
Abstract
Spinal cord injury (SCI) is a devastating condition affecting 270,000 people in the United States. The use of growth factors is a potential treatment for reducing secondary damage, promoting axon growth, and restoring some of the lost function post-SCI. Glial cell line-derived neurotrophic factor (GDNF) is an important growth factor, because it can affect both neurons and support cells. Here, we give an in-depth review of the previously published literature where GDNF was used to treat SCI. The effects of GDNF have been shown to decrease lesion size, improve allodynia, and regenerate axons in the central nervous system and peripheral nervous system. GDNF is necessary for early development, and lack of GDNF can lead to abnormal development of the autonomic nervous system or death. Exogenous administration of GDNF either before or immediately after SCI is most effective. Even though GDNF can be directly administered, genetically modified cells are often used as a delivery vehicle. Several different types of genetically modified cells have been used with varying success. Although GDNF is effective when used alone, it has been shown to be more effective when used in combination with other neurotrophic factors. Overall, GDNF significantly improved functional recovery, increased the number of sprouting neurons, reduced lesion size at the injury site, and had minimal adverse effects.
Introduction
I
One potential treatment that has been researched over the past two decades is glial cell line-derived neurotrophic factor (GDNF), a small protein that potently promotes the survival, function, and differentiation of many types of neurons. 20,21 Treatment with GDNF has been shown to be neuroprotective, provide trophic support to neurons, protect against harmful secondary inflammation, and it is useful in combination therapies. GDNF has been experimentally shown to decrease lesion volume, reduce allodynia, and improve functional recovery in rat and mouse models. GDNF is also being researched as potential treatment for other health issues, such as amyotrophic lateral sclerosis, 22 spinal muscular atrophies, Parkinson's disease (PD), 20,23 and many others. 24
In this article, we will review the structure and cellular action of GDNF, investigate the role of GDNF in the central nervous system (CNS), the rationale behind choosing GDNF as a potential therapy for treating SCI, and perform a comparison of the previous experiments that used GDNF post-SCI.
Structure and Signaling Action
Structure
GDNF family ligands (GFLs) consist of four neurotrophic factors: GDNF, Neurturin, 25 Artemin, 26 and Persephin, 27 with GDNF being the most potent. 28 –30 The GFLs have seven conserved cysteine residues with similar spacing, which make them distant members of the transforming growth factor (TGF)-β superfamily. 31,32 Although originally characterized as a growth factor promoting the survival of ventral midbrain dopaminergic neurons, 33 GDNF is now believed to have a potent survival effect on motor neurons and other neuronal subpopulations in the CNS and peripheral nervous system (PNS). 34
GDNF was initially identified and purified from the B49 cell line while searching for neurotrophic factors to promote the survival of dopaminergic neurons. 31 Two hundred eleven amino acids make up the inactive form of GDNF; pre-GDNF, which is later cleaved into the active form, 134 amino acids. GDNF was later characterized as a glycosylated homodimer connected by disulfide bond weighing 33–45 kDa. 31,35,36
Signaling mechanism
GDNF utilizes a multicomponent receptor complex consisting of Ret receptor tyrosine kinase, which specializes in transmembrane signaling, and glycosylphosphatidylinositol-anchored coreceptor GDNF family receptor alpha (GFRα). GFRα specializes in ligand binding, and together with GDNF, they aid in the proliferation, migration, and differentiation of neuronal cells. 36,37 Despite being the common signaling receptor for all the GFLs, Ret is unable to bind any of the GFLs without the presence of the GFRα coreceptor, which include four members (GFRα 1–4) responsible for determining ligand specificity (Fig. 1). 34 GFRαs can activate Ret signaling in a non-cell-autonomous fashion, presenting GFLs in trans to Ret-expressing cells. This led to the discovery of an alternative signaling receptor, which utilizes neural cell adhesion molecule (NCAM) for GFLs (Fig. 1). 34
