal slice cultures by immunohistochemistry and Western blotting and present preliminary data from human tissue that is consistent with these experimental results. Together, these data suggest that the in vitro model of TBI used in this study strongly recapitulates the in vivo TBI pathobiology and is well suited for future mechanistic or therapeutic studies. The data also suggest the possible involvement of SORLA in the post-traumatic cascade linking TBI to AD.
Introduction
E
The primary mechanical event sets in motion a myriad of secondary injury cascades that cause delayed cell death that may continue for days or longer. 9,10 Effective clinical treatments for TBI have been elusive, perhaps because of this complex pathophysiology. A better understanding of the secondary mechanisms leading to delayed cell death and dysfunction post-TBI may support the discovery and development of novel and clinically viable treatments. To this end, several in vivo and in vitro models have been developed to investigate TBI. 11,12 Both in vitro and in vivo models have respective strengths and weaknesses, but ultimately the utility of in vitro models depends on their ability to reproduce the in vivo TBI cascade. 12
The primary motivation of this study was to determine the degree of transcriptomic similarity between an in vivo and an in vitro model of TBI. Global gene expression changes likely coincide with the activation of different molecular pathways as the secondary injury process unfolds. Therefore, a high degree of correlation between the models would support the utility of in vitro models as research tools to complement in vivo models.
Utilizing DNA/RNA microarrays, it is possible to quantify global changes in gene expression. 13 This method eliminates the need to choose a specific gene of interest and produce the necessary tools (e.g., primers) for its study, but instead allows for thousands of genes to be analyzed simultaneously. From these data, entire physiological pathways can be studied, which may lead to further insight into injury mechanisms.
Previous studies of TBI have shed light on new genes and pathways of interest, but many of these studies focused on a single injury model or a single time point post-injury. 14 –16 Because of the complex nature of the post-traumatic pathology, temporal changes in gene expression may provide more insight than examination of a single time point. For example, temporal changes in gene expression have been studied for over a decade and have been integral in understanding physiological changes during circadian rhythm cycles. 17 –20 A temporal analysis could provide insight to the progression of a disease or pathology and could identify novel targets for therapies that would be overlooked with analysis at a single time point.
Our in vitro model, which has previously been characterized, utilized organotypic hippocampal slice cultures (OHSCs) that were subjected to an equibiaxial deformation, which was verified by high-speed video analysis. 12,21 –24 To serve as a comparison to the in vitro data, a group of animals were subjected to a closed-head injury using a pneumatically driven piston. 25 –27 Changes in gene expression between the two models were highly correlated at similar time points post-injury. From these data, we identified a transmembrane protein SORLA (sorting protein-related receptor with A-type repeats) that was significantly reduced post-TBI and confirmed its reduced expression with both Western blotting and immunohistochemistry (IHC) in our in vitro model. Interestingly, SORLA has been implicated in Alzheimer's disease (AD), and mutations that reduce SORLA's ability to function properly may lead to AD by decreasing amyloid-beta (Aβ) clearance, possibly linking TBI as a risk factor for AD. 28,29 In conclusion, our study confirms a strong correlation of the pathobiology between our in vitro model and that of an in vivo model, further supporting its utility as a research tool for the discovery of potential therapeutic targets post-TBI.
Methods
Organotypic hippocampal slice cultures
Animal procedures to generate OHSCs were approved by Columbia University's Institutional Animal Care and Use Committee (IACUC). OHSCs were cultured as previously described.
30
–32
Briefly, Sprague-Dawley rat pups (post-natal day [PND] 8–10) were rapidly decapitated, the hippocampus excised, and placed in ice-cold Gey's balanced salt solution (Life Technologies, Grand Island, NY) supplemented with
In vitro model of traumatic brain injury
After 10 DIV, OHSCs cultured on silicone membranes were subjected to a moderate mechanical injury. The injury was induced by stretching the underlying silicone substrate to a predetermined strain at a pre-determined strain rate to produce the desired tissue injury under motion control. Our well-established model produces a highly accurate and reproducible injury to OHSCs. 12,23,32 Tissue deformation was verified by image analysis of high-speed video (MotionPro; Redlake, Pasadena, CA) at 1000 frames per second. 21 Lagrangian strain of the tissue was determined by calculating the deformation gradient tensor by locating fiduciary markers on the tissue slice before and at maximal stretch using custom MATLAB software (The MathWorks, Inc., Natick, MA) scripts. 21
In vivo model of traumatic brain injury
Animal procedures for closed-head TBI were approved by Drexel University's IACUC and were in compliance with the Guide for the Care and Use of Animals. Brain injuries were induced using a pneumatically driven controlled cortical impact device (AmScien Incorporated, Richmond, VA) as described by Raghupathi and colleagues. 27 Briefly, rat pups (PND11) were anesthetized with isofluorane (2%) using a nose cone, and once a loss of a tail-pinch reflex was observed, a midline incision was made to expose the skull. The periosteum was reflected, the animal was placed in a restrainer, and the head was supported by a soft foam pad to make it level with the body. The restrainer was positioned under the cortical impact device, the nose cone was removed, and the zero-point for the indenter was made on the skull over the left parietal cortex, midway between the lambda and bregma sutures. The exposed skull was subjected to a 5-mm-diameter piston impact of 3.0 mm in depth, 5 m/s velocity, and 100 ms in duration for moderate injury. Sham-injured animals underwent the same procedures, except that the piston was not activated. Total time from initiation of anesthesia to removal of the nose cone preceding zeroing the impactor tip was typically 5–6 min. After recovery from injury/anesthesia, animals were reanesthetized with isofluorane for the scalp to be sutured, and pups were returned to the dam. Animals were placed on a heating pad kept at 37°C to maintain body temperature throughout the procedures and recovery.
