
Abstract
Vepoloxamer is an amphipathic polymer that has shown potent hemorrheologic, cytoprotective, and anti-inflammatory effects in both pre-clinical and clinical studies. This study was designed to investigate the therapeutic effects of vepoloxamer on sensorimotor and cognitive functional recovery in rats after traumatic brain injury (TBI) induced by controlled cortical impact. Young adult male Wistar rats were randomly divided into the following groups: 1) sham; 2) saline; or 3) vepoloxamer. Vepoloxamer (300 mg/kg) or saline was administered over 60 min via intravenous infusion into tail veins starting at 2 h post-injury. Sensorimotor function and spatial learning were assessed using a modified neurological severity score and foot fault test, and Morris water maze test, respectively. The animals were sacrificed 35 days after injury and their brains were processed for measurement of lesion volume and neuroinflammation. Compared with the saline treatment, vepoloxamer initiated 2 h post-injury significantly improved sensorimotor functional recovery (Days 1–35; p < 0.0001) and spatial learning (Days 32–35; p < 0.0001), reduced cortical lesion volume by 20%, and reduced activation of microglia/macrophages and astrogliosis in many brain regions including injured cortex, corpus callosum, and hippocampus, as well as normalized the bleeding time and reduced brain hemorrhage and microthrombosis formation. In summary, vepoloxamer treatment initiated 2 h post-injury provides neuroprotection and anti-inflammation in rats after TBI and improves functional outcome, indicating that vepoloxamer treatment may have potential value for treatment of TBI. Further investigation of the optimal dose and therapeutic window of vepoloxamer treatment for TBI and the mechanisms underlying beneficial effects are warranted.
Introduction
T
Poloxamer 188 (P-188) is an amphiphilic polyethylene-polypropylene-polyethylene tri-block copolymer that is reported to seal membranes and restore plasma membrane integrity in cells damaged by mechanical and electrical injury, and in experimental models of muscular dystrophy, heart failure, and neurodegenerative disorders. 5 P-188 significantly inhibits apoptosis and prevents necrosis in an in vitro model of neuronal injury. 6 P-188 also has been reported to attenuate TBI-induced brain injury by reducing blood–brain barrier damage, brain edema, apoptosis, and mitochondrial dysfucntion. 7,8 P-188 also attenuated TBI-induced motor and cognitive dysfunction in mice after TBI. 8 These studies suggest that P-188 might serve as a potential drug for treatment of TBI.
In early clinical studies with P-188, elevations in serum creatinine were observed, especially in elderly subjects or those with pre-existing renal dysfunction. 9,10 Vepoloxamer is a purified form of P-188 where impurities associated with renal dysfunction have been removed. 9,10 Clinical studies investigating vepoloxamer in patients with sickle cell disease demonstrated it was well tolerated and the elevated creatinine observed with commercially available P-188 was not observed. 11 With the notable exception of renal dysfunction, vepoloxamer exhibits the same pharmacological actions as unpurified P-188. 10,12,13 Based on preliminary studies with P-188, vepoloxamer may be a promising treatment for TBI. In the current study, we report the effects of vepoloxamer on motor and cognitive functional recovery in an animal model of TBI, and the potential mechanisms underlying its beneficial effects.
Methods
All experimental procedures were approved by the Institutional Animal Care and Use Committee of Henry Ford Health System. To prevent potential biases of performance and detection, the persons who performed the experiments, collected data, and assessed outcome were blinded throughout the course of the experiments and were unaware of the treatment allocation.
Animal model and experimental groups
A controlled cortical impact (CCI) rat model of TBI was utilized to induce moderate focal brain injury in the present study. 14 After an appropriate period (1 week) of quarantine and acclimation, adult male Wistar rats weighing 326 ± 14 g (2–3 months old) were anesthetized with intraperitoneal (i.p.) ketamine/xylazine (100/10 mg/kg). Rectal temperature was maintained at 37 ± 5°C using a feedback-regulated water-heating pad. Rats were placed in a stereotactic frame. Two 10-mm-diameter craniotomies were performed adjacent to the central suture, midway between lambda and bregma. The second craniotomy allowed for lateral movement of cortical tissue. The dura mater was kept intact over the cortex. Cortical injury was delivered by impacting the left cortex (ipsilateral cortex) with a pneumatic piston containing a 6-mm diameter tip at a rate of 4 m/sec and 2.5 mm of compression. Velocity was measured with a linear velocity displacement transducer.
Young adult male Wistar rats were randomly divided into the following groups: 1) sham (no injury; n = 7); 2) saline (TBI rats treated with intravenous saline administered 2 h post-injury; n = 8); or 3) vepoloxamer (TBI rats treated with intravenous vepoloxamer; 300 mg/kg) administered 2 h post-injury; n = 8). Vepoloxamer or saline was administered via intravenous infusion into tail veins over 60 min starting at 2 h post-injury. An additional set of rats (sham, saline, and vepoloxamer, n = 5/group for each time-point) were used for bleeding time assay, which was performed 1 h and 24 h post-drug treatment using a tail transection bleeding test. 15 Rectal temperature of anesthetized animals was maintained at 37°C using a feed-back heating pad. Rat tail tip was cleaned with betadine and 70% alcohol, and transected 5 mm from the tip with a sterile surgical scalpel blade. The resultant wound was gently blotted with Whatman filter paper until bleeding stopped. Time from transection to cessation of bleeding was recorded as bleeding time. Each animal was monitored for 20 min even if bleeding ceased, in order to detect any re-bleeding. Animals were returned to their cages after bleeding had stopped.
Modified neurological severity score test
To investigate neurological functional outcome, the modified neurological severity score (mNSS) test was performed. 16 The test was carried out on rat pre-injury and at 1, 7, 14, 21, 28, and 35 days after TBI. mNSS is a composite of the motor (muscle status, abnormal movement), sensory (visual, tactile, and proprioceptive), and reflex tests and has been used in previous studies. 17 Neurological function was graded on a scale of 0 to 18 (normal score, 0; maximal deficit score, 18). One point is awarded for each abnormal behavior or for lack of a tested reflex; thus, the higher the score, the more severe the injury.
Foot fault test
To evaluate sensorimotor function, the foot fault test was carried out prior to TBI and at 1, 7, 14, 21, 28, and 35 days after TBI. The rats were allowed to walk on a grid. With each weight-bearing step, a paw might fall or slip between the wires, and if this occurred, it was recorded as a foot fault. 18 A total of 50 steps were recorded for the right forelimb.