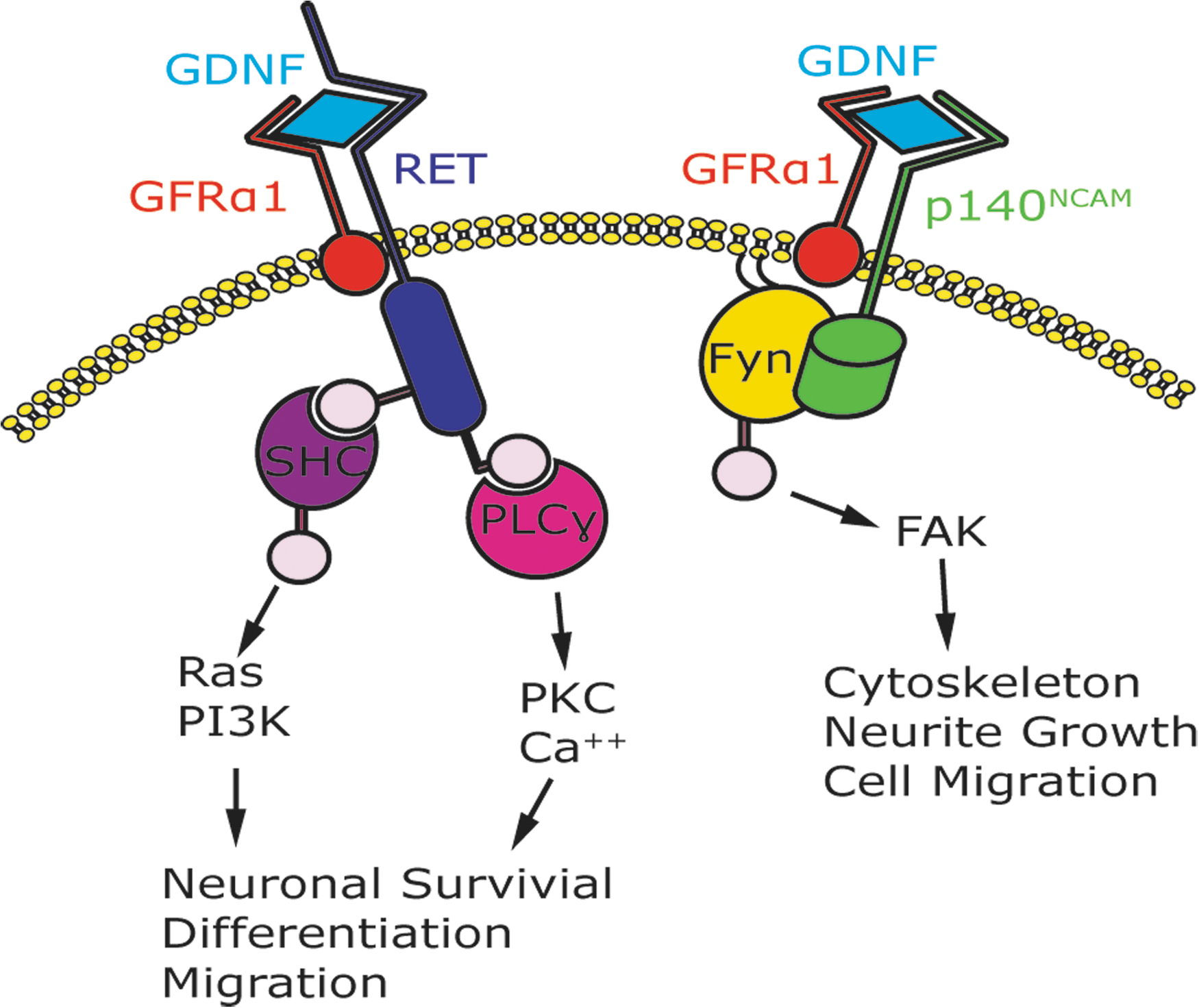
GDNF can signal through either the RET-independent or RET-dependent pathway. In the RET-dependent pathway, GDNF binds specifically to GFRα1. This complex brings two molecules of RET together, causing transphosphorylation of specific tyrosine residues 21 and intracellular signaling. Mediators, such as Shc, are soluble adaptors that will recruit RET to lipid rafts and trigger raft-specific signaling. 21 PI3-kinase and phospholipase (PLC-γ) are involved in enhancing neurotransmission. 21 The RET-dependent pathway has been shown to lead to neuronal survival, differentiation, and migration. In the RET-independent pathway, GDNF binds specifically to GFRα1 and with high affinity to p140NCAM, which activates the Src-like kinase, Fyn, and FAK. 51 The Ret-independent pathway has been shown to lead to cytoskeleton, neurite growth, and cell migration. FAK, focal adhesion kinase; GDNF, glial cell line-derived neurotrophic factor; GFRα1, GDNF family receptor alpha-1; RET, rearranged during transfection; PI3K, phosphoinositide 3-kinase; PKC, protein kinase C; FAK, focal adhesion kinase.
Recent studies suggest that, in intracellular communication, the extracellular domain of cell-surface receptors can act as either a transmitter or a receiver. GFLs function by signaling through a multicomponent receptor complex comprising of the transmembrane Ret tyrosine kinase and a member of a family of glycosylphosphatidylinositol-anchored cell-surface proteins, the GFRα1.36 38 GFRα1 resides on the plasma membrane of the nerve terminal. GFRα1 is released by neuronal cells, Schwann cells (SCs), and injured sciatic nerve. 39 All neurons, except motorneurons, require TGF-β to activate GFRα1 transport to the cell membrane. 33 Intracellular signaling is performed by Ret, whereas ligand binding is carried out by the GFRα. 40 The Ret tyrosine kinase was the first GDNF intracellular target identified. As we mentioned above, there is a Ret-independent signaling pathway, which activates GFRα1-associated Src kinase. GFRα has also been shown to use neural cell adhesion molecules, such as NCAM, as signaling receptors for GDNF family ligands (Fig. 1). 33
GFRα receptors may function in a non-cell-autonomous way to capture and concentrate diffusible GFLs from the extracellular space and then present these factors in trans to afferent Ret-expressing cells. 37,41 GDNF could activate Ret through either GFRα1 or GFRα2. In a motor-neuron–derived cell line expressing Ret and GFRαs, GDNF stimulated sustained activation of the Ras/extracellular signal-regulated kinase and phosphatidylinositol 3-kinase (PI3K)/protein kinase B pathways, cyclic adenosine monophosphate response element-binding protein phosphorylation, and increased c-fos expression. 21,42
GFRα1 and GFRα2 expression is more prominent in the CNS whereas GFRα3 and GFRα4 are predominantly expressed in the PNS. 43 Because GDNF is unable to cross the blood–brain barrier because of a lack of transport protein, it cannot be given orally or intravenously. However, conjugating or fusing GDNF with other molecules, such as viral proteins or antibodies, could overcome this limitation. 44 –50
Further, GFRα1 has been shown to form a complex with NCAM, mediated by the N-terminal domain of GFRα1 and the fourth immunoglobulin domain of NCAM, which is important for GDNF high-affinity binding and downstream signaling. 51,52 Mediated by the cyclin-dependent-like kinase 5 (Cdk5) kinase, soluble GFRα1 is capable of capturing GDNF, presenting it to axonal Ret receptors, and thereby inducing axonal growth in a directional fashion. 53 Additionally, it was accidently discovered that the expression of GFRα1 receptor allowed GDNF ligand to induce the formation of synapses in hippocampal neural cultures, initiating ligand-induced cell adhesion (LICA), a property the receptor by itself was unable to accomplish in standard cell aggregation assays. 38
Several targets in the brain have been identified to express GDNF messenger RNA, including striatum, nucleus accumbens, thalamic nuclei, olfactory tubercle, hippocampus, cerebellum, and cingulate cortex as well as in the internal granular cell layer of the olfactory bulb. 28