Affymetrix gene chips
OHSC samples, including time-matched controls, were collected after the in vitro model of TBI at 1, 6, 12, and 24 h post-injury. Whole hippocampal samples, including time-matched controls, were collected after the in vivo model of TBI at 1, 6, and 24 h post-injury. Tissue was collected, flash frozen, and stored at −80°C until total RNA was isolated using an RNeasy Mini Kit (Qiagen, Valencia, CA), following the manufacturer's protocols. Quality of RNA was assessed using a NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA). Purified RNA was used to synthesize complementary (cDNA) using the HT One-Cycle cDNA Synthesis Kit (Affymetrix, Santa Clara, CA), following the manufacturer's protocols. Amplified complementary RNA cRNA was produced from the cDNA template using the 3′ IVT Labeling Kit (Affymetrix). The cRNA was then hybridized to the Affymetrix Rat 230 2.0 chips, washed, and scanned at the Columbia Genome Center according to the manufacturer's protocols.
Microarray data analysis
Using custom MATLAB scripts, each array was normalized for variations in background using the robust multi-array average procedure. 33 After performing a Student's t-test between experimental groups on an individual gene basis, p values were adjusted for false discovery rate (FDR) using Storey-Tibshirani's method to improve confidence in p values. 34 An FDR-corrected p value <0.05 and a 2-fold biological change in expression were chosen as criteria for identifying genes whose expression was altered both statistically and biologically. 35 Analysis of covariance (ANCOVA) was used to determine whether gene expression over time was significantly different between experimentally injured groups and noninjured controls. An ANCOVA p value <0.05 was considered significant. To enrich for the most significantly changed genes in our data set for further analysis, we focused on genes that were significant in both the Student's t-test and ANCOVA.
To determine the correlation between the two injury models, Pearson's correlation coefficient was determined for the 323 genes found to be significant in both the in vitro and in vivo models by the Student's t-test.
Using Database for Annotation, Visualization and Integrated Discovery (DAVID) Bioinformatics Resources provided by the National Institutes of Health (NIH), each time point was analyzed for the most enriched Gene Ontology (GO) terms. 36 We chose to focus on biological process terms and have excluded cellular component and molecular function GO terms from analysis. Specifically, GO FAT biological process terms, as described by DAVID, were used for analysis. The GO FAT subset excludes the broadest GO terms, so that they do not overshadow more-specific terms.
Western blots
Additional samples were collected 48 h post-injury to confirm the transcriptional results at the protein level. This time point was chosen to allow for changes in messenger RNA expression to be translated to protein expression. For in vitro injured and uninjured controls, nine slices were collected for protein extraction. Slices were rinsed twice with ice-cold phosphate-buffered saline and immediately placed in lysis buffer A (40 mM of HEPES, 120 mM of NaCl, 1 mM of ethylenediaminetetraacetic acid, 1% Triton X-100, 10 mM of sodium pyrophosphate, 50 mM of sodium fluoride, 0.5 mM of sodium orthovanadate, and 10 mM of β-glycerophosphate; Sigma-Aldrich). Samples were sonicated (Sonicator 3000; Misonix, Farmingdale, NY), incubated on ice, and then centrifuged to remove cell debris. Approximately 50 μg of protein per sample was loaded in a 4–12% Bis-Tris gel (Life Technologies), and proteins were separated by electrophoresis (150 V, 1 h). Proteins were then transferred to a nitrocellulose membrane (Life Technologies) using a semidry apparatus (Thermo Fisher Scientific). After transfer, the membrane was blocked in Tris-buffered saline (TBS; pH 7.4) with 5% bovine serum albumin (BSA) for 2 h. Membranes were incubated overnight at 4°C with primary antibodies (Anti-SORL1 [S9200; Sigma-Aldrich], anti-β-tubulin to serve as an internal control [32-2600; Life Technologies]) in TBS-T (0.1% Tween-20; pH 7.4) and 5% BSA. After primary antibody incubation, membranes were washed 3 × 10 min in TBS-T. For detection, membranes were labeled with a corresponding secondary antibody (goat/anti-rabbit Alexa Fluor 546 or goat/anti-mouse Alexa Fluor 647; Life Technologies). Fluorescence was detected using a CRi Maestro 2 Imaging System (PerkinElmer, Akron, OH). Grayscale images were analyzed in ImageJ (NIH, Bethesda, MD) using the gel analysis function. 37 SORLA band intensities were normalized by the intensity of the β-tubulin band.
