
Abstract
Transplanted human olfactory ensheathing cells (hOECs) were mixed with collagen into a unilateral transection of four dorsal roots (C6–T1) in a rat model. By mixing with collagen, the limited numbers of hOEC were maximized from an olfactory bulb biopsy and optimize cavity filling. Cyclosporine was administered daily to prevent immune rejection. Forelimb proprioception was assessed weekly in a vertical climb task. Half of the rats receiving hOEC transplants showed some functional improvement (“responders”) over six weeks of the study while the other half did not (“nonresponders”) and performed similarly to “injured only” rats.
Transplanted cells were seen at both one week and six weeks after the surgical procedure; many were concentrated within the lesion cavity, but others were found with elongated processes in the overlying connective tissue. There were some fibers in the injury area associated with transplanted cells that were immunostained for neurofilament and TUJ1. Responder and nonresponder rats were compared with regard to microglial activation within the deep dorsal horn of cervical levels C7, C8 and also axon loss within the cuneate fasciculus at cervical level C3. Little difference was seen in microglial activation or axonal loss that could account for the improved proprioception in the responders group. This preliminary study is the first to transplant human olfactory bulb cells into a rat model of dorsal root injury; by refining each component part of the procedure, the repair potential of OECs can be maximized in a clinical setting.
Introduction
I
Olfactory ensheathing cells (OECs) are specialized glial cells that can be obtained from either olfactory bulb or olfactory mucosa tissue. 7 Transplantation of OECs has been shown to be effective in a number of different experimental central nervous system injury models. 8 –10 It has been shown that OECs have the potential to promote axonal regeneration, remyelination, angiogenesis, glial scar remodeling, and immune system interactions. 11 –15 Functional restoration of long-tract injuries to reinstate forepaw grasping, supraspinal control of breathing, or proprioceptive function on a climbing task has also been demonstrated in various rat studies. 16 –21 A trial in which dogs with a thoracolumbar spinal cord injury showed improved limb coordination after olfactory mucosal OEC transplantation also helped lay the foundations for their use in a clinical setting. 22
Clinical trials of transplantation of autologous OECs into spinal cord injuries using the more accessible cells obtained from the olfactory mucosa 23 –25 showed little or no neurological improvement as shown in animal studies. 26 –28 Transplantation of autologous cells obtained from the olfactory bulb in a patient with complete thoracic spinal cord injury caused by a transection, however, showed that the patient had better trunk stability, increased muscle mass in the left thigh and calf, and some return of voluntary movements and sensation in the lower extremities after two years. 29 The precise mechanism underlying these improvements is hard to pinpoint exactly, because the procedure also involved spinal cord resection, a dose of methylprednisolone, and an extensive rehabilitation program. The outcome of this clinical application using the cells from the olfactory bulb is consistent with the findings from the animal studies. During the course of the surgical procedure, Tabakow and colleagues 29 used autologous peripheral nerve strips to span a gap of approximately 11 mm between the severed spinal cord stumps after injecting cells at multiple sites into the spinal cord rostral and caudal to the lesion.
In the present study, we developed a method to mix the cells with a collagen gel to maximize the use of the limited number of human OECs (hOECs) from the olfactory bulb biopsy. This cell-collagen scaffold would occupy a cavity and give structural integrity to support cell transplants spanning the lesion. Placing the transplant in a scaffold form would avoid time-consuming multiple injections that may cause further damage to the spinal cord. In addition, this scaffold would enable the transplanted cells to have direct contact with the nerve graft and the spinal cord. In addition, if OECs had a pre-surgical period at which to differentiate, produce neurotrophic factors, and elongate their processes, they could more easily integrate within the injured host environment and optimize any potential for repair. 30,31
We first determined whether the cells would survive and differentiate into their typical morphology within collagen. We then tested their function in vivo by transplanting the cells mixed with gel into a rat model of brachial plexus injury.
We believe that this current study is the first to examine transplantation of human olfactory bulb OECs in a rat model of brachial plexus injury. We show that the cells: (1) survive at the lesion site over six weeks, (2) migrate within and out of the gel, (3) interact with host tissue, (4) induce axonal regeneration, and (5) partially reinstate proprioception in a four root dorsal root transection injury in rats.
Methods
Human olfactory bulb tissue
Obtaining tissue
Human olfactory bulbs (OBs) were obtained in the period between October 2013 and March 2017 from 15 patients who had undergone an operation for repair of anterior skull base fracture with cerebrospinal fluid leak or an operation to remove an anterior skull case tumor. The decision to obtain a patient's olfactory bulb was determined by the clinical and intraoperative evidence of an irreversible lesion of the olfactory bulb or olfactory tract, caused by the skull base fracture or the neoplastic infiltrative process (Fig. 1). The transcranial bifrontal microscopic endoscopy-assisted approach or the endoscopic transnasal approach was used for repair of the skull base fractures. Brain tumors were approached via craniotomy. The removal of OB tissue was approved by the Wroclaw Bioethics Committee and written consent was obtained from all of the patients.