Initially, heparin sulphate (HS) was suggested to interact with GDNF, 54 which led to research that believe HS glycosaminoglycans are required for GDNF signaling. 33 Other studies suggested that GFR-α-RET complex to be the main mechanism of interaction between GDNF and HS. 55,56 Additional data further implicated the involvement of integrin β1 adhesion molecule with a GDNF-induced NCAM mechanism with the heparin interaction. 57 Moreover, other researches explored the presence of heparin in GDNF functionality, highlighting the role of heparin-binding determinants in keeping GDNF concentrated at the injection site. This benefit was further validated by showing improvement of rotational behavior in a 6-hydroxydopamine (6-OHDA) striatal injury model. 58 Later work also identified Sydecan-3 as an alternate GDNF receptor and its role in cellular adhesion mechanisms. 59
Role of Glial Cell Line-Derived Neurotrophic Factor in the Nervous System
GDNF has developmental, neuroprotective, and neurotrophic effects on several types of neurons in the CNS and PNS. 60 GDNF has a broad impact on recovery post-SCI, including protection, axon regeneration, remyelination, neuronal survival, and synaptic formation. 61 In addition, the successful axonal regeneration can consequently lead to recovery of motor function post-SCI.
Development and neurodifferentiation
In mammals, GDNF concentration decreases with development and is undetectable in adults. 62 Initially, the developing forebrain region exhibits the strongest GDNF expression. Ultimately, GDNF will be expressed along the entire anteroposterior axis. 20 RET is the first GDNF receptor identified. It is a tyrosine kinase receptor that can be mutated in several endocrine conditions, namely multiple endocrine neoplasia and Hirschsprung's disease. 43 Gershon and colleagues showed that the RET activation by GDNF is essential for the pull information, migration, and differentiation of enteric crest–derived cell. More specifically, GDNF is essential for gangliogenesis of the bowel, with the exception of the esophagus and adjacent stomach, thus explaining the colonic aganglionosis observed in Hurshsprung's disease. 63 GDNF was also used as a marker of differentiation of reprogrammed mesenchymal stem cell into dopaminergic-like cells. 64
Chemoattractant and migration
GDNF plays a role in chemoattraction in several developmental and neurological systems, a function best explored in embryological studies. GDNF and its receptor, RET, assist with directing the growth of ureteric bud epithelium and its branching. 65,66 More specifically, RET activation increases cell motility and scattering, whereas GDNF acts as the destination for Madin-Darby canine kidney epithelial cells, which resemble renal distal tubular epithelium. 67,68 On the neuronal front, GDNF's chemoattraction capabilities were demonstrated by showing the migration of rostral migratory stream RMS-derived neuronal precursors. 69 GDNF increased Cdk5 activity in RMS cells and supported the participation of NCAM during this guidance process. 69 Expression of GFRα receptor post-axotomy also highlighted the role of GDNF in modulating the formation of penis projecting autonomic neurons. 70 The development of the enteric nervous system is also dependent on GDNF. Directional migration of enteric neurons is dependent on GDNF and is crucial for normal gut development. 71 More recently, GDNF was described as a potent chemoattractant for SCs, another descendant of the neural crest cells. 72 GDNF also induced attractive turning of spinal lateral motor column (LMCL) axons. 73
Neuroprotection
Neuroprotection is the therapeutic strategy intended to slow or halt the progression of neuronal loss. 74,75 Combinations of neurotrophic factors have been used to induce neuroprotection post-SCI. 24 GDNF has been used as a component of cerebrolysin, a combination of neurotrophic factors, to induce neuroprotection subsequent to intoxication from engineered metal nanoparticles. 76 In this study, in order to induce the same level of neuroprotection, a higher dose of cerebrolysin was needed when given post-injury versus pre-injury dose. 76 Cerebrolysin has also been shown to rescue avulsed spinal motor neurons when administered intrathecally, 77 help reinnervate avulsed peripheral nerves, 78 reduce permeability of the blood–spinal cord barrier (BSCB), 79 and modulate morphine induced pain. 80 Although all these effects cannot be exclusively attributed to GDNF, the neuroprotective effect of cerebrolysin, at least in part, can be attributed to GDNF.