Morris water maze test
To measure spatial learning impairments, an updated version of the Morris water maze (MWM) test was used. 19 The MWM test was performed during the last 5 days (Days 31–35) before they were sacrificed. A dark blue swimming pool (1.8 m in diameter) was located in a large room with many clues external to the maze (e.g., pictures on the walls, lamps and a camera on the ceiling), which were visible from the pool and presumably used by the rats for spatial orientation. The position of the cues remained unchanged throughout the experiment. Data collection was automated using the HVS Image 2020 Plus Tracking System (US HVS Image, San Diego, CA), as described previously. 20 During set up of the tracking system, the swimming pool was subdivided into four equal quadrants formed by imaging lines. At the beginning of each trial, the rat was placed at one of four fixed starting points, randomly facing toward a wall (designated North, South, East, and West) and allowed to swim for 90 sec or until it found the platform which was transparent and invisible to animals. If the animal found the platform by spatial navigation, it was allowed to remain on it for 10 sec. If the animal failed to find the platform within 90 sec, it was placed on the platform for 10 sec. At each testing interval, animals were tested with four trials per day. Throughout the test period, the platform was located in the northeast (NE) quadrant 2 cm below water in a randomly changing position, including locations against the wall, toward the middle of the pool, or off-center but always within the target quadrant.
Data collection
If the animal was unable to locate the platform within 90 sec, the trial was terminated and a maximum score of 90 sec was assigned. If the animal reached the platform within 90 sec, the percentage of time traveled within the NE (correct) quadrant was calculated relative to the total amount of time spent swimming before reaching the platform and employed for statistical analysis. The latency to find the hidden escape platform also was recorded and analyzed. The advantage of this protocol is that rodents should find the platform purely and extensively by reference to the extra-maze spatial cues, which improves the accuracy of evaluating spatial performance in the MWM. 19
Tissue preparation and measurement of lesion volume
On Day 35 after TBI, rats were anesthetized with ketamine (80 mg/kg, i.p.) and xylazine (13 mg/kg, i.p.) and perfused transcardially with saline solution, followed by 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS), pH 7.4. Rat brains were removed and post-fixed in 4% paraformaldehyde for 2 days at room temperature. Using a rat brain matrix (Activational Systems Inc., Warren, MI), each forebrain was cut into 2-mm thick coronal blocks for seven equal blocks from bregma 5.2 mm to bregma −8.8 mm per animal. The tissues were embedded in paraffin and a series of 6-μm thick slides were cut from each block in the coronal plane and stained with hematoxylin and eosin (H&E). For lesion volume measurement, one 6-μm thick section from each of seven coronal blocks was traced by a microcomputer imaging device (Imaging Research, St. Catharine's, Ontario), as described previously. 21 Briefly, the ipsilateral and contralateral sides were traced on each section to obtain the area excluding ventriculomegaly and subcortical tissue loss, and this was multiplied by the known distance between sections to obtain the volume. Then the volume of the ipsilateral hemisphere was subtracted from that of the contralateral side to obtain the volume of total brain tissue loss. The cortical lesion volume was expressed as a percentage calculated by [(contralateral hemisphere volume – ipsilateral hemisphere volume)/(contralateral hemisphere volume) × 100%]. 22
Immunohistochemistry
To evaluate the anti-inflammatory effects of vepoloxamer, cluster of differentiation 68 (CD 68; marker for microglia/macrophages) immunostainings and glial fibrillary acidic protein (GFAP; marker for astrocytes) immunostainings were performed using paraffin-embedded sections (6 μm thick), which were deparaffinized and rehydrated. Antigen retrieval was performed by boiling sections in 10 mM citrate buffer (pH 6.0) for 10 min. After washing with 0.1 M PBS, sections were incubated with 0.3% H2O2 in PBS for 10 min, blocked with 1% bovine serum albumin containing 0.3% Triton-X 100 at room temperature for 1 h, and incubated with mouse anti-CD68 (1:200; AbD Serotec, Kidlington, UK), rabbit anti-rat thrombocytes (1:1000; Accurate Chemical, Westbury, NY) and polyclonal rabbit anti- GFAP (1:1000; Dako, Denmark) at 4°C overnight. For negative controls, primary antibodies were omitted. After washing, sections were incubated with biotinylated anti-mouse or anti-rabbit antibodies (1:200; Vector Laboratories, Inc., Burlingame, CA) at room temperature for 30 min. After an additional washing, sections were incubated with an avidin-biotin-peroxidase system (ABC kit; Vector Laboratories, Inc.), visualized with diaminobenzidine (Sigma), and counterstained with hematoxylin. 23
Cell counting and quantitation
CD68+ microglia/ macrophages, and GFAP+ astrocytes were counted in the lesion boundary zone, corpus callosum, and the dentate gyrus, four slides from each brain. The fields of interest were digitized under the light microscope (Nikon, Eclipse 80i) at a magnification of either × 200 or × 400 using a CoolSNAP color camera (Photometrics) interfaced with MetaMorph image analysis system (Molecular Devices, Sunnyvale, CA), as previously described. 24 Five fields of view in the lesion boundary zone from the epicenter of the injury cavity (bregma −3.3 mm), 2 fields of view in the corpus callosum and 9 fields of view in the ipsilateral dentate gyrus were counted in each section. In the present study, one blinded observer performed the cell counting in all brain sections. Vascular structures containing positive signals for thrombocytes are considered as vessels containing thrombosis, and data are presented as the density (vessels/mm2) of vessels with thrombosis, regardless of vessel size. The thrombosis index, defined as the number of microthrombi per total vessel number in the sample, was determined on H&E-stained coronal brain sections. Brain hemorrhage was measured 24 h after TBI by using H&E staining under light microscopy in all animals. Petechial hemorrhage, defined as a cluster of red blood cells outside of the lumen of blood vessels, was measured on seven H&E-stained coronal sections with a Global Laboratory Image analysis program (Data Translation). Each H&E-stained coronal section was evaluated, and the distribution of hemorrhage was recorded under a × 20 objective. The area of hemorrhage (μm2) was calculated by tracing the areas on the computer screen, and a summation of the area of hemorrhage from each section was presented as the total area of hemorrhage. 25
Statistical analysis
All data are presented as the means with standard deviations. The sham group was used to study the severity of TBI. Repeated analysis of covariance was used to study the treatment effect on functional recovery with consideration that the treatment effect might vary over time. Generalized linear mixed models using proc GLIMMIX in SAS 9.4 was performed. The analysis would start testing for treatment by time interaction, followed by testing the overall treatment effect if no interaction was detected with p value <0.05 or testing the treatment effect at each time-point if otherwise. A one-way analysis of variance followed by post hoc Tukey's tests was used to compare the differences in bleeding time, microthrombosis, and hemorrhage area between the vepoloxamer-treated, saline-treated, and sham groups. Two-sample t-test was used to study the treatment difference on lesion volume, collected at Day 35 after sacrifice. The eight animals per group with two groups for the treatment effect would yield an effect size of 1.51 with power of 80%.