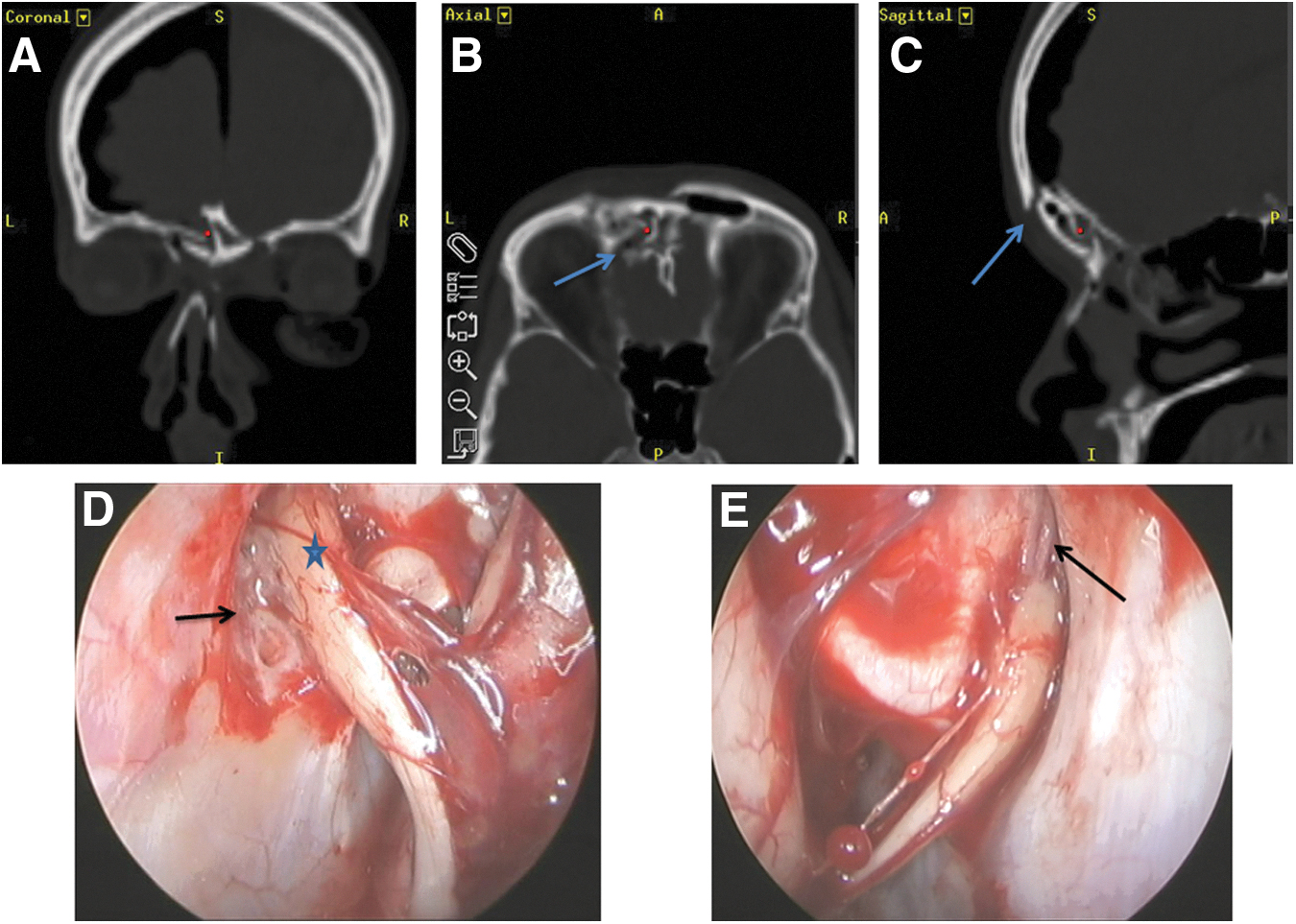
A case of a 35-year-old woman who sustained severe anterior skull base fracture with cerebrospinal fluid leak and pneumocephalus. (
The OBs were transported on ice in a medium containing Dulbecco Modified Eagle Medium F/12 Nutrient Mix (DMEM/F12, Thermo Fischer) and 10% fetal bovine serum (FBS, Thermo Fischer) supplemented with 100 U/mL of penicillin and 100 μg/mL of streptomycin (PenStrep, Thermo Fischer: DMEM/F). Transportation of the tissue took approximately 24 h.
Cell culture
On arrival, images of the OBs were taken and their weights recorded. All dissecting and culture protocols were performed under sterile conditions. After peeling off the adherent meningeal membranes and removing a small piece for histological examination, the OBs were cut open, the outer nerve and glomerular layers of approximately 1 mm thickness were dissected out and cut into 2 mm2 fragments that were then incubated in 1% trypsin in Hank balanced salt solution (HBSS) at 37°C for 15 min. After the enzymatic reaction was stopped by addition of DMEM/F and then spun down at 250g by centrifugation, the supernatant was discarded and the precipitated cell pellet was re-suspended into DMEM/F. Cells were stained with trypan blue, counted on a cell counter (Countess, Invitrogen), and seeded on poly-D-lysine (PDL, Sigma-Aldrich) coated 35 mm plastic dishes at a density of 1.5–2 × 104 viable cells/cm2. The cultures were maintained in a humidified incubator enriched with 5% CO2 at 37°C. Culture medium was first replaced on the fifth day and then changed every three days.
Preparation of collagen gel containing hOECs
After 10 days in culture, the cells were transduced with LV-GFP as described previously. 32 They were then washed three times with DMED/F12 and incubated in trypsin/ethylenediaminetetraacetic acid (EDTA) (TE) for 5 min to lift the cells from the surface of the culture dish. After TE activity was inactivated by adding DMEM/F, the cells were collected and triturated into suspension. The hOEC/collagen gel was made by mixing the cell suspension with collagen, NaOH (Sigma) and modified Eagle medium (MEM, 10 × ; Sigma).
We refer to the cultured human olfactory bulb cells within the gel as hOECs throughout this text. This includes fibroblasts and other cell types such as mesenchymal cells that are found in the olfactory biopsies, but here we simplify the nomenclature to hOEC.
Microsurgery
Dorsal root injury model
Twenty-four male Sprague Dawley rats (200–220 g at operation) received a unilateral four root dorsal root transection injury. Under isoflurane anesthesia, dorsal roots C6, C7, C8, and T1 were exposed as described previously. 33 Briefly, in a posterior approach, a skin incision was made through the dorsal midline, the paraspinal muscles were separated, and the prominent T2 process located. Hemilaminectomies were performed from C4 to T2, and the dura was incised with microscissors to reveal the dorsal roots. The four dorsal roots of C6, C7, C8, and T1 were then transected with microscissors as close as possible to the spinal cord in a plane approximately perpendicular to their entry into the cord.
After hemostasis was achieved, 19 rats received human olfactory bulb cultured cells (hOECs) within a collagen scaffold. In all cases, the transplant was positioned between the central and peripheral stumps of the cut dorsal roots and held in place with fibrin glue (Tisseel Kit, Baxter, Thetford, UK). Immediately after transplantation, cyclosporine (Novartis, Surrey, UK) made up at 15 mg/mL in 20% Tween 80 solution (Sigma-Aldrich) at a dosage of 6 mg/kg with 100% ethanol (VWR, Fontenay Sous Bois, France) was delivered intraperitoneally (IP) via Microfine insulin syringes (BD Ltd, Dun Laoghaire, Ireland) and repeated daily.
The remaining five rats received the same surgical procedure, but no cells were transplanted; cut roots were reapposed and held in place with fibrin glue. With all animals, muscles were repaired in layers, the skin sutured, and the rats were given post-operative pain relief (0.05 mg/kg buprenorphine subcutaneously daily for three days).
In terms of housing conditions, room temperature was maintained between 20°C and 22°C under standard lighting conditions (12:12 light-dark cycle) with food and water available ad libitum. Animals were handled according to the UK Home Office regulations for the care and use of laboratory animals, the UK Animals (Scientific Procedures) Act 1986 with the ethical approval of the University College London (UCL) Institute of Neurology.
Vertical climbing test
On arrival, the rats weighed 150–175 g and were acclimated to housing conditions for one week. The following week, they were handled each day and placed on a 1 m near vertical grid (15 degree inclination) to climb up the rungs to the top. They did this naturally without any need for a reward. Starting one week before operation and for up to six weeks post-operation, rats were video-recorded climbing the vertical cage weekly.
In the subsequent slow motion analysis, the forepaw on the injured side was scored for degree of accuracy in locating and grasping the grid bars. As described previously, 33 a grasp was recorded as successful when there was purposeful movement ending in functional full flexion of the digits to grasp the bars. Unsuccessful grasps were graded from 1–4 in increasing order of severity depending on how far the limb protruded through the grid (1, grid level but no grasping; 2, paw protrudes through grid as far as the wrist; 3, as far as the elbow; 4, as far as the axilla.)