The neuroprotective role of GDNF has been attributed to the reduction in the permeability of BSCB, and the downregulation of nitric oxide synthase (NOS), and heat shock proteins expression subsequent to spinal trauma. 81 Aside from GDNF's effect on motor function, GDNF applied topically 30 min post-SCI attenuated the leak of tracers across the BSCB. 81
The BSCB plays an important role in the pathophysiology of edema and cell injury post-SCI. 82,83 Spinal cord trauma disrupts the BSCB through direct impact on microvessels as well as the induction of release of vasoactive molecules. 83,84 The BSCB can be evaluated using Evans blue, 85 Iodine, and Lanthanum La3+ tracers. 84 Using these tracers in an SCI model, GDNF reduced the extravasation of all tracers by over 50% when given 30 min post-SCI. 81 Formation of cord edema is also an important factor in cord pathology and functional recovery, which may be influenced by GDNF. 81 GDNF reduced cord width, as a surrogate for cord edema, by 20% when given 30 min post-SCI. The reduction in pathological signs of edema evidenced by water content were attenuated with GDNF use when given 30 min before to 10 min after SCI. This attenuation decreased with later administration of GDNF, that is, 30 and 60 min post-SCI. 81
Several studies support the idea that the upregulation of NOS is detrimental to the spinal cord. 86 –92 In addition, counteracting the effect of NOS attenuated cord edema and decreased cell injury. 89,90,92 Given that GDNF reduces NOS upregulation, it is conceivable that this mechanism explains some of the neuroprotective effects of GDNF. 82,86,93 –95 Finally, the main goal of neuroprotection is preservation of neural cells, and it is prudent to remember the importance of endothelial and glial cells for nerve cell function, including astrocytes. 96,97
GDNF is produced by human astrocytes. 98 Cultured astrocytes express both tumor necrosis factor alpha (TNF-α) receptor 1 (TNFR1) and TNF-α receptor 2 (TNFR2). GDNF is expressed when these receptors are activated by TNF-α. 99 Astrocytes also express GFRα1, which could imply that they are sensitive to GDNF through autocrine and/or paracrine mechanisms. 39 However, increasing the GDNF concentration did not increase the number of astrocytes. 31
GDNF has been shown to have survival-promoting effects on the following cell populations: noradrenergic, 42 cortical, 100 –104 retinal ganglion, 105,106 sensory, 107,108 parasympathetic, 21 peripheral, 109,110 motor neurons, 100,111 –121 neonatal motor neurons, 119,122 microglia, 123 dopaminergic, 31,57,58,64,124,125 dopamine-like mesenchymal stem cells, 64 and retinal Müller glial cells. 126,127
Axon sprouting
GDNF supports the survival of axons by acting as a chemoattractant for axons and some migration. Starting with embryogenesis, Charoy and colleagues reported that in the developing spinal cord, prominent source of GDNF is present in the floorplate where commissural axons cross the midline. 128 Interestingly, this study showed that this effect is RET independent and mediated by NCAM/GFR-α1 signaling. 128
Boyd and colleagues illustrated the importance of long-term treatment of continuous exogenous GDNF on increasing the axonal sprouting within the distal nerve stump. 129 In this study, however, the beneficial effect of GDNF was not dose dependent.
After a unilateral corticospinal tract (CST) injury, GDNF, expressed from the unlesioned side, was shown to support the enhancement of axonal sprouting. 130 This was described as a compensatory process as a result of injuring the contralateral side. This phenomenon resembles findings observed in SCI models using other treatments, namely IN-1, 131,132 inosine, 133 neurotrophin-3 (NT-3), 134 and brain-derived neurotrophic factor (BDNF). 135 To better characterize this phenomenon, anti-GDNF antibodies were used to neutralize the peripheral effect of GDNF post-axotomy. Interestingly, a low dose of anti-GDNF antibodies potentiated, rather than inhibited, axonal branching of motor neurons. 136 This effect was interpreted as a result of controlling the overexpression of endogenous GDNF, 136 thus highlighting the importance of post-injury GDNF levels. In another study, peri-wound sprouting fibers grew toward the wound margins along an increasing trophic gradient of GDNF generated by macrophages and microglia. 137 However, the growth ceased at the point of maximal neurotrophic expression at the wound edge. 137 On the other hand, GDNF failed to enhance axonal spouting in retinal ganglia cells, suggesting the cell specificity of this process. 138
Neuroplasticity
GDNF has been described to promote the survival and morphological differentiation of dopaminergic neurons and increase their dopamine uptake. 31,139 In the dopaminergic nigrostriatal neurons, the effects of GDNF are mediated by a multi-subunit receptor system. 140 This leading to the expression of GFRα1 receptor allows GDNF ligand to induce the formation of synapses in hippocampal neural cultures, through LICA. 38 This property was further explored by attempting to use GDNF for treating PD. One study attempted delivering biodegradable microspheres containing GDNF as a treatment for partial dopaminergic lesions. 141 The study showed improved rotational behavior induced by amphetamine in GDNF-treated animals. Increase in the density of tyrosine hydroxylase–positive fibers at the striatal level was also noted. Whether this recovery represents compensation for the injury or merely the survival of the injured neurons is unclear.