Results
Vepoloxamer administration significantly decreases the volume of total brain tissue loss in rats after TBI
To explore the neuroprotective effect of acute treatment with vepoloxamer on tissue loss, the lesion volume was measured at 35 days after TBI. Compared with the saline treatment group, acute treatment (2 h after TBI) with vepoloxamer significantly decreased the lesion volume (Fig. 1; p < 0.05).
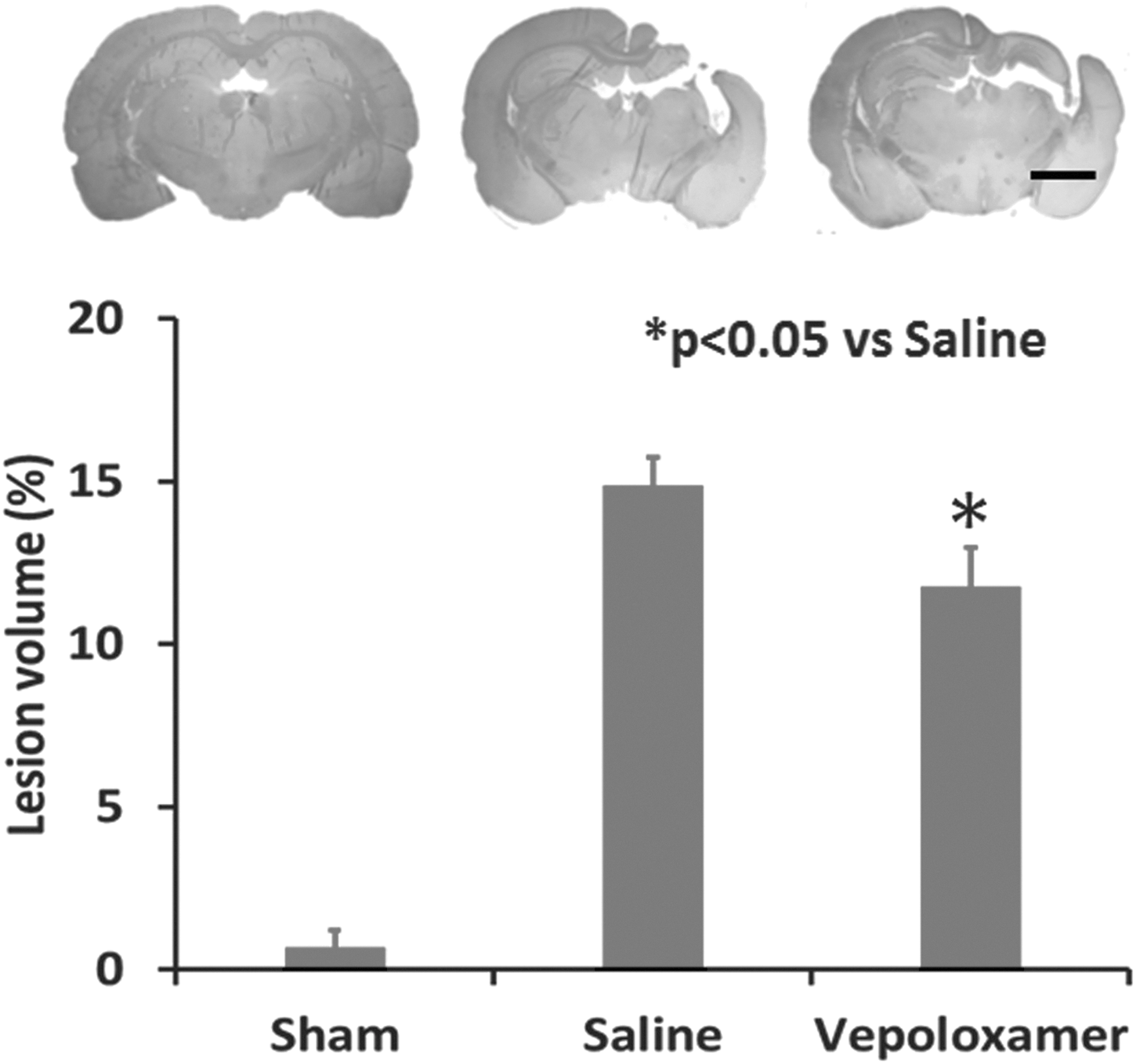
Vepoloxamer administration significantly decreases the volume of total brain tissue loss in rats after traumatic brain injury (TBI). Lesion volume was examined at Day 35 after TBI. Hematoxylin and eosin staining. Scale bar = 3 mm. Data represent mean ± standard deviation. n = 7–8 rats/group.
Vepoloxamer administration significantly enhances spatial learning in rats after TBI
Vepoloxamer administration significantly enhances spatial learning in rats after TBI. The MWM test is a sensitive and reliable index of hippocampal injury. 19 The higher the percentage of time the rats spend in the correct quadrant where a platform is submerged, the better the spatial learning function. The shorter the time for the rats to find the platform, the better the memory function. Interaction of group by day is significant on both water maze time percentage and latency (p < 0.0001). Treatment effect on water maze time percentage and latency was significant from Day 32 (p < 0.0001). Specifically, from Days 32–35, the percentage of time the rats spent in the correct quadrant increased significantly after vepoloxamer treatment, compared with the saline treatment group (Fig. 2; p < 0.0001). From Days 33–35, the time that rats found the platform decreased significantly after vepoloxamer administration, compared with the saline treatment group (Fig. 2; p < 0.0001).