After behavior analysis, rats that received hOEC transplants were separated into nonresponder and responder groups based on vertical climbing performance at two weeks post-injury—if their % error score was above or below 85%, respectively. This was based on previous studies with rat bulb OEC transplants, in which animals that failed to improve by two weeks post-surgery did not show any signs of recovery in the rest of the study (Collins, A and Li, Y, unpublished observations). One rat (what received an hOEC transplant) had to be culled at three weeks post-surgery because there was bleeding under the nails of the injured forepaw because of autotomy; spinal cord sections from this animal were not used for immunostaining.
Tissue processing and immunohistochemistry
The rats were terminated under deep carbon dioxide anesthesia and perfused transcardially with 50 mL 0.1 M phosphate buffer solution followed by 400 mL phosphate buffered with 4% paraformaldehyde for 30 min. The vertebral columns were dissected out from the craniocervical junction to the upper thoracic level and left to harden in the same fixative for 48 h. The spinal cord and associated roots were dissected carefully out of the bony skeleton under a dissecting microscope to preserve the continuity across the avulsed dorsal roots and transplants to the spinal cord. The dissected tissues were placed into 10 and 20% sucrose solution until sunk, frozen with cryospray (Cell Path, UK), and 16 μm sections were cut on a cryostat (Leica CM3050). Tissue from 23 rats was sectioned at cervical levels C6–T1 (n = 15 in the transverse plane; n = 8 in the horizontal plane) to check for the presence of transplanted cells and for immunostaining. Of these, six rat spinal cords were also sectioned at cervical level C3 for quantification of axon loss.
For immunostaining, the sections were incubated in 2% milk (Oxoid Limited, Basingstoke, Hampshire, UK) containing primary antibodies outlined in Table 1 and incubated overnight at 4°C. Secondary antibodies are also detailed in Table 1 (all Molecular Probes, Invitrogen) and were incubated for 2 h in the dark at room temperature. Some sections were counterstained with the nucleus stain, Sytox (1/1000; Thermofisher, UK). Control sections were performed with primary antibodies and the omission of secondary antibodies.
GFAP, glial fibrillary acidic protein; ms, mouse; gt, goat; rb, rabbit; GFP, green fluorescent protein; dk, donkey.
Quantification of neurofilament (NF)+ axons within the cuneate fasciculus at cervical level C3
To assess the degree of axon loss within the ascending dorsal columns, we quantified neurofilament-positive axon bundles in a region of interest (ROI1) within the cuneate fasciculus of transverse sections at cervical level C3. Specifically, we wanted to compare axon bundles at ROI1 between responder rats that showed some return of proprioceptive function with nonresponders that did not show any improvement.
Three responder rats and three nonresponder rats that had six week survival times were chosen at random from their respective groups. For each animal, three cryostat sections at cervical level C3 were immunostained as described below for NF (light and heavy). For ROI1, a square area (600 μm × 600 μm) was imaged immediately dorsal to the medial part of the substantia gelatinosa. For each section, images were taken at ROI1 on both the injured and the noninjured sides of the spinal cord section.
To quantify the number of axon bundles, imageJ (National Institutes of Health, Bethesda, MD) was first used to threshold manually the ROI1 images from injured and noninjured sides of a tissue section. Using the Particle Analysis function, two distinct size ranges of axon bundles were counted: small axon bundles had a diameter of 2–4 μm, large bundles were 4–10 μm. This analysis was performed by someone blinded to whether the tissue came from responder or nonresponder groups.
Quantification of Iba1+ within the deep dorsal horn at cervical level C7/C8
To compare the extent of dorsal horn microglia activation in responder and nonresponder rats, we first selected three animals at random from these groups, all with a survival time of six weeks. For ROI2, a square area (1200 μm × 1200 μm) was imaged adjacent to the central canal within Rexed laminae V and VI. This was performed for both the injured and noninjured sides of the spinal cord, for three separate tissue sections in each animal, all at cervical level C7–C8.