GDNF enhanced both the number and caliber of regenerated axons and increased the neurotic outgrowth of dorsal roots ganglion neurons. 142 Applying GDNF on sciatic nerve grafts placed in a gap prompted by complete transections of the spinal cord enhanced motor recovery and fiber growth. 143 This promotion of differentiation of dopaminergic neurons in the brainstem and the translated sciatic nerve grafts suggests that GDNF directs the ability to enhance neuroplasticity. Interestingly, expression of GDNF in macrophages/microglia increases post-SCI. 140 Nonetheless, it is important to realize that the paracrine/autocrine effects of GDNF on specific macrophage/microglial function remain unclear.
Glial Cell Line-Derived Neurotrophic Factor Knockout Studies
GDNF has been found to be a crucial element for the survival of knockout mice, who without GDNF, RET, or GFRα1 died soon after birth. 40,144 These mice all share kidney agenesis and the absence of several enteric and parasympathetic neurons. 40 RET was expressed in virtually all cells and tissues affected in GFRα and GFL knockout mice, suggesting that RET is the main signaling receptor in vivo for GFLs for developmental functions. 21
Further, lacking GDNF affected a myriad of neuronal structures spanning all aspects of the autonomic nervous system. Newborn mice deficient in GDNF, RET, or GFRα1 had no otic and sphenopalatine ganglia, suggesting that GDNF signaling by the RET-GFRα1 receptor complex is required for the proliferation and migration of parasympathetic neuronal precursors and thus essential for parasympathetic neuronal development during embryogenesis. 145,146 Similarly, neuronal hindrance of the sympathetic nervous system was signified by the absence of superior cervical ganglia (SCG) neurons. However, in mice that lack GFRa1, only 30% loss of SCG was reported. 21,40 Several of the petrosal ganglion's primary sensory neurons required target derived GDNF for survival, before birth. 147 In addition, GDNF- and RET-deficient mice showed disturbances in breathing and respiration control attributed to a decreased in the survival of visceral chemoafferent sensory neurons innervating the carotid body. 21
Mice lacking GDNF, RET, or GFRα1 also displayed missing glial cells and enteric neurons that were derived from the sacral and vagal neural crest, below the stomach. Knockout mice with GDNF-heterozygous had a variable enteric phenotype closely resembling Hirschsprung's disease. 148,149
A lack of GDNF or GFRα1 was not a factor for the number of sensory neurons found in spinal and trigeminal ganglia of newborn mice. 150,151 However, post-natal survival required GDNF signaling in GFRα1/RET-expressing subpopulations of sensory neurons. 21 Postnatal loss of myelinated mechanoceptors was noted in GDNF-heterozygous mice, suggesting that GDNF supports cutaneous sensory innervation. 152
Primarily, SCs produce GDNF during development. 21 When compared to wild types, GDNF-deficient mouse embryos displayed a substantial loss of cranial and spinal motor neurons and a corresponding increase in dying cells. 153 Continued GDNF subcutaneous injections or transgenic overexpression in post-natal life or skeletal muscles produced hyperinnervation of neuromuscular junctions. This occurred through the promotion of terminal branching, ultimately resulting in continuous synaptic remodeling at the neuromuscular junction. 154,155 This implies that the primary role of GDNF in post-natal motor neurons is to promote synapse formation as well as terminal axonal branching. 4
Combination with Other Neurotrophic Factors
GDNF was most effective in neuron recovery when combined with other neurotrophic factors. The following combinations were shown to increase axonal sprouting and aid in the axon regeneration process. GDNF/NT-3 locally increased the sprouting of injured CST neurons. 156 GDNF/NGF (nerve growth factor) improved axon elongation and branching 157 and rescued nearly 100% of sensory neurons. 158 GDNF/BDNF reduced spinal cord pathology and motor dysfunction 24 and increased the number of motor axons. 25,159 GDNF-microspheres/vascular endothelial growth factor enhanced neuroregeneration/neuroreparation in the substantia nigra. 160
GDNF/BDNF/NT-3 w/PNS tissue grafting regrew injured CST axons in the gray matter. 161 GDNF/BDNF/TGF/fibroblast growth factor 2/GDF5 increased survival of dopaminergic neurons. 162 GDNF/Neurturin improved the development and survival of enteric neurons and provided structural and functional recovery in PD. 23,32 GDNF/Netrin-1 increased length of transplanted dopaminergic axons. 163 GDNF/growth/differentiation factor 5 further increased survival rates of dopaminergic neurons. 164 Finally, cerebrolysin, a combination of BDNF/GDNF/NGF ciliary neurotrophic factor and other peptides was neuroprotective in treating SCI post-nanoparticle intoxication. 76
Glial Cell Line-Derived Neurotrophic Factor Treatment after Spinal Cord Injury
Timing
Several studies administered some form of GDNF treatment and assessed several aspects of neurological function (Table 1). Most studies initiated the GDNF either before or immediately after the SCI. In several studies, transplantation of a nerve graft in the injury site was performed. Pre-treatment of the lesion site with GDNF has been shown to significantly increase the number of neurons after transplantation. 165 Cheng and colleagues showed that initiation of GDNF therapy within the first hour was crucial for functional restoration. 111 Additionally, findings from Tzeng and colleagues demonstrated that initiation of GDNF therapy 24 h post-injury failed to elicit any functional recovery. 111,123 This was explained by the presence of the secondary injury mechanism. It was initially proposed that GDNF therapy is effective for 2 weeks 122 ; other studies provided evidence of the efficacy of treatment beyond that time frame. 100,112,113
GDNF, glial cell line-derived neurotrophic factor; SD, Sprague-Dawley; DRG, dorsal root ganglia; SC-GDNF, Schwann Cell expressing GDNF; OECs, olfactory ensheathing cells; rhGDNF, recombinant human GDNF; pEGFP, plasma expressing green fluorescent protein; i.v., intravenous; NSCs, neural stem cells; BBB, Basso, Beattie, and Bresnahan; N/A, not applicable.