Patient samples
Autopsy case material was obtained from the New York City Office of Chief Medical Examiner (OCME; New York, NY). According to the policies of the New York City OCME and New York University School of Medicine, this research did not require institutional review board (IRB) approval; however, it was submitted to and approved by the OCME legal office to insure that it complies with OCME policies and did not require IRB approval. Two types of subjects who died of TBI were sought: those that died immediately from the TBI and those that survived for at least 12 h post-TBI. The immediate-death cases served as controls for comparison of changes in post-traumatic protein expression in the delayed-death cases. Blocks of fixed brain tissue from subjects with TBI were embedded in paraffin.
Immunohistochemistry
All steps were performed with a Ventana Benchmark Ultra automated staining instrument (Ventana Medical Systems, Tucson, AZ). Formalin-fixed, paraffin-embedded tissue samples from either human (at indicated time points) or rat OHSCs (48 h post-injury) were cut at 8 or 6 um, respectively. Using the automated Ventana equipment, paraffin was removed at 72°C, and antigen retrieval was achieved by incubation with CC1 reagent (Ventana Medical Systems) at 95°C. After antigen retrieval, slides were incubated with anti-SORL1 (S9200, 1:1000; Sigma-Aldrich) for 32 min before being washed. Tissue nuclei were stained using hematoxylin. Slides were then dehydrated using gradient alcohols and cover-slipped. Adjacent sections were stained with Luxol fast blue and hematoxylin and eosin (LHE) for routine pathological analysis.
Quantification of staining in human tissue