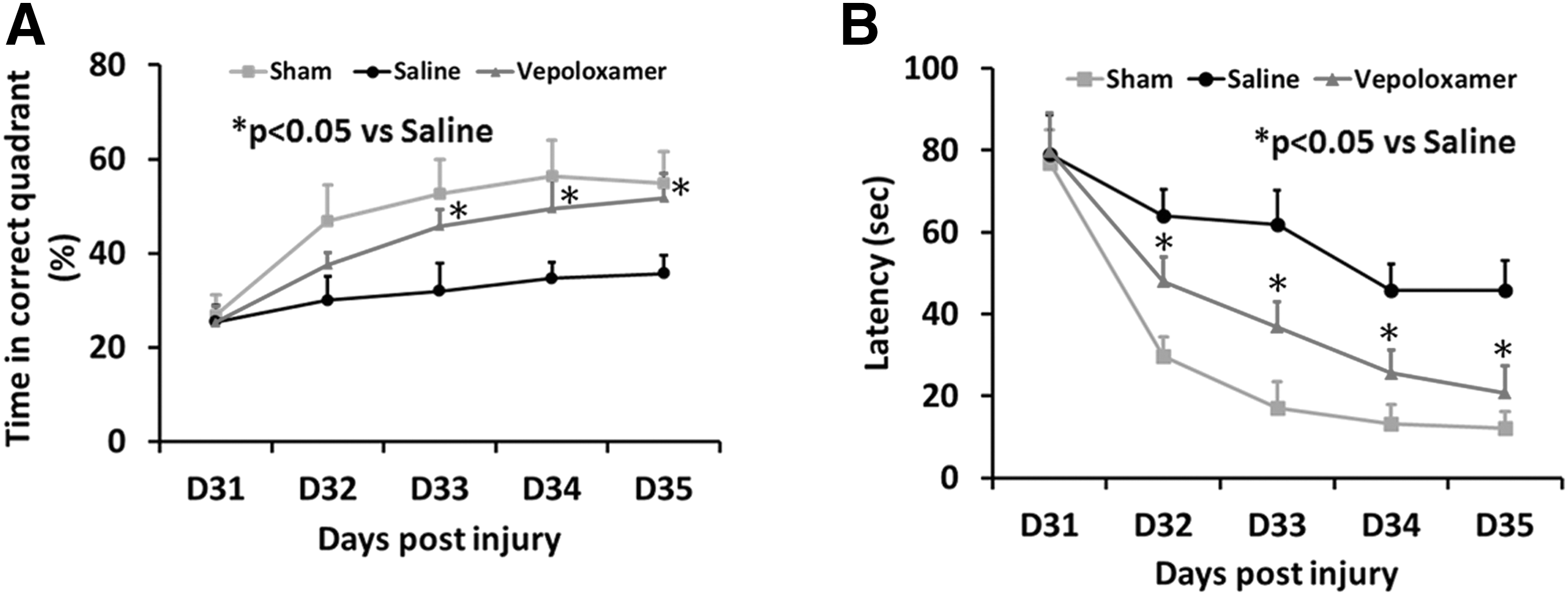
Vepoloxamer administration significantly improves spatial learning in the Morris water maze test.
Vepoloxamer administration significantly promotes sensorimotor functional recovery in rats after TBI
For sensorimotor functional assessment, foot fault steps and mNSS score were recorded. An increased mNSS score is associated with a more severe injury. Interaction of group by day is significant on foot fault (p < 0.0001) and mNSS (p = 0.0002). Treatment effect on mNSS and foot fault was significant at each day (p < 0.0001). Specifically, vepoloxamer treatment 2 h post-TBI significantly improved sensorimotor functional recovery by reducing numbers of foot fault steps (p < 0.0001) and mNSS scores (p < 0.0001) from Day 1 to Day 35 after TBI, compared with the saline treatment group (Fig. 3). Compared with sensorimotor deficits at Day 1, the foot fault steps and mNSS scores were significantly decreased starting from Day 7, which suggests that spontaneous functional recovery occurred over time after TBI.
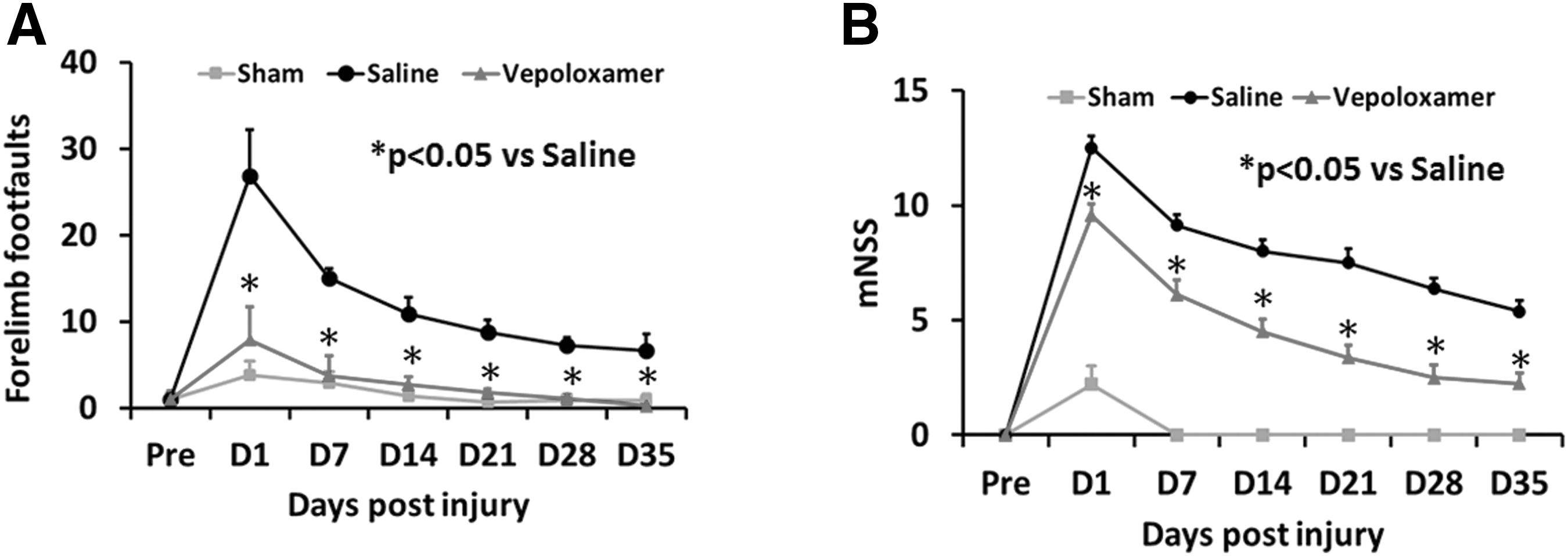
Vepoloxamer administration significantly improves sensorimotor function measured by right forelimb foot fault test and modified neurological severity score (mNSS) in rats after traumatic brain injury. The line graph shows that vepoloxamer treatment significantly reduces forelimb foot faults at Days 1–35
Vepoloxamer administration significantly decreases the number of brain astrocytes in rats after TBI
GFAP+ areas were present in TBI rats after saline treatment (Fig. 4). After vepoloxamer administration, GFAP+ areas were significantly decreased in the cortex, corpus callosum and dentate gyrus, compared with the saline treatment group (Fig. 5; p < 0.05).