Methods of administration
The GDNF delivery method to the injured cord varied significantly, ranging from genetically modified GDNF-secreting cells, 61,166,167 osmotic minipumps, 60,107 channels seeded with SCs containing GDNF, 142 to direct injection into the spinal cord. 111 Other forms of GDNF delivery include poly(lactic-co-glycolic acid) microspheres, 85,168,169 injectable alginate hydrogel, 170 and intrathecal epigallocatechin gallate induction of GDNF. 171
Several genetically modified cells have been used for GDNF delivery to the spinal cord. These cell types include: SCs, 61,142,166,172 olfactory ensheathing cells (OECs), 173 neural stem cells (NSCs), 174 and fibroblasts. 114 While comparing cell types for delivering GDNF to injured spinal cord, several observations can be deduced. First, different protocols and primary outcomes were used with each cell type. Second, although the success of gene therapy was confirmed in all cell types, the final concentration of GDNF in vivo was not assessed for all studies. Third, OECs provided the most promising functional recovery. 173 Fourth, fibroblasts provided probably the most histologically favorable results; however, the translation of these histological findings into statistically significant functional improvement was limited. 166 Finally, studies that utilized Schwann Cells (SCs) had histologically significant decrease in lesion size, but only one study demonstrated a significant increase in Basso, Beattie, and Bresnahan (BBB) score. 61
Inflammation
It has been shown that GDNF has the capacity to modulate the microglial inflammatory response. 175,176 GDNF reduced the expression of proinflammatory cytokines interleukin-6 and TNF-α, 176 which are upregulated in cerebral inflammation and neurodegeneration. 177 GDNF and other members of the GDNF family reduce microglial activation by inhibiting p38 mitogen-activated protein kinases (MAPKs)-mediated inflammatory responses, 177,178 by increased phosphorylation of p38MAPK. 178 It should be noted that neuroinflammation leads to decreased levels of GDNF, 179 which, in turn, leads to a loss of anti-inflammatory capacity. 177
Concentration
Different concentrations of GDNF were used in the previous studies ranging from 1 to 300 μg/mL of recombinant human GDNF (rhGDNF). Dose-dependent variation in response was noted in one study, 180 where GDNF was administered using a micropump. However, with intracortical administration, only low concentrations of GDNF (2–40 μg/day for 7 days) supported corticospinal neuron survival. 100 In contrast, higher concentrations of GDNF failed to induce any rescue effect. This was attributed to desensitization of GDNF receptors by the higher concentrations. Additionally, in pre-conditioned neurons, GDNF therapy promoted axonal growth beyond the lesion site by 1.4 ± 0.3 mm at the lower dose (0.2 ng/day for 7 days). 180 Higher doses seemed to further enhance the growth (at 2 and 20 ng/day for 7 days); however, the growth was not hindered at the higher doses. Given the findings of these studies, it is conceivable that GDNF demonstrates saturation kinetics at low concentrations, comparable effect with moderate doses, and overwhelming desensitization, even harmful, at ultrahigh doses.
Directional guidance
Directional guidance in neural regeneration is an important matter. Several studies provided evidence to support the need for a neurotrophic factor gradient. 181,182 For example, GDNF induced attractive turning of LMCL motor axons. 73 Activated microglia and macrophages inherently also express GDNF at progressively increasing amounts as the wound edge is approached. 183 This gradient governed the directional growth of dopaminergic fibers and loss of the gradient led to a decrease in sprouting in the striatum. 137,184 Interestingly, cell death can also be configured in a gradient, as in models where 6-OHDA injections are used to induce CNS lesions. 185 Therefore, an increasing gradient of GDNF is much more likely to support directional growth than ungraduated therapy. This provides a potential integrative understanding of the axonal sprouting and chemoattractive properties of GDNF.
Adverse effects of glial cell line-derived neurotrophic factor
To our knowledge, no significant serious side effects were reported with GDNF therapy administered for SCI. In addition, no benefit was reported when GDNF was administered before 24 h post-injury. On the other hand, significant weight loss was a major side effect reported, particularly after intracerebral neurotrophic factor treatment. With intracerebral treatment, weight loss occurred at all given doses. 186 GDNF given intracisternally, rather than intraventricularly, at low doses was able to promote the survival of corticospinal neurons without negatively affecting rat body weight. 186
Mechanism of action
Recovery after SCI is hindered by the inability of severed axons to regenerate. Several studies provided evidence to help explain this inability. Proposed explanations include: lack of neurotrophic stimulation, 187 presence of myelin-associated inhibitory molecules, 180,188 and extracellular matrix inhibitors, 189 extensive post-injury secondary inflammation, 190 diminished intrinsic capacity of adult neurons for growth, 191,192 and absence of permissive substrates promoting growth. 193 GDNF counteracts some of these injurious processes by providing neurotrophic support, reducing lesion volume, and increasing spared white matter. This is achieved by reducing reactive astrocytosis and macrophage accumulation, thus negating some of the effects of myelin and glial scar inhibition and chemorepellent molecules, as discussed below.