Stained sections from autopsy cases were analyzed semiquantitatively by a blinded observer. For SORLA IHC, a 4-point rating scale (0, +, ++, and +++) was devised to correspond to the intensity of staining (none, weak, moderate, or strong, respectively). A similar scheme (0, +, ++, and +++) for the LHE-stained sections (none, rare, occasional, or frequent, respectively) was devised to quantify pathological findings, including shrunken neurons, vacuolization, neuronal loss, and dark neurons.
Results
Comparison of in vitro and in vivo traumatic brain injury models
Across all time points, a total of 2073 unique genes were differentially expressed in at least one time point after in vitro TBI, and a total of 877 genes were differentially expressed in at least one time point after in vivo TBI compared to uninjured controls (Supplementary Data) (see online supplementary material at
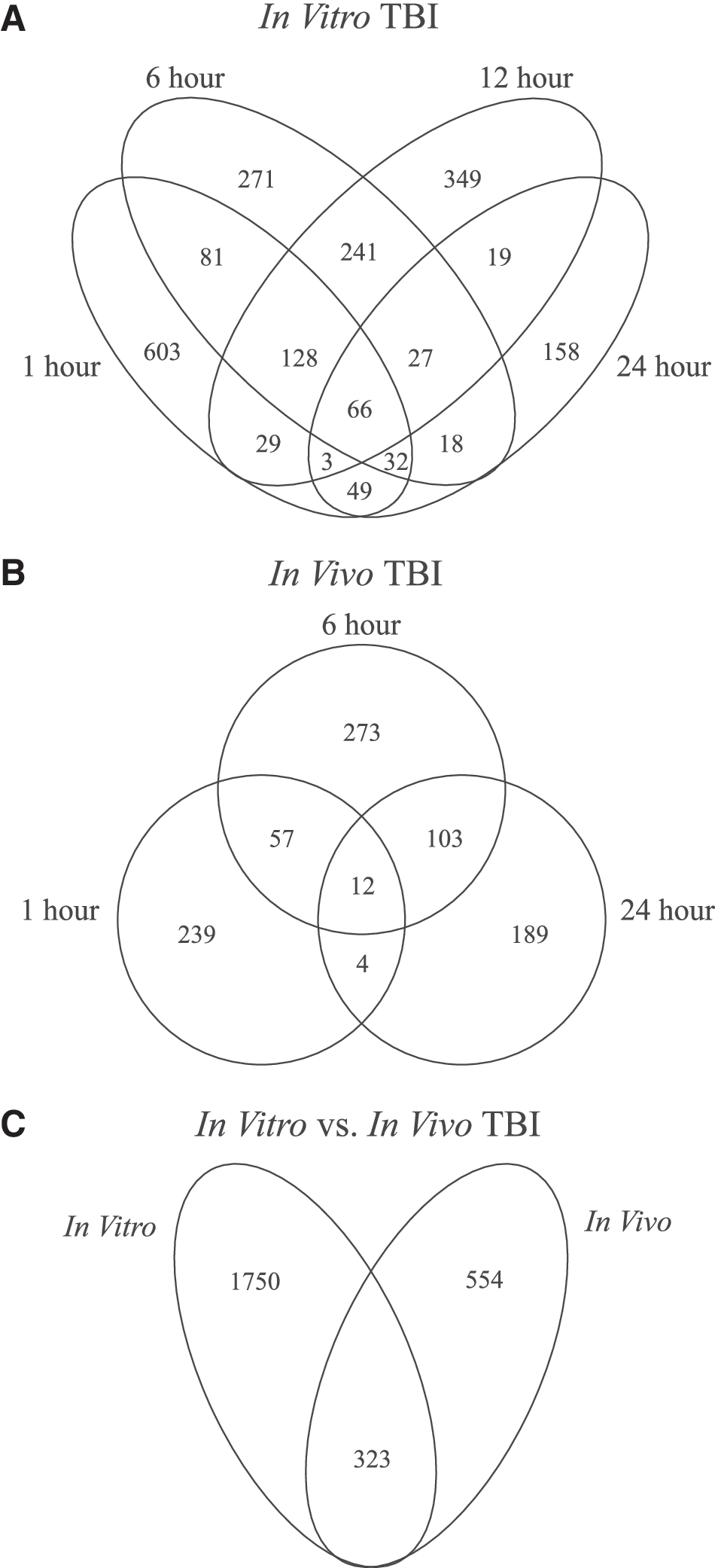
Overview of significant changes in gene expression post-TBI. Venn diagrams show the number of overlapping genes whose expression was significantly altered compared to time-matched controls for (
We next investigated how correlated gene expression was between the two models of TBI for the 323 differentially expressed genes common to both the in vivo and in vitro models. The comparison of fold changes is graphed in Figure 2. There was a strong correlation in gene expression between the in vitro and in vivo models at matching time points (1 h, r = 0.54; 6 h, r = 0.64; 24 h, r = 0.59). Within the in vitro injury group, there were very strong correlations between the 6- and 12-h time points (r = 0.89) and strong correlations between the 6- and 24-h time points (r = 0.67) and the 12- and 24-h time points (r = 0.62). This trend was similar for the in vivo injury between the 6- and 24-h time points (r = 0.85). A moderately negative correlation between the 1-h time point and all later time points within a given model (Fig. 2) suggests that an abundance of genes were initially differentially expressed post-injury and returned to baseline over time.
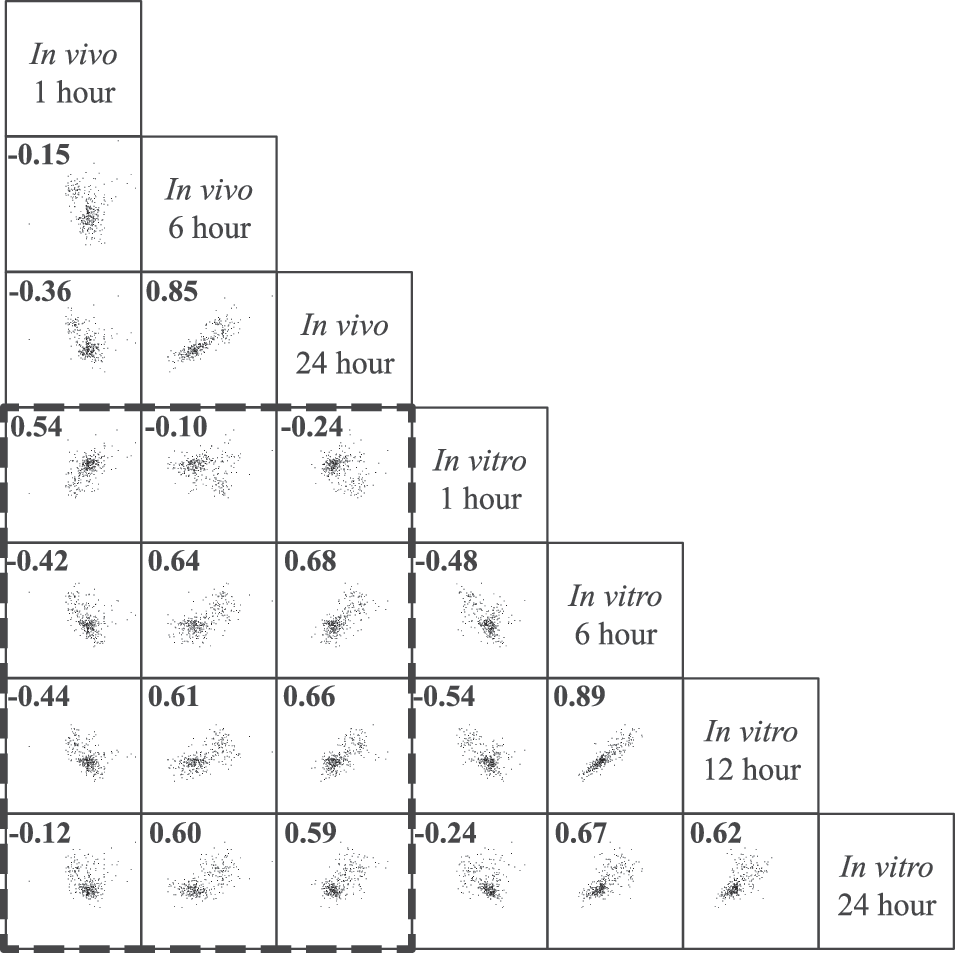