To count the number of cells positively stained for Iba1, imageJ was used to threshold manually the images before the number of Iba1+ cells in ROI2 was found using the Particle Analysis function. Images from injured and noninjured sides of the tissue were analyzed under the same conditions, by a blinded assessor. We distinguished small (400–2000 μm2) microglia from larger, activated microglia (>2000 μm2), because recent evidence has suggested that changes in microglial size might underlie behavioral changes such as after exposure to chronic stress. 34
Statistical analysis
Results are expressed as means ± standard error of the mean, with statistical comparison between groups made using a one-way analysis of variance, to determine F-ratio significance. Post hoc analysis was with Bonferroni multiple comparisons, and IBM SPSS Statistics 22.0 software was used. Details of animal numbers are given below each graph.
Results
Climbing performance
Before operation, rats climbed the vertical cage with baseline error scores of only approximately 5% (see Fig. 2, BL). One week after C6–T1 dorsal roots transection, however, both the control rats (injury alone) and those receiving hOEC transplants had an error score of over 95%—virtually every attempt to grasp a vertical bar with the injured paw resulted in failure.
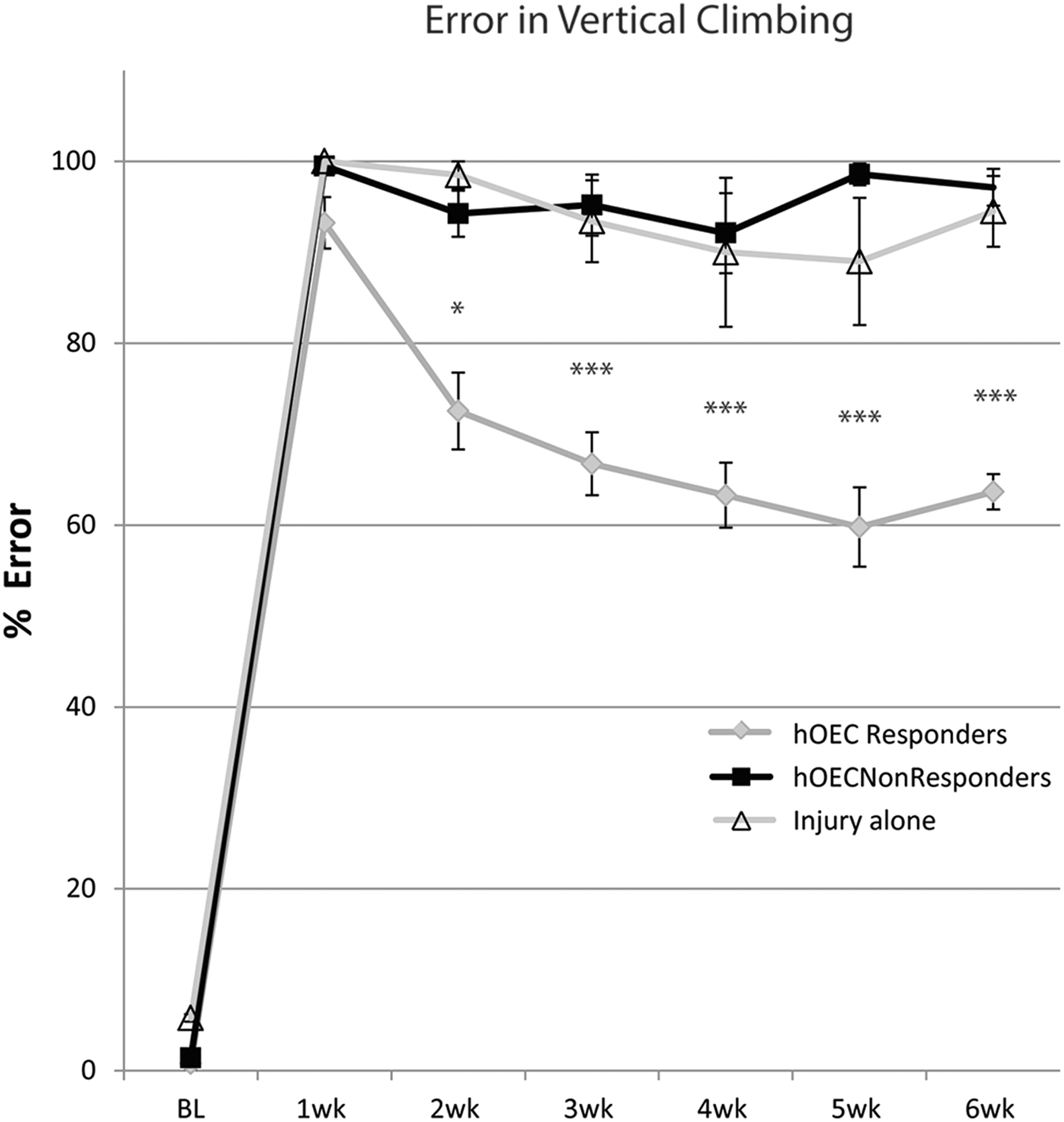
Error on a vertical climbing task. Half of the rats that received human olfactory ensheathing cells (hOEC) transplants showed some recovery (gray circles) after operation; the other half did not (black squares) and performed no differently from rats that had the injury alone (open triangles). Error bars: mean ± standard error of the mean. One-way analysis of variance F(20,126) = 88.763, p < 0.05 (*p < 0.05, **p < 0.01, ***p < 0.001 post hoc Bonferroni). hOEC responders: n = 9; hOEC nonresponders: n = 9; injury alone: n = 5.
The hOEC nonresponder group were rats that received transplants but had no noticeable recovery. Video recording analysis showed no sign of improvement in proprioception over the six-week course of the study; their mean error score deteriorated and was close to 97% in the final week.
In the hOEC responder group, the rats received transplants and also had some functional recovery. This group of rats decreased their error score by a noticeable 20% between one week post-operation (93.2% ± 2.8) and two weeks post-operation (72.5% ± 4.2). Their performance improved gradually each week until at six weeks post-operation when they grasped accurately close to 36% of cage bars (corresponding to 63.7% ± 2.0 error) on the operation side in each upward movement.
In comparison, control rats (injury alone) performed similarly to the nonresponder group. They had a high error score of 100% one week after surgery and then continued to perform poorly (89–99% range) over the course of the six-week study. At six weeks post-surgical procedure, the control rats made approximately 30% more errors than rats from the hOEC responder group (94.5% vs. 63.7%).
Foot fault score
To examine forepaw performance more closely and determine whether nonresponder rats had any subtle improvements in proprioception, we assessed the magnitude of error for each of the misplaced grasps. Misplaced grasps were graded from 0 to 4, with 4 representing the worst level of fault in which the forelimb slid straight through to the axilla.
Before injury, rats had a foot fault score very close to 0 (0–0.4), where almost all grasps successfully located the vertical bars of the cage during upward movements. At one week after operation, injury alone control rats and those in the hOEC nonresponder group both had similar scores of 3.3 (± 0.2) (Fig. 3). This means that the average misplacement of the forearm was to where it slipped through to just past the elbow.
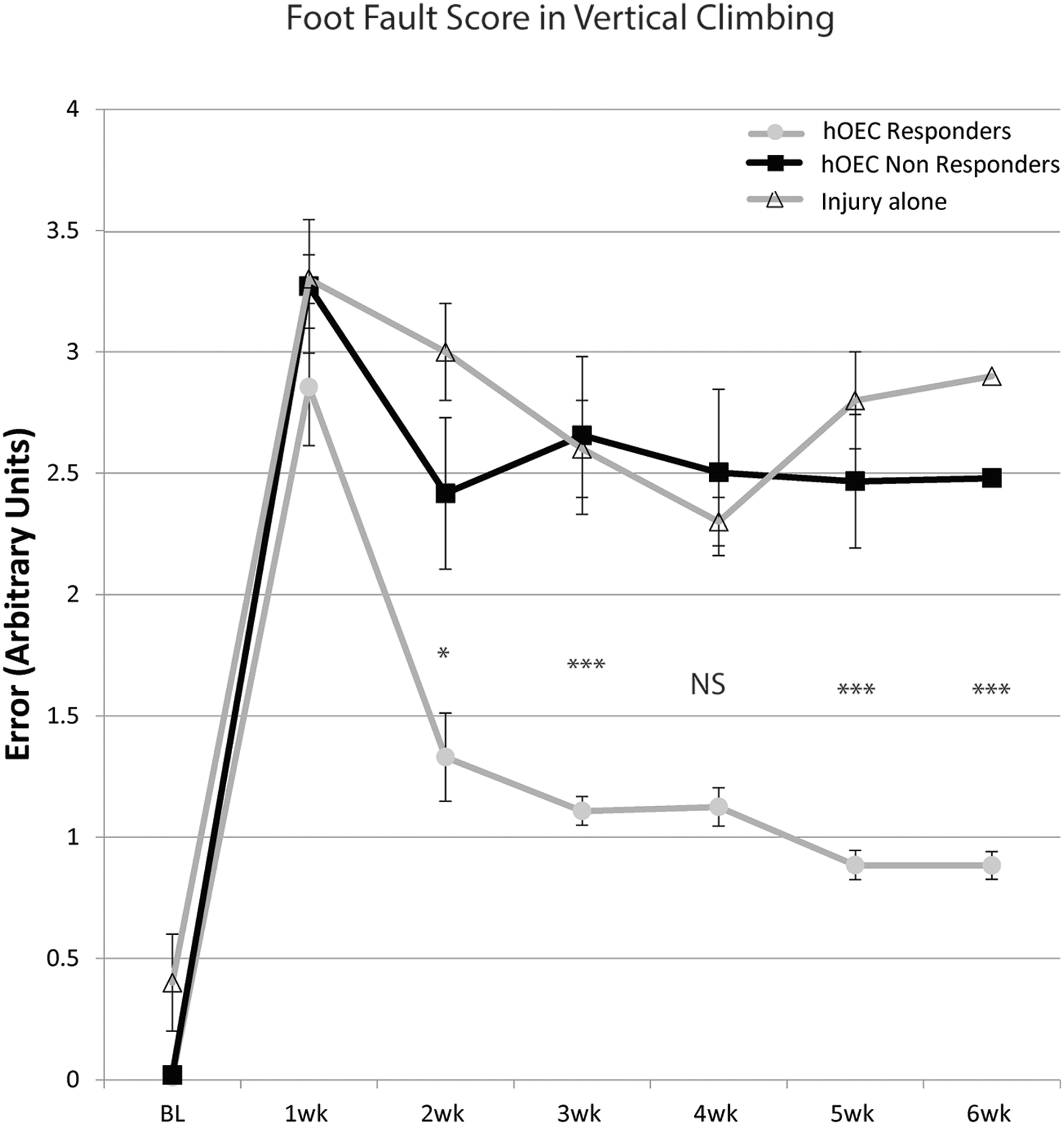
Foot fault score on vertical climbing task—magnitude of error in misplaced grasps. Half of the rats that received human olfactory ensheathing cells (hOEC) transplants had a reduction in the magnitude of grasp misplacement error after the surgical procedure (gray circles); the other half (black squares) showed a similar grasping error misplacement as injury alone controls (open triangles). Error bars: mean ± standard error of the mean. One-way analysis of variance F(20,124) = 23.475, p < 0.001. (NS p > 0.05, *p < 0.05, ***p < 0.001 post hoc Bonferroni). hOEC responders: n = 9; hOEC nonresponders: n = 9; injury only: n = 5.