Therapeutic Effect of Glial Cell Line-Derived Neurotrophic Factor
Lesion size
The lesion size at the injury site can be measured to analyze the extent of damage post-SCI. Figure 2A shows three studies that performed a variety of GDNF treatment and then measured lesion size. Exogenous administration of GDNF by the tail vein was the most successful in decreasing lesion size. GDNF administered systemically will migrate to the injury site and participate in the healing process by ameliorating apoptosis in the injured spinal cord. 194 The next two studies used different delivery mechanisms, but showed approximately the same amount of improvement in lesion size. 60,140 The GDNF infusion (Alzet miniosmotic pump) allowed for constant intrathecal infusion of GDNF to penetrate into the cord parenchyma that surrounded the injury. This resulted in a reduction of the total lesion volume and an increase of spared white matter, 60 attributed to the neuroprotective effects of GDNF. GDNF in Matrigel promoted propriospinal axonal regeneration by GDNF at the lesion site undergoing retrograde transport along the severed axons. GDNF treatment produced a significant reduction in lesion size by reducing reactive astrocytosis, macrophage accumulation, and cavitation at graft-host interfaces. 140
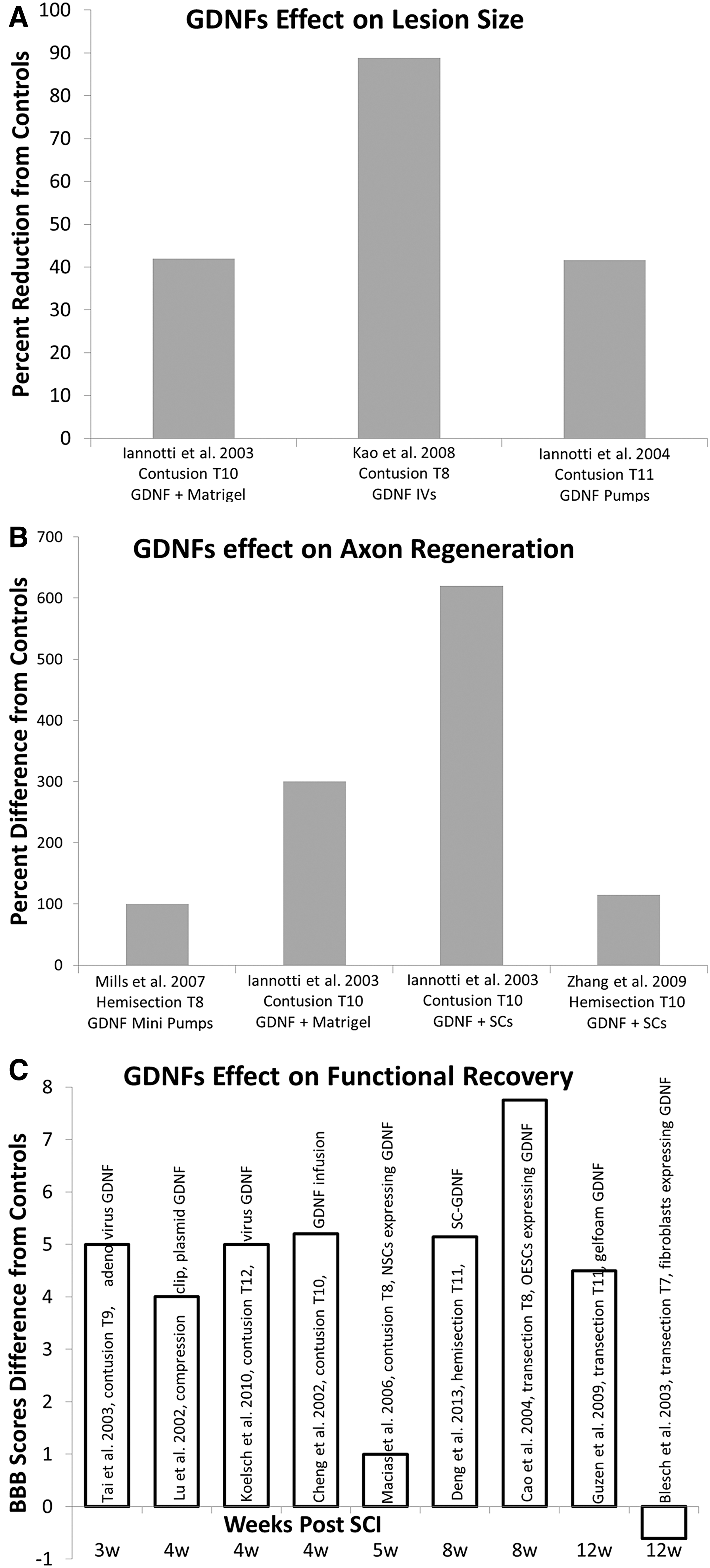
Survey of GDNF's effect on functional improvement, lesion size, and neuroprotection post-SCI. All data are depicted from the authors listed in the series legend, which includes injury model and animal used. Data are presented as percent comparison to each study's vehicle control. (
Axon growth and regeneration
Axon growth post-SCI is difficult because of inhibition by myelin, glial scar tissue, and chemorepellent molecules. 195 GDNF shows promise in aiding to overcome this inhospitable environment. Four studies that performed a variety of GDNF treatments and performed BBB scores at varying time points also measured axon regeneration (Fig. 2B). SCs expressing GDNF showed the greatest improvement in axon regeneration. GDNF treatment had a strong neuroprotective effect on white matter sparing and sparing of a subset of proprio- and supraspinal axons. 60 Propriospinal tracts showed the strongest regenerative response. 196 GDNF in Matrigel enhanced neurite outgrowth from spinal cord explants by increasing the total surface area and density of axonal processes. 140 This method ranked second in improving axon regeneration. The GDNF group also showed a significant increase in blood vessels and the regeneration of a large number of unmyelinated axons, with a lower ratio unmyelinated to myelinated axons. Mills and colleagues used minipumps to deliver GDNF. Low concentration of GDNF applied directly to the dorsal root ganglia (DRG) neurons by minipumps showed that GDNF significantly increased both the percentage of neurons that initiated neurites and neurite outgrowth. 140 SCs expressing GDNF showed that treatment with GDNF enhanced both the number of caliber of regenerated axons in vivo and increased neurite outgrowth of DRG neurons in vitro. This was attributed to the direct effect of GDNF on neurons. The combination of GDNF and SCs allowed for enhanced axonal regeneration and spinal cord myelination.