Correlation of gene expression changes post-TBI. Scatter plots of log2 fold changes of the 323 genes that were significantly and differentially expressed in both models. Pearson's correlation coefficients (r) are located in the top left corner. Axes represent log2 fold change from −5 to 5. Dashed line encloses in vitro versus in vivo comparisons. Both models were strongly correlated, particularly at matching time points.
For an overall comparison of our in vitro to in vivo TBI models, we combined the gene expression fold changes for the 37% of genes that were significantly regulated in both models at identical time points post-injury (1 h in vitro vs. 1 h in vivo, 6 h in vitro vs. 6 h in vivo, and 24 h in vitro vs. 24 h in vivo). Gene expression fold changes for the time-point–matched analysis had a combined Pearson correlation of r = 0.69. This strong correlation gives us confidence that our in vitro model is at least partially representative of in vivo TBI.
Gene ontology analysis
After analysis for the most significantly enriched GO terms in our data set, TBI-relevant GO terms were chosen, and the number of genes within a GO term for in vitro and in vivo injuries were graphed (Fig. 3). Several TBI-relevant GO terms contained significantly differentially expressed genes after both injury models, and further, many of the genes in these relevant groups were identical as indicated in Figure 3.
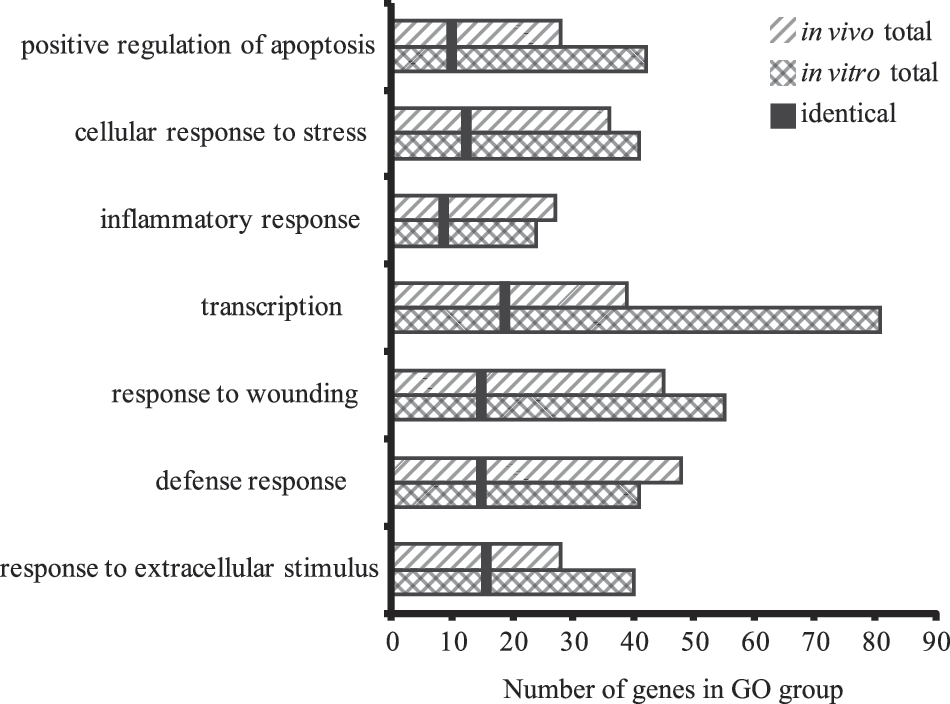
GO analysis of significantly altered gene expression post-TBI. The number of significantly altered genes from each model within the specified GO groups is graphed. The vertical black bar represents the number of identical genes that were significantly regulated in both models within the specified GO group. TBI-relevant GO groups were remarkably similar between the in vitro and in vivo models, and many identical genes were altered in both models. GO, Gene Ontology; TBI, traumatic brain injury.