At the same time point, the hOEC responder group had a slightly lower fault score of 2.9 (± 0.3). These rats then showed a noticeable reduction in foot fault score between one and two weeks post-operation, down from 2.9 to 1.3, indicating that the forearm slipped less far through, to an area between the digits and the wrist. In contrast, at two weeks post-injury, control and hOEC nonresponder rats had foot fault scores of 3.0 (± 0.2) and 2.5 (± 0.3), respectively.
Between four and six weeks post-operation, control and hOEC nonresponder rats showed similar foot fault scores of approximately 2.4–2.8. Rats from the hOEC responder group gradually decreased their foot fault score for the remainder of the study, with a six-week post-operation score of 0.9 (± 0.1), indicating a misplacement slippage only to the palmar area of the forepaw.
Immunohistochemistry
Analysis of OEC-collagen gel
Eight intact olfactory bulbs were used from 15 biopsy samples. After removal of meningeal membrane, a portion of tissue for histology and olfactory tract tissue, the average tissue mass used for OEC culture weighed 84.18 ± 3.91 mg (mean ± standard error). On the third day of culture, a small number of cells started to attach to the bottom of the culture dish and extend processes. By the fifth day, a large number of cells had attached to the dish and differentiated into two major morphology types: (1) small, 5 μm cell bodies with a low ratio of cytoplasm/nucleus in a spindle shape with long, wire-like processes, or (2) large cell bodies (10–20 μm) with a high ratio of cytoplasm/nucleus with a rounded or square shape and short, thick processes. Immunostaining showed that the majority of cells with a spindle shape with long processes were positively stained for p75 (OECs) and the larger cells with short processes that stained for fibronectin were olfactory nerve fibroblasts (ONFs).
On day 10, the culture was 80% confluent and ready for transferring to gel. At this stage, immunostaining showed that the ratio of OECs to ONFs was 1:2. After four days in collagen, the cells had extended their processes to form a continuous meshwork (Fig. 4C,D). Almost 90% of the cells were stained positively for p75 and very few for fibronectin. We found that 4.8 mg/mL collagen with a cell density of 1 × 106/mL was appropriate to give a gel that had enough cells to form a continuous cellular meshwork (Fig. 4D) and at the same time was easy to handle and manipulate for transplantation. Each collagen gel was made up of 250 μL collagen and cells and was approximately 10 mm in diameter and had a thickness of 200 μm.

Cultured human olfactory bulb cells labeled with lentiviral green fluorescent protein (GFP) shown before and after transplantation with various immunostaining. Cultured human olfactory bulb cells immunostained for anti-p75 and anti-fibronectin (FN) along with a DAPI nuclear stain (
Cell survival and distribution
In vitro
Before transplantation, epifluorescent microscopy revealed densely packed hOECs labeled with GFP within the collagen gel (Fig. 4C,D). The hOECs were distributed evenly within the collagen gel to form a cellular meshwork. Cell processes were of varying lengths, from between 30 μm to 130 μm, and extended in all directions within the gel.
The gel itself was pliable and so could be cut into thin strips that facilitated its transplantation into the injury site (Fig. 4B). These strips were easy to manipulate and could be broken down further and tailored to suit the actual size and contour of the junction between the cut ends of the dorsal root and the spinal cord. For example, the first thoracic dorsal root (T1) tended to be narrower and enter the spinal cord at a more oblique angle than dorsal roots C7 and C8. 33 More pieces were thus required for the cervical roots, and ad hoc manipulations of the gel were simple to perform at the time of transplantation.
In vivo
Transplanted hOECs were identified at both early and late stages after operation by using three separate approaches: a lentiviral mediated GFP tag, anti-human mitochondria immunostaining, and an antibody against GFP to amplify the fluorescent signal. Transplanted hOECs were seen close to the dorsal root entry zone at one week and also at six weeks post-surgical procedure. Fluorescent cells tended to be seen either individually or as part of a larger, concentrated group at the injured site (Fig. 4E). The morphology of transplanted hOECs remained similar to those seen before transplantation. Where hOECs were found in a large cluster, it was apparent that they remained within the collagen gel. Individual hOECs that were found in the surrounding connective tissues, however, had elongated processes, which suggests they migrated from the gel.
To account for potential fading of the GFP signal within the gel over the course of the study, immunostaining against human mitochondria and against GFP was employed, as shown in Fig. 4, parts E–I. The scattered distribution of human-mitochondria+ cells within the dorsal horn contrasted with the concentrated aggregation of cells that was often seen (arrows to these cells in Fig. 4E and Fig. 4G) in the center of the cavity and also adhering to its contours (arrowheads in Fig. 4H). The cavity was enclosed dorsally by connective tissue, which presumably helped to retain the transplanted cells at the injured site.
Astrocyte responses and axonal outgrowth
The horizontal sections show that tissue from six weeks post-injury rats revealed outgrowth from the spinal cord in the form of GFAP+ immunostaining. In horizontal sections, multiple strands of filamentous GFAP+ axons were seen to extend for up to approximately 200 μm (Fig. 5A) at the same angle toward the transplanted cells as we have seen in previous studies. Two adjacent sets of serial sections with immunostaining against neurofilament and TUJ1 antibodies showed numerous axonal outgrowths (NF+ in Fig. 5B) at the transplant site in the same region where GFAP+ processes were extended and where TUJ1+ axons were found within the vicinity of the injured area in Fig. 5C.