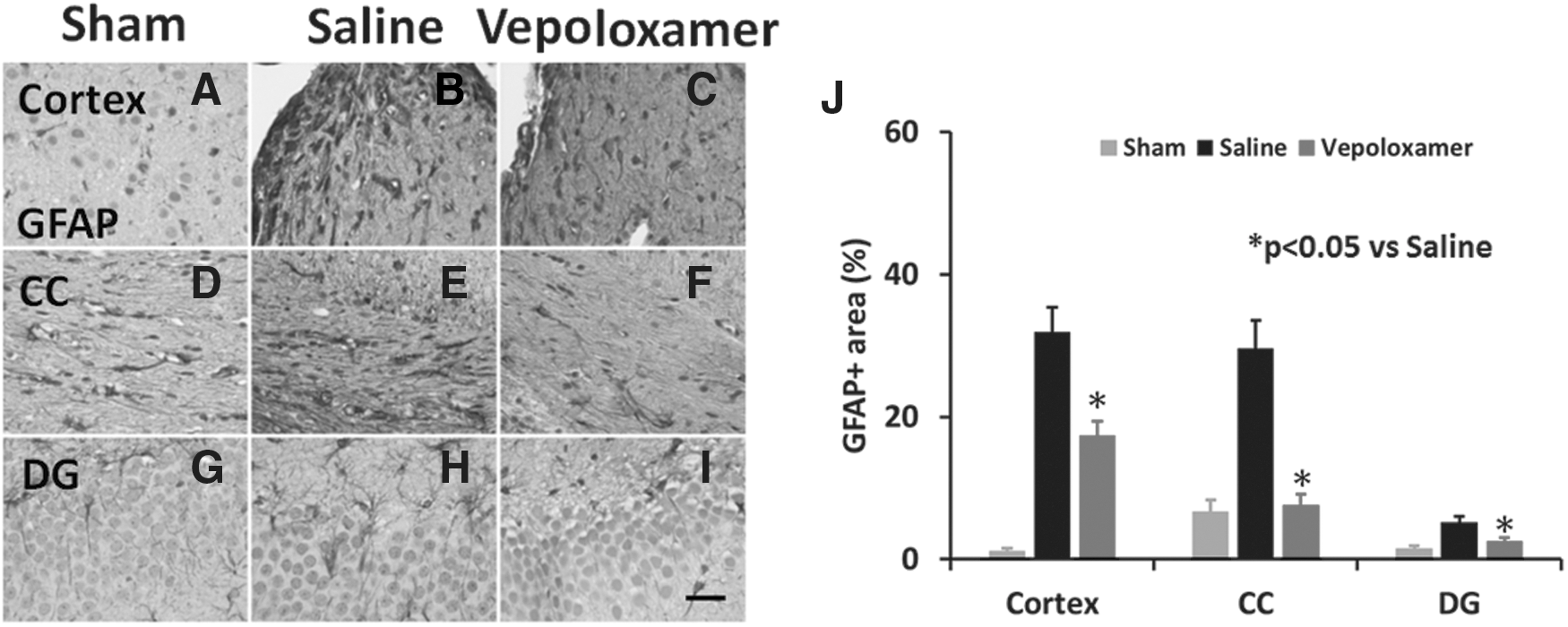
Vepoloxamer administration significantly decreases the number of glial fibrillary acidic protein (GFAP)+ astrocytes in the ipsilateral cortex, corpus callosum, and dentate gyrus in rats after traumatic brain injury (TBI). GFAP staining was performed to detect activation of astrocytes 35 days after TBI. Some expression of GFAP was observed in brain regions of sham animals (
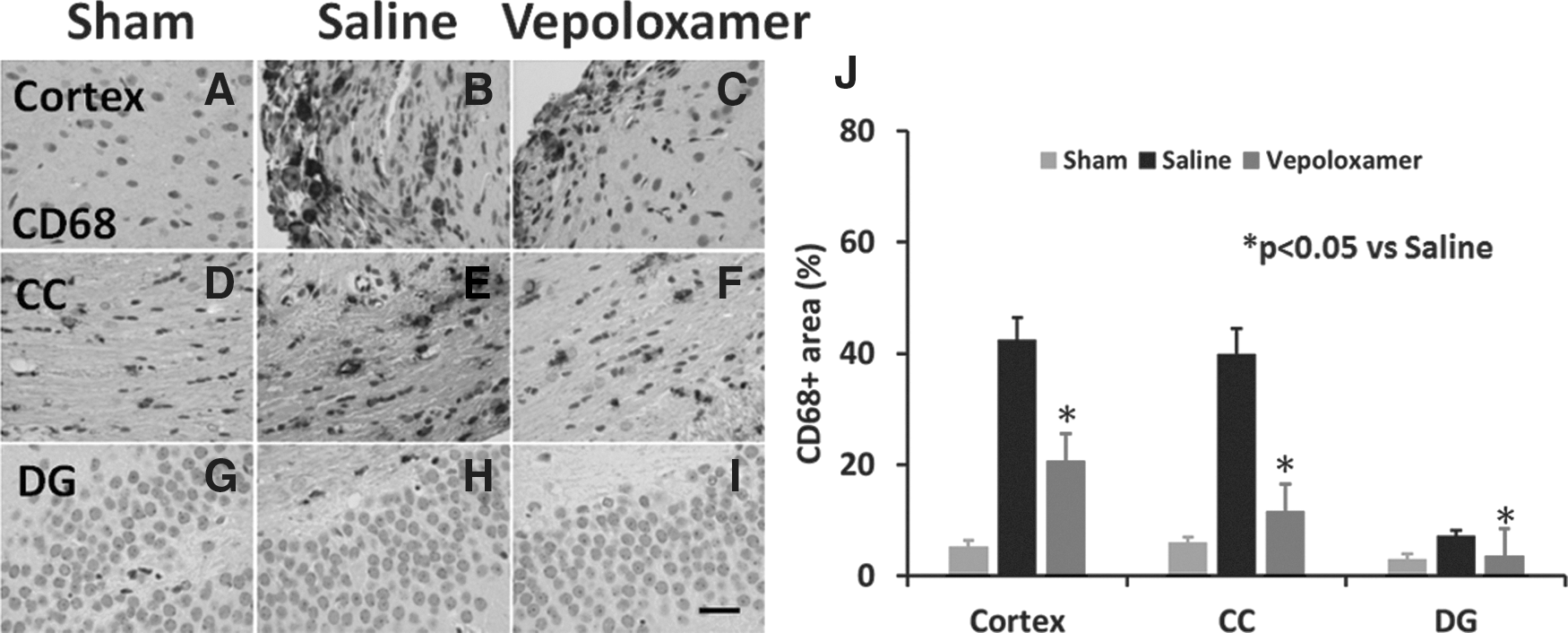
Vepoloxamer administration significantly decreases the number of cluster of differentiation 68 (CD68+ microglia/macrophages in the ipsilateral cortex, corpus callosum, dentate gyrus in rats after traumatic brain injury (TBI). CD68 staining was performed to detect microglia/macrophages 35 days after TBI. Some expression of CD68 was observed in corpus callosum
Vepoloxamer administration significantly decreases the number of brain microglia/macrophages in rats after TBI
There were abundant CD68+ cells in the cortex and corpus callosum of TBI rats treated with saline (Fig. 5). After vepoloxamer administration, CD68+ cells were significantly decreased in the cortex, corpus callosum, and dentate gyrus, compared with the saline treatment group (Fig. 4; p < 0.05). In the sham group, only a few scattered CD68+ cells were evident in the corpus callosum (Fig. 5).