Analysis of covariance of in vitro traumatic brain injury
The comparison of significant genes between the ANCOVA analysis and Student's t-test yielded 32 genes representing changes of particular interest. These genes are listed in Table 1. For further analysis, we decided to focus on genes that were not involved in cell adhesion, were predominantly expressed in the central nervous system (CNS), and were previously linked to diseases of the CNS. Following these guidelines we further validated SORLA, which has been linked to AD, with Western blot and IHC. 28,29
ANCOVA, analysis of covariance; NCBI, National Center for Biotechnology Information.
Protein expression of sorting protein-related receptor with A-type repeats
Although SORLA is widely expressed in the brain, it is predominately expressed in neurons of the hippocampus. 38 Cytoplasmic puncta were observed in the cornu ammonis 1 (CA1) pyramidal neurons of OHSCs of uninjured controls (Fig. 4A,B). Interestingly, at 48 h after mechanical injury of the OHSCs, these puncta were reduced or completely missing from the cytoplasm of pyramidal neurons (Figure 4 C,D). Injured regions also exhibited a vacuolar appearance, suggesting cell death. These data further support our gene chip data, corroborating reduced SORLA expression post-injury. To further confirm this claim, we measured protein expression of SORLA by Western blotting after in vitro TBI. We found that SORLA expression was significantly reduced by 44% compared to uninjured control OHSCs (p < 0.05; Fig. 4E,F).

SORLA expression after in vitro injury. Micrographs of SORLA staining in uninjured (
Next, we assessed changes in SORLA expression in the brains of human subjects after severe TBI. Sections of brain tissue from 8 TBI subjects were studied with details given in Table 2. Of these subjects, 5 died shortly post-injury, and 3 had prolonged survival (≥12 h). Semiquantitative results are summarized in Table 3. In the immediate deaths, a granular label was detected prominently within neurons, but the number and sizes of these granules varied considerably across brain regions. Neurons in CA1 often had single or scattered medium-sized cytoplasmic granules (∼1 μm in diameter; Fig. 5A). Within the dentate gyrus and entorhinal cortex, numerous small, finely granular staining was present that filled the cytoplasm (Fig. 5B,C). Neurons in the entorhinal cortex often had an additional single large, perinuclear granule (∼8 μm). A diffuse granular label of the cytoplasm was also detected occasionally within microglia. Staining in astroglia and oligodendroglia was not prominent. In the cases that died ≥12 h post-injury, the signal intensity of SORLA staining was less prominent (Fig. 5D); however, the significance of this is uncertain given the small sample size and the heterogeneity that is associated with human tissue. In the frontal white matter from 1 case, we found numerous SORLA-positive cellular processes (Fig. 5E,F). The significance of this finding is also unclear, but this is a brain region known to be vulnerable to diffuse axonal injury and may reflect secondary changes in axons. Together, these results open up the possibility that changes in SORLA expression may occur in human brain post-TBI.

SORLA expression after human TBI. Localization of SORLA in human post-mortem brain post-TBI. (
TBI, traumatic brain injury.
0 = none; + = weak/rare; ++ = moderate/occasional; +++ = strong/frequent.
SORLA, sorting protein-related receptor with A-type repeats; H+E, hematoxylin and eosin; IHC, immunohistochemistry; DG, dentate gyrus; CA1, cornu ammonis 1; Sub, subiculum; EC, entorhinal cortex; PG, parahippocampal gyrus.
Discussion
In vitro models of TBI have the advantage of higher throughput than in vivo methods, but their utility depends on their ability to replicate the in vivo injury cascade(s). To that end, this study compared the global changes in gene expression post-TBI in both an in vitro and an in vivo model. Time points were chosen to investigate the near-immediate cellular response (1 h), which may lead to downstream cascades at later time points (6 and 12 h) and potential stabilization and recovery (24 h). Although the effects from TBI, such as cell death, can persist for several hours or days after the initial injury, this study focused on the time frame post-TBI during which therapeutic interventions may be successful, and our data may point to novel targets for future clinical treatments. 9,10,39,40 The authors acknowledge that additional time points would be relevant and could further elucidate long-term pathological effects of TBI. Nonetheless, the current study revealed interesting and novel effects of TBI that were identified with an in vitro study and confirmed in vivo.