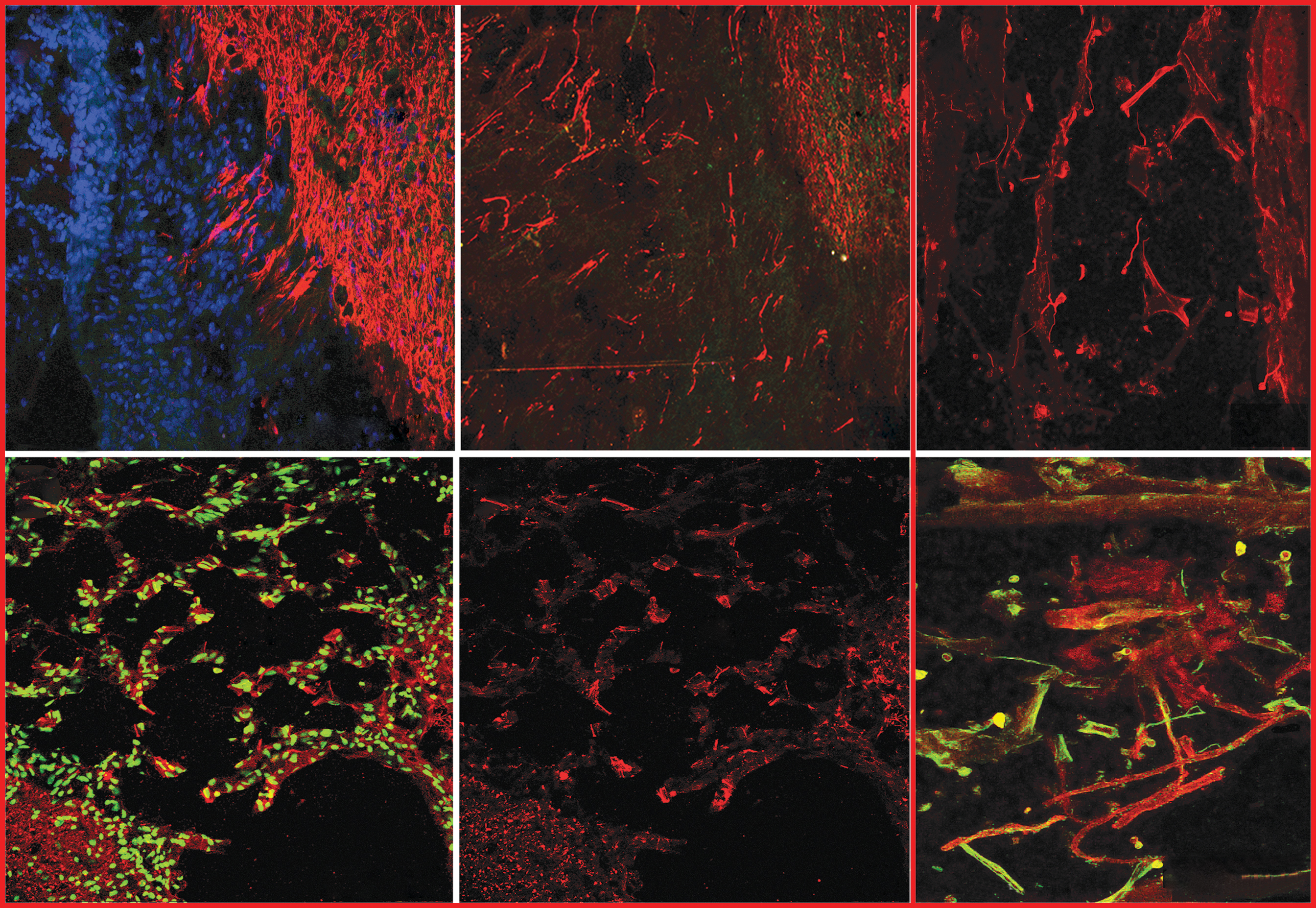
Host spinal cord tissue and axons responded to the human olfactory ensheathing cell (hOEC) transplants after dorsal root transection. The horizontal plane of sections (
Transverse sections also showed neurofilament+ axons (red in Fig. 5D,E) in the damaged dorsal horn, distributed among the vacuole-like spaces that were formed after injury. This damage observed to the dorsal horn (dh in Fig. 5D,E) is typical of a dorsal root transection injury and was most apparent between the injured segments, from the sixth cervical level to thoracic level T1. A number of TUJ1-positive fibers (red in Fig. 5F) were found to associate closely with GFP-positive hOECs (green in Fig. 5F) at the transplant site, with a number apparently showing ensheathment of the newly grown axons by the transplanted hOECs.
Quantification of axon numbers and activated microglia: Responders vs. nonresponders
We examined sections of the spinal cord with adjacent tissue from both responder and nonresponder rats at six weeks post-operation to find any differences that could account for their disparity in proprioceptive performance. We did not see any difference in cell survival between responder and nonresponder rats; tissue sections showed multiple portions of collagen gel at the injury site, where some hOECs remained within the gel and others had migrated into the spinal cord. We quantified axon numbers within the cuneate fasciculus at cervical level C3 and also the activation of microglia within the deep dorsal horn at cervical level C7, C8.
Counting axon bundles from NF immunostaining
The cuneate fasciculus tract has axons concerned with fine touch, fine pressure, and, importantly, proprioception, which synapse at the cuneate nucleus in the brainstem. At six weeks post-operation, there was an obvious loss of axon bundles within the cuneate fasciculus sample area at level C3 (Fig. 6A), three spinal cord segments rostral to the injury.

Quantification of axonal loss within the cuneate fasciculus at cervical level C3 in rats that recovered proprioception (responders) and those that did not (nonresponders). Low power image showing the region of interest (ROI1) in (
Counting revealed a marked reduction in small (2–4 μm) axon bundles from 201 (±11) per sample area (ROI1) on the normal side to 75 (±12) on the operation side. There was also a reduction in large (4–10 μm) axon bundles, from 44 (±9) on the normal side to 25 (±13) after the surgical procedure. This corresponds to a 63% and a 43% reduction in the small and large axon bundles in the hOEC responder group. The extent and pattern of loss of axons in the same region of nonresponder hOEC rats was similar—68% and 65% loss of small and large axon bundles, respectively.
Examining the activation of microglia with Iba1+ immunostaining
To determine whether altered immune responses could account for differences in proprioception on the vertical climbing task, we examined microglial activation (by Iba1+ immunostaining) within the deep dorsal horn at cervical levels C7, C8 at six weeks post-operation. Both responder and nonresponder groups showed similar levels of Iba+ staining on their normal side (Fig. 7B,D). There was a substantial increase in the number of cells positively stained for Iba1, from 49 to 112 on the operation side (Fig. 7C) of responder rats. There was a similar although slightly attenuated increase in Iba1+ staining in the nonresponder group, from 46 cells to 94 cells after injury. In terms of morphology, it was apparent that after operation, microglia had enlarged processes and took up a larger area than those at baseline (refer to Fig. 7D). Preliminary studies showed CD68-positive stained macrophages at the injured dorsal horn six weeks post-operation; there was no detectable difference in terms of the distribution or quantity of macrophages between responder or nonresponder rats (unpublished data).