Vepoloxamer administration normalizes the bleeding time in rats after TBI
TBI rats treated with saline had a decreased bleeding time at 1 h post-treatment (i.e., 3 h post-injury, 173.0 ± 8.4 sec; p < 0.0001) and an increased bleeding time at 22 h post-treatment (i.e., 24 h post-injury, 264.8 ± 14.9 sec; p = 0.0122), compared with sham controls (238.2 ± 10.9 sec). Vepoloxamer treatment significantly increased the bleeding time at 1 h post-treatment (252.70 ± 7.8 sec; p < 0.0001), compared with saline treatment but there was no significant difference, compared with sham controls (p = 0.0508). Vepoloxamer treatment did not alter the bleeding time at 22 h post-treatment (225.8 ± 7.9 sec; p = 0.0738), compared with sham controls but significantly reduced the bleeding time, compared with saline treatment (p = 0.0009).
Vepoloxamer administration reduces brain microthrombosis and hemorrhage in rats after TBI
TBI increased brain hemorrhage in the injured cortex, compared with sham controls (p < 0.0001), while vepoloxamer significantly reduced the area of hemorrhage (Fig. 6; p = 0.0197). Microthrombosis was identified by thrombocyte immunohistochemical staining at Day 1 after TBI (Fig. 7). TBI increased brain microthrombosis formation, compared with sham controls (p < 0.0001), while vepoloxamer significantly reduced microthrombosis formation (Fig.7; p = 0.0002).