Functional recovery
The primary objective of most SCI treatments is to increase the level of motor function that is either retained or regained post-injury. The BBB scoring method for rats with an SCI was the most predominantly used analysis to assess functional recovery. The amount of BBB score improvement varied between studies was attributed to different SCIs (contusion, hemisection, and complete transection), level of injury, GDNF concentrations, and delivery mechanisms. GDNF delivered by OECs were the most successful in improving functional recovery, with an increase in BBB score of almost 8 compared to their controls (Fig. 2C). The GDNF gene was transduced into OECs using a retroviral system. These genetically modified OECs were able to produce high levels of GDNF in vivo, which significantly improved recovery post-SCI by stimulating the regeneration of cortico- or rubrospinal axons. 173 The combination of transplantation and gene therapy may be the most effective strategy to promote CNS repair. 173 The majority of the investigators observed an increase of approximately 5 in BBB score compared to the studies control group. Deng and colleagues formed a continuous growth-promoting pathway by grafted SCs overexpressing GDNF. This combination bridged the lesion gap and also extended into the caudal host spinal cord, which allowed for enhanced axonal growth. Both grafted and host-derived SCs were responsible for the myelination. 61 A major benefit of this delivery system is that the graft-host interference can be modified to be more permissive to axonal growth. Sciatic nerve grafts treated with GDNF also showed a significant increase in functional recovery at both 4 and 8 weeks. Nerve grafts provide an adequate environment by supplying morphological and chemical substrates for fiber growth. 197 Certain cells of the nerve grafts, such as fibroblasts and SCs, are able to express various axonal-growth–promoting factors. 198 Exogenous GDNF might have amplified the properties of the nerve graft to promote spinal cord regeneration and repair. In another study, simvastatin was used as an indirect way to upregulate the expression of GDNF and BDNF, 199 because of its lipophilic properties, which make it capable of crossing the BSCB. 200 Simvastatin therapy significantly reduced myelin loss and increased BBB scores.
All investigators observed a significant improvement in motor function, except Blesch and colleagues 2003 and Macias and colleagues 2006 (Fig. 2C). Blech's group performed a complete transection and removed 3 mm of spinal cord at the T7 level on male Fischer rats, and then implanted GDNF secreting fibroblasts embedded in collagen gels. When BBB scores were assessed at 6 weeks and 3 months, there was no significant difference when compared to controls. Although they did not observe a significant improvement in BBB scores, they did observe histological differences and significant sensory improvement. Blesch and colleagues is also the only study that used Fischer rats. Considering the strain differences in rodents with regard to the capacity of endogenous regeneration, 201 –204 it is possible that Fischer rats host a different capacity for regeneration post-SCI. Macias and colleagues used immortalized NSCs to deliver GDNF into the injured CNS by ex vivo gene transfer. There was no improvement in locomotor function. However, after transplantation with NSC populations, regardless of whether GDNF was included, there was significant forelimb thermal and mechanical allodynia. They observed that, post-injury, NSCs proliferated and differentiated almost exclusively into astrocytes, which could explain the increase in allodynia and lack of functional recovery. 205
Conclusion
GDNF is a potential treatment for reducing secondary damage, promoting axon growth, and restoring some of the lost function post-SCI.
By affecting both neurons and support cells, GDNF have been shown to decrease lesion size, improve allodynia, and regenerate axons in the CNS and PNS. Exogenous administration of GDNF either before or immediately after SCI is most effective. GDNF delivery through genetically modified cells has been used with varying success. GDNF seems to work more effectively in combination with other neurotrophic factors. In conclusion, animal studies have shown that administration of exogenous GDNF post-SCI is a desirable treatment post-SCI because of its ability to reduce lesion size, and increase functional recovery and axonal regeneration.
Footnotes
Author Disclosure Statement
No competing financial interests exist.