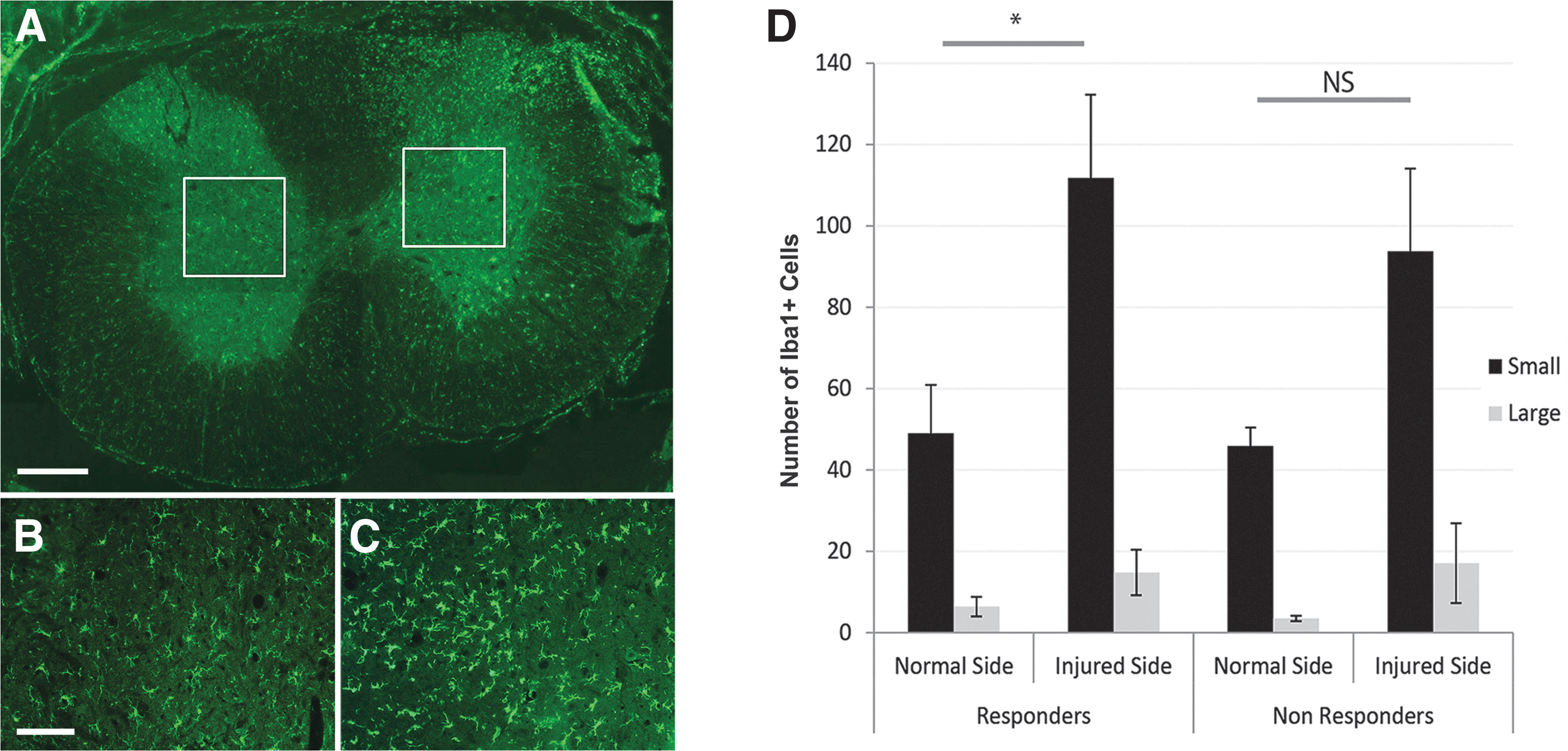
Quantification of microglial activation within the deep dorsal horn at cervical levels C7-C8 in rats that recovered proprioception (responders) and those that did not (nonresponders). Low power image shows the region of interest (ROI2) in (
Discussion
For the first time, in this study, OECs from human olfactory bulb tissue were mixed with collagen and transplanted into a rat model of spinal roots injury. This builds on a number of pre-clinical animal studies that found benefits from OEC transplantation and has culminated in their safety being confirmed in recent clinical trials. 25,26,28 We obtained and cultured cells from human olfactory bulb samples and mixed these hOECs with collagen gel. The cells were transplanted into a four dorsal root transection injury at the cervical level in rats. Half of the rats receiving hOEC transplants showed an improvement in proprioception as assessed on a vertical climbing task.
Optimizing hOEC transplants for cavity filling
Damage to the spinal cord often involves the progressive loss of tissue over time, which can result in cavitation at the injury site. 35 The present protocol of hOEC cell preparation cannot produce the quantity or composition of cells to fill a lesion cavity adequately on their own. In a clinical trial, Tabakow and colleagues 29 injected hOECs into the patient's sectioned spinal cord stumps rostral and caudal to a lesion cavity. The 11 mm gap between the injured ends was filled with four autologous peripheral nerve grafts. The limited mass of biopsy tissue yields far lower cell numbers than what is needed for repair, so the use of collagen may allow hOECs to distribute within a much larger volume. In this current study, mixing hOECs with collagen enabled cell expansion before transplantation where they distributed evenly in the gel, and many had formed elongated processes.
This method was suitable because hOECs were positioned optimally within the gel before transplantation, with early process elongation potentially enhancing their interaction with host astrocytes. For example, it has been shown that aligned poly-lactic acid microfibers provided a better scaffold for regenerating axons and astrocytes than randomly assorted fibers after a transection injury in rats. 36
The further benefit of using a gel is that it could avoid the numerous time-consuming cell injections that can damage the spinal cord. Although it has been shown in clinical studies in patients with complete spinal cord injuries that multiple spinal cord OEC-microinjections were safe and did not cause deterioration of the patient's neurological state, 25,27 –29 a delivery of OECs embedded within scaffolds and not as single cell suspension microinjections seems to be less invasive both for the spinal cord tissue and transplanted cells.
Transplanted hOECs were located predominantly as a concentrated aggregate at the lesion site, while a few other individual cells with elongated processes apparently had migrated away. These findings are similar to a study by Deng and coworkers 37 in which they injected human OECs (from the nasal lamina propria) into a rat spinal cord after a hemisection injury. In our current study, some small diameter axons were found alongside outgrowths of spinal cord astrocytic processes that have both been seen previously with rat OEC transplants 16,38 ; this suggests the human OECs have a similar effect, even if the host species is different.
Climbing performance
The vertical climbing test is an objective and reliable measure of forelimb proprioception; our previous work determined that transection of dorsal roots C6, C7, C8, and T1 led to simple, consistent, and permanent functional deficits of the upper limb. 33 In this and in previous studies, animals that failed to show any improvement by approximately two weeks post-operation tended to perform poorly for the remainder of the experiment (Collins A and Li Y, unpublished observations). For this reason, after analysis, we assigned rats as either responder or nonresponder based on their early post-injury climbing performance.
At the end of the six weeks study, half of the rats that received hOEC transplants (i.e., nine responders) had scored 30% higher in terms of climbing performance. They also had a foot fault score approximately two points lower than the control rats (injury alone). By assessing the magnitude of error of misplaced grasps—i.e., the foot fault score—we could detect subtle changes in proprioception rather than simply whether a paw grasp was successful or not.