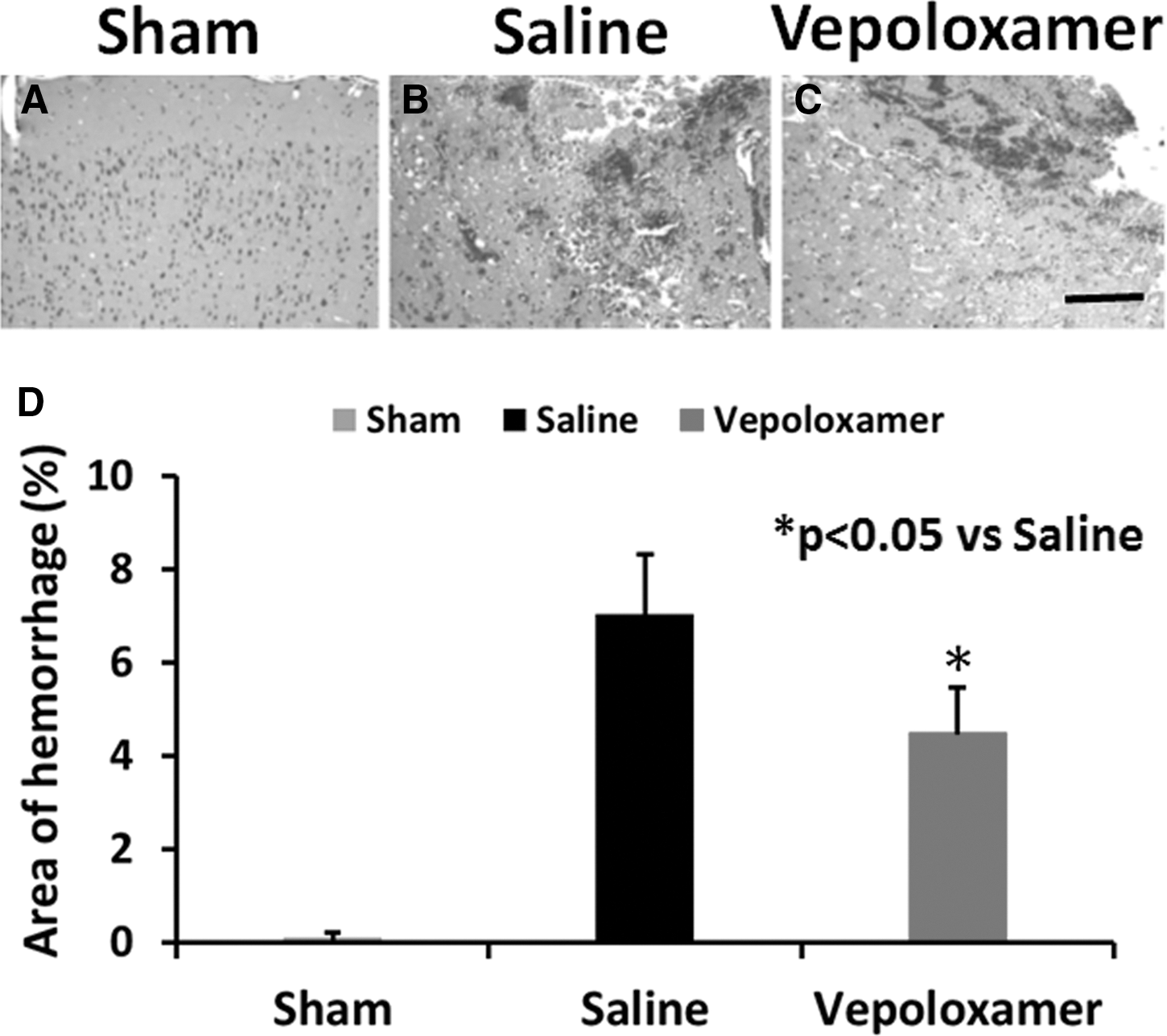
Vepoloxamer administration significantly decreases the area of brain hemorrhage in rats after traumatic brain injury (TBI). Hematoxylin and eosin staining was performed to detect hemorrhage. There was no hemorrhage detected in sham animals (
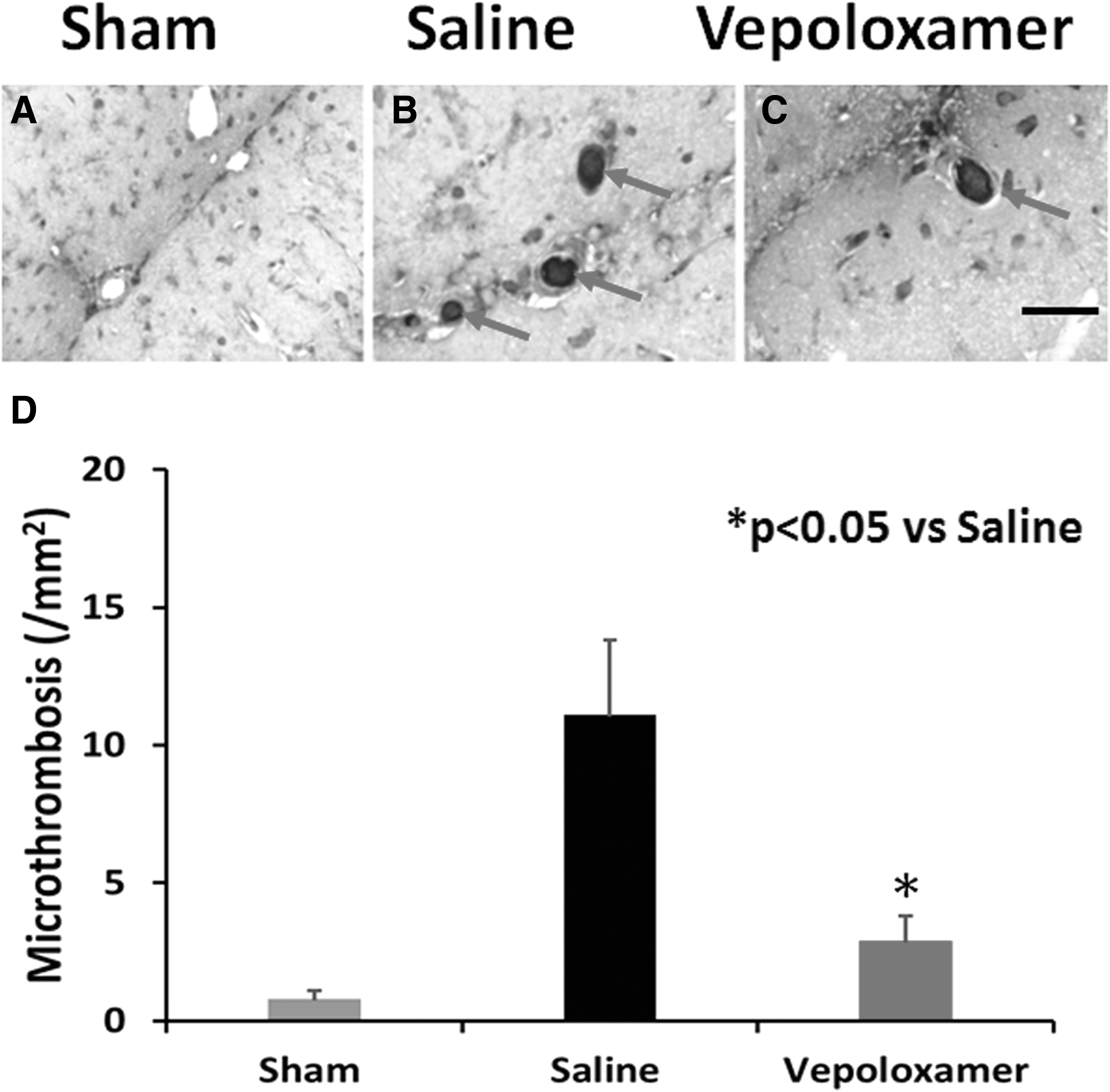
Vepoloxamer administration significantly decreases brain microthrombosis formation in rats after traumatic brain injury (TBI). Thrombocyte staining was performed to detect microthrombosis. These was no microthrombosis detected in sham animals (
Vepoloxamer administration does not affect body weight in rats after TBI
There was no significant difference in body weight detected between TBI rats treated with saline and vepoloxamer and sham controls. In general, all animals gained body weight over time (Fig. 8).
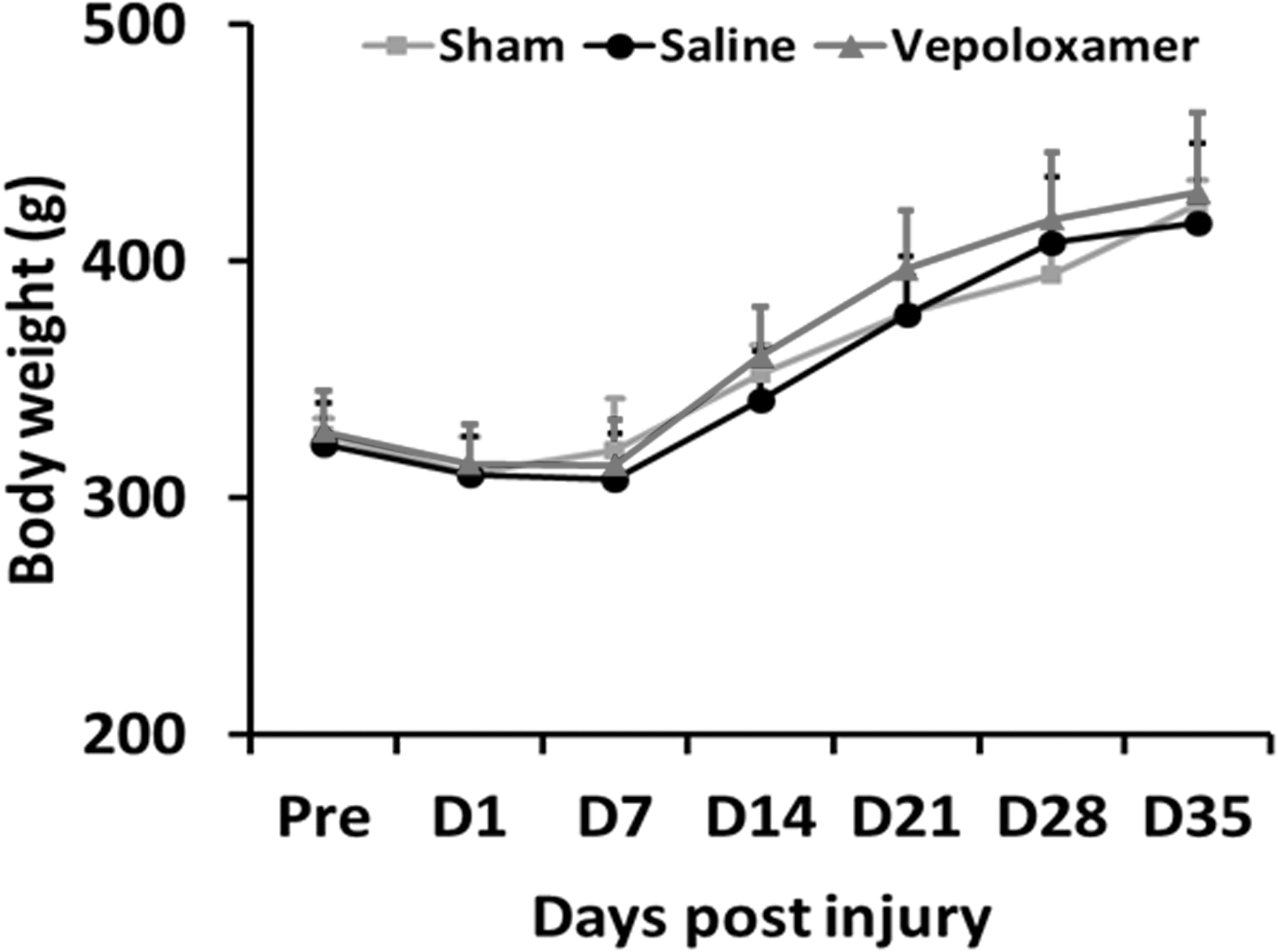
Vepoloxamer administration does not change body weight in rats after traumatic brain injury (TBI). There was no significant difference in body weight change between TBI rats treated with saline and vepoloxamer. All animals gained body weight over time. Data represent mean ± standard deviation. n = 7–8 rats/group.