Some of the significant gene expression changes in our current study have been identified previously either in genome-wide or single-gene studies. 14 –16,41 –45 Di Pietro and colleagues, using the same in vitro model as ours, investigated mild (10%) and severe (50%) injuries in OHSCs at 24 h post-injury and found significant changes in expression of inflammatory and apoptotic genes, such as Cox-2, Cxcl2, and Mmp2. 16,46,47 However, only a single time point of 24 h was used for their analysis. Our data show many changes in gene regulation at 1 h post-injury that are not evident at 24 h. It is possible that, in their model, the severe injury may induce significant gene expression changes at an earlier time point than mild TBI, which would be overlooked by focusing on a single time point. Both Crack and colleagues and Poulsen and colleagues used an in vivo TBI model and analyzed gene expression changes at multiple time points post-injury. 15,48 However, no temporal analysis method was employed; instead analysis focused on gene expression changes at individual time points. In both studies, similar inflammatory genes were found to be differentially expressed. A reanalysis of their data sets, taking into account that temporal aspects could reveal interesting and statistically significant changes that would otherwise be missed. White and colleagues analyzed gene expression changes 24 h post-TBI in rats. 41 They compared ipsilateral to contralateral tissue and found significant changes in inflammatory genes. The researchers acknowledged the temporal limitation of their study and suggested that a larger study investigating many time points would be necessary to track changes in gene expression. To our knowledge, ours is the first study of global gene expression post-TBI that takes advantage of multiple time points (ANCOVA) to further enrich our conclusions and provide novel targets for future study.
Comparison of in vitro and in vivo traumatic brain injury models
Gene expression between the two models was strongly and positively correlated 1 h post-injury. Interestingly, these initial changes in gene expression were moderately negatively correlated with later changes in gene expression suggesting a transient burst of expression followed by a return to baseline similar to post-traumatic immediate early gene expression observed by others. 49 –52 This relationship was apparent in both the in vitro and in vivo models of our study. The strongest correlations within a model were between the in vitro 6- and 12-h time points and the in vivo 6- and 24-h time points. This trend extended to the cross-model comparison at the later time points (6, 12, and 24 h) as well. Biological fold changes between the in vitro and in vivo model at matched time points (1 h in vitro vs. 1 h in vivo, 6 h in vitro vs. 6 h in vivo, and 24 h in vitro vs. 24 h in vivo) had a strong correlation (r = 0.69). These data suggest that the post-injury changes in gene expression follow a similar trend and therefore the in vitro model is well representative of the in vivo model.
Gene oncology analysis
Several TBI-related GO terms were enriched post-injury in both models. Not surprisingly, several GO terms related to inflammation were enriched including response to wounding (GO:0009611), response to extracellular stimulus (GO:0009991), defense response (GO:0006952), cellular response to stress (GO:0033554), and inflammatory response (GO:0006954). These GO terms contain genes activated by damage to the organism. Response to wounding specifically encompasses genes that are activated directly by an external stimulus, whereas defense response is the reaction of a foreign body introduced to a cell from an attack. Genes from both models that fall within these GO categories included Cd14, Cox-2, Ptges, Cxcl2, and Ccl2. Ccl2 has been implicated in recruitment of macrophages post-TBI, and elevated levels can be measured for 10 days post-trauma in severe TBI patients. 53 Cox-2 is one of the most widely studied enzymes responsible for inflammation, and pain and is the target of most nonsteroidal anti-inflammatory drugs, such as ibuprofen. 54,55 Cox-2 has previously been targeted as a potential therapy for TBI and has shown promise in pre-clinical studies. 47
Secondary sequelae post-TBI can lead to delayed cell death for hours, days, or even years. 56,57 Genes within the GO category positive regulation of apoptosis (GO:0043065) were significantly altered in both injury models and could prove to be targets for therapeutic intervention. For instance, Wwox is an essential mediator of tumor necrosis factor alpha–induced apoptosis and is upregulated post-injury in our models of TBI. 58 Mmp2, which was significantly increased after closed-head TBI and is implicated in synapse loss, was also found to be upregulated in our study immediately post-injury. 59 Jnk2 plays an important role in neuronal cell death as well as astrocyte activation, and inhibitors to Jnk2 have shown promising pre-clinical results. 60 –63
The GO term transcription (GO:0006350) included several differentially expressed genes in our study that could have wide-ranging effects on cellular response post-injury. Some of these genes are well-known markers of injury, such as Atf3, and others are part of the transcription machinery, such as Polr2a, which forms the largest subunit of RNA polymerase II. 64 Perhaps more interestingly, some of the identified genes are directly associated with development and neurogenesis, including Notch-2 and Adnp, respectively. It has previously been reported that peptides based on Adnp can have direct, neuroprotective effects after closed-head injury in mice. 65 Adnp has also been shown to be vital to embryogenesis and neurogenesis and could have direct implications in neural repair post-injury. 66
Sorting protein-related receptor with A-type repeats downregulation after traumatic brain injury