In our previous electrophysiological study in which we examined transplants of rat olfactory bulb OECs in the same four dorsal root transection model, we found that 70% of rats showed some recovery of proprioception while the remaining 30% lacked any recovery (nonresponders). 16 Given the preliminary nature of this study, it is unsurprising that as many as half of the injured rats that received hOECs failed to show signs of recovery; potential reasons for this are discussed below.
Source of OECs
The anatomical source of OECs is a crucial factor in functional repair. We have shown previously that transplantation of cells cultured from rat olfactory bulb but not olfactory mucosa could reinstate proprioception in a climbing task. 39 In a subsequent clinical case study, a patient showed some functional improvement after autologous transplantation of olfactory bulb OECs to manage a thoracic level injury. 29 The observed clinical, neurophysiological, and radiological evidence of regeneration of descending and ascending axons from a central origin throughout the area of spinal cord reconstruction contrasted with clinical studies performed with olfactory mucosal OECs that showed either minimal 25 or no recovery from the transplantation. 26,28 Here, we have transplanted human olfactory bulb OECs into another species, the rat, to see for the first time whether olfactory bulb cells from one species can repair dorsal root injury in another.
Few studies have investigated transplants of human OECs in rat models of spinal cord injury. Olfactory bulb tissue from nonhuman primates has been used to show behavioral recovery after a T9/T10 level spinal transection in immunodeficient rats. 40 Other groups have used human OECs but not from the olfactory bulb; Gorrie and associates 41 transplanted human mucosa OECs into a rat spinal cord contusion model, and Kato and colleagues 42 transplanted human olfactory nerves into a rat model of demyelination. There was some functional recovery after the contusion injury and evidence of remyelination of rat axons in the other study.
Limiting factors that may account for partial recovery
A number of factors could explain why only half of rats (nine of 18) that received hOEC transplants in this current study showed recovery and why the best performance in the vertical climbing test at the end of the trial still resulted in 60% of grasps being misplaced.
First, although hOECs have been shown to have similar repair properties to rat OECs in terms of remyelination, their heterogeneity, morphological diversity, and lack of cell specific markers makes it difficult to compare fully OECs from these two different species. 43 –45
Second, the yield of hOECs in this study is lower than what would be obtained from a rat. This reduction is because of the long time (24 h) taken for tissue transportation (unpublished data). It should also be noted that these olfactory bulbs may have had pathomorphological changes before being obtained, such as intraparenchymal hemorrhages that were visible in some cases of OB microscopic dissection. This could have been caused by contusion injury to the frontal lobe and the olfactory bulb accompanying the skull base fracture or the presence of neoplasmatic infiltration in other cases. Also, interpersonal variability in the anatomy of the human olfactory bulb or previous exposure to toxic agents or infections could alter the condition of bulbar cells. 46 –48
Third, the proportions of rat OECs and ONFs used in our previous studies were approximately 50%, respectively. In our cellular scaffold, the proportion of p75+ OECs was close to 90%, and fibronectin deposition was low. This may have an effect on their repair function.
Fourth, this is the first study in which we have mixed OECs with a collagen gel—we assume this would not negatively impact on their repair properties but as yet this has not been explored fully. We do not know if hOECs that remain within the gel and those that migrate out have similar repair properties. We need to explore how to optimize the transplant biomaterial for repair. In particular, we must consider what proportion of hOECs, olfactory fibroblasts, and collagen gel characteristics are best for promoting repair.
Fifth, there are a number of technical aspects of the surgical procedure that may have led to differential functional recovery in the rats. For example, although care was taken to transplant a similar number of cells to each injury site, it is possible that some cells may not have remained at the injury site or may not have formed a complete bridge between the cut ends of the dorsal roots and the spinal cord or that fibrin glue may not have completely covered each portion of collagen gel. Examining tissue at the end of the study, we saw no apparent difference in cell survival within the responder or nonresponder groups of rats.
Responders vs. nonresponders: Quantification of axon loss and microglial activation
Rats that recovered proprioception (responders) and those that did not (nonresponders) showed similar grooming behavior in their home cage and had no locomotor deficits when exploring their cage (Collins, A and Li Y, unpublished observations). The two groups could only be distinguished when their forelimb grasping was assessed on a vertical cage climbing test. Histological examination could not uncover any difference in terms of location or survival of hOECs, which could explain the disparity in climbing performance. In both groups, we saw the collagen gel maintain its structure at the injury site, with hOECs remaining in the gel and others in a cluster within the spinal cord; the elongated spindle-like morphology of these cells was also similar in responder and nonresponder rats. Very few OEC processes were seen crossing the injury site into the spinal cord; in the case of those that did, the axons traveled with them at the entry zone.
We know from our previous electrophysiological study that showed responder rats that had received bulb OECs had partial recovery of an afferent volley through the transplanted dorsal root entry zone, which ascended rostrally up the dorsal columns to the cuneate nucleus. 16 The transplants may have preserved better or limited the secondary damage to axons of the ascending dorsal column to enable this attenuated afferent transmission. In the present study, we therefore compared responder and nonresponder rats in terms of axon loss within the cuneate fasciculus of cervical level C3.
George and Griffin 49 have described the proximo-distal spread of axonal degeneration, which is known to begin approximately one day after dorsal root rhizotomy and proceed as quickly as 3 mm per hour. Axons within the cervical cuneate fasciculus are known to transmit, among other afferents, skin and tactile proprioception from the upper half of the body, which is relevant given the nature of the vertical climb test used in the present study. 50 Although we could not identify any quantifiable difference in axon loss between responder and nonresponder groups, this method is likely not sensitive enough to detect the relatively few repaired axons it would require for transmission through to the cuneate nucleus.
Early, neuroprotective actions of OECs have been suggested to account for recovery of appropriate forelimb responses to thermal stimuli in a recent dorsal root injury study. 51 Although short, calcitonin gene-related peptide-positive fibers were observed at the injured dorsal root entry zone of rats that received OEC transplants, there was no accompanying increase in sensitivity to heat or cold stimuli. We did not examine responses to painful stimuli in this current study, but future work should investigate whether hOEC transplants in a rat model of dorsal root avulsion injury can prevent abnormal sensory responses in the same way as rat OEC transplants. 51 This would have obvious implications for their use in a clinical setting.
The interaction between the immune system and the damaged spinal cord is now viewed as highly relevant. 52,53 Microglia have been implicated heavily in generating and maintaining pain states after spinal cord injuries, as well as modulating levels of inflammation. 54,55 By choosing to examine microglial activation at the deep dorsal horn, we were able to see whether improved proprioception was associated with altered immune system responses. In this case, there was no obvious association: both responder and nonresponder groups of animals showed a very similar activation of microglia at six weeks post-operation. We cannot rule out the possibility that administering cyclosporine daily throughout the study may have obscured or modified any hOEC transplant related effects on immune system function.
Footnotes
Acknowledgments
This work was supported by funds from the UK Stem Cell Foundation, the Nicholls Spinal Injury Foundation, and the Polish National Centre of Science in the years 2013–2016 (study number GR-797/NCN/2013 and 2012/06/M/NZ4/00138).
Author Disclosure Statement
No competing financial interests exist.