Discussion
This study demonstrates that acute post-injury vepoloxamer administration: 1) decreases tissue loss of brain after TBI; 2) promotes functional recovery, including sensorimotor function and cognitive function, after TBI; 3) has an anti-inflammatory effect on microglia/macrophages and astrocytes; 4) normalizes the bleeding time, and reduces brain hemorrhage and microthrombosis formation; and 5) is well tolerated, as indicated by the absence of a significant difference in body weight between the saline, vepoloxamer, and sham control groups.
Vepoloxamer treatment initiated 2 h post-injury provides neuroprotection by reducing lesion size in rats after TBI. Previous studies showed that P-188 is capable of resealing several types of damaged cell membrane and protecting neurons after TBI by suppressing apoptosis. 7,26 –29 An in vitro study, using an in vitro cell shearing device (VCSD) to induce injury revealed that P-188 treatment protects neurons against VCSD-induced mitochondrial and lysosomal membrane damage. 21 P-188 also reduced neuronal cell death after TBI by regulating the MAPK pathway with acute activation of ERK1/2, JNK1/2, and p38. 6 Preventive treatment with P-188 (30 min before injury) reduced neural cell death and improved motor and cognitive function in a mouse model of TBI. 8 Further, in a stroke model, P-188 increased blood flow in areas of severe or moderate ischemia but had little effect in areas with mild or no ischemia. 30 It is possible that these vepoloxamer-induced actions account for the neuroprotection and cognitive and motor function recovery observed in our study.
Astrocytes are vital for the normal functioning of the brain by providing nutrients to support neuron viability, releasing factors that aid formation of synapses, as well as many other important functions. 31 Reactive astrocytes have both beneficial and detrimental effects by releasing anti-inflammatory and pro-inflammatory factors after TBI. 32 –34 Our data show that vepoloxamer improves sensorimotor and cognitive function, with a significant decrease in the GFAP+ astrocyte activation, suggesting that astrocyte activation may play an important role in functional impairment in rats after TBI. Astrocytes, together with microglia, can release pro- and anti-inflammatory cytokines, free radicals and antioxidants. 35,36 There is still debate whether activated microglia/macrophages protect neuron cells or further damage them. 37 However, in our current study, activated astrocytes and microglia/macrophages were dramatically increased in the saline treatment group and significantly decreased after vepoloxamer treatment. Further, previous studies indicate that the anti-inflammatory effects of therapeutic treatments are critical for functional recovery after TBI, 38 –41 which supports the hypothesis that the anti-inflammatory effect of vepoloxamer possibly contributes to the functional recovery after TBI in rats.
There are several limitations to this study. We only sacrificed animals at 35 days after TBI and measured lesion volume, GFAP+ astrocytes, and CD68+ microglia/macrophages in the brain at this single time-point. To obtain their dynamic profiling, measurements at multiple time-points are needed, especially at early time-points after injury. In addition, although our data indicate that activation of astrocytes and microglia/macrophages may contribute to functional deficits, the precise mechanisms underlying vepoloxamer-neuroprotection after TBI largely remain elusive. The reduced brain tissue damage may elicit less inflammation, as measured by GFAP+ and CD68+ cells at 35 days post-injury, in addition to a direct effect of vepoloxamer on neuroinflammation. To further investigate the molecular mechanisms underlying vepoloxamer-mediated neuroprotection, it is warranted to determine whether treatment alters the dynamic profiling of pro-inflammatory and anti-inflammatory cytokines in the brain after TBI at various time-points. For obtaining optimal dosing, dose–response and therapeutic window studies for vepoloxamer treatment of TBI are required. Vepoloxamer also may provide beneficial effects by reducing blood-brain-barrier damage in the acute phase after injury and by promoting neurovascular and axonal remodeling in the subacute phase after injury, measurements that we did not perform.
Dansdill and colleagues investigated drug interactions between tissue plasminogen activator (tPA) and vepoloxamer in a rat internal jugular vein thrombosis model, and demonstrated that the effect of tPA was facilitated by vepoloxamer but no rat exhibited spontaneous clot lysis with either saline or vepoloxamer. 13 A clinical study for evaluating the safety and efficacy of adjunctive therapy with P-188 at concentrations up to 300 mg/kg (a dose similar to one used in the present study) in patients with acute myocardial infarction receiving thrombolytic therapy with tPA demonstrated that there was no difference in the incidence of major or minor bleeding. 42 Our present study demonstrates that TBI reduced the bleeding time at 3 h post-injury but increased the bleeding time at 24 h post-injury, indicating that TBI alters the status of coagulation in rats. This is consistent with the report that TBI-associated coagulopathy often follows a course of transition from hypercoagulable to hypocoagulable state. 43 Vepoloxamer treatment initiated 2 h post-injury did not increase the bleeding time, compared with sham controls, but normalized the bleeding time measured at 3 h and 24 h post-injury, compared with saline treatment. These findings suggest that vepoloxamer treatment did not increase bleeding time but reduced the brain tissue loss in rats after TBI, indicating that vepoloxamer treatment initiated 2 h post-injury has no increased risk