By combining different statistical approaches (single-time-point t-tests and ANCOVA), 32 genes were identified as being of particular interest. From this enriched subset of genes, Sorla, which plays a key role in recycling amyloid precursor protein (APP) from the cell surface through the endocytic pathway, was identified. Accumulation of APP in the axon is a hallmark pathology of TBI observed in multiple animal models as well as clinically. 67 –72 In the pig model of inertial brain injury, accumulation of APP with BACE, PS-1, and activated caspases has been suggested to lead to increased Aβ processing, leading to formation of amyloid plaques. 73 Our results suggest another molecular contributor in the accumulation of Aβ post-TBI. Specifically, SORLA directs trafficking of APP into recycling pathways by direct binding. Common mutations of SORLA, which are predictive of AD, reduce the binding affinity of SORLA for APP, leaving APP to enter Aβ-generating compartments and accumulate. 28,29 It is speculated that SORLA, in this manner, is mechanistically involved in AD. Accumulating evidence from epidemiological studies implicates TBI as a possible risk factor in AD development. 74 Although the link between TBI and AD is controversial, extensive research continues to explore the relationship, if any, between injury and disease. At the forefront of this research is an apolipoprotein E (APOE) genotype (ɛ4) that results in higher Aβ deposition post-TBI. However, the link between the APOE ɛ4 genotype and TBI is inconclusive, and no other biological explanation has been well substantiated. In our model, Sorla was downregulated post-TBI, which may lead to a disruption in APP recycling leading to the accumulation of Aβ, suggesting a possible link to AD. Therefore, the identification of decreased Sorla expression post-injury was an interesting and potentially clinically relevant discovery from our in vitro model of TBI.
IHC staining of SORLA in uninjured OHSCs was similar to previous reports in both rat and human tissue. 38 Punctate staining was specific to neurons located at the base of the apical dendrites within structures hypothesized to be butrysomes (Fig. 4A,B), suggesting that SORLA expression was not altered during the process of culturing the OHSCs. Further confirming our microarray analysis, SORLA expression was considerably lower, and puncta were almost nonexistent, after in vitro TBI (Fig. 4C,D). Western blotting analysis of SORLA expression confirmed that SORLA was significantly downregulated after mechanical injury in our in vitro model. Preliminary results of human tissue show a similar response, although a larger sample size is necessary to confirm these changes. Further, human samples suitable for analysis by Western blot would strengthen our confidence that these changes are truly biologically significant. These results suggest a potential mechanism for Aβ accumulation post-injury and that SORLA may be a potential therapeutic target for post-TBI treatment, given that an abundance of Aβ can lead to neuronal cell death. 75 Further research to determine the injury threshold for SORLA reduction and temporal response are necessary to better understand the connection between TBI and SORLA changes. These data suggest the potential therapeutic benefit of targeting SORLA post-injury and may expose links between TBI and neurodegenerative diseases.
Although several genes were highly correlated in both models of TBI, there were many genes that did not overlap between the two models. It is possible that the severity of injury was not equal in both models, although moderate levels were specifically chosen. Further work will be required to determine the importance of these nonoverlapping genes. Although many inflammatory genes were upregulated in our in vitro model, the OHSCs are missing the systemic component of the inflammatory response, which could have major effects on the efficacy of any therapy tested in vitro. Other limitations of the in vitro model include a lack of a blood–brain barrier and circulatory system. We also opted to include more time points in our study at the expense of more arrays per experimental condition. Although the high correlation of genes is a promising result, a more extensive study with additional arrays would provide higher statistical confidence.
Utilizing microarrays, we compared our in vitro model of TBI to an in vivo model. We found many overlapping genes that were categorized in similar GO groups. The correlation of gene expression between models was high; of the 12 Pearson correlations made between the two models, seven had a strong positive correlation (r > 0.50). A remaining two had moderately negative correlations (r < −0.40) and were comparisons of the in vivo 1 h to the in vitro 6- and 12-h time points. These negative correlations could potentially represent immediate early genes responding to the initial mechanical injury, which then normalize at later time points, but this speculation would require further study to confirm. These data, taken together and with the previously mentioned limitations, provide confidence in using the in vitro model to represent some of the in vivo TBI cascade. This important conclusion supports the use of in vitro models such as ours for drug discovery and mechanistic studies, which may reduce the number of animals necessary for TBI research. Further, our study identified significant changes in Sorla expression, which previously has not been implicated in TBI research, and may provide a new connection between TBI and AD, as well as an avenue for therapeutic intervention.
Footnotes
Acknowledgments
This work was supported, in part, by NIH NINDS R03 NS054256 (BMIII), National Institute of Child Health and Development R01 HD 069163 (RR), a National Science Foundation Graduate Research Fellowship (to M.R.L.), and a Natural Sciences and Engineering Research Council of Canada postgraduate scholarship (to B.S.E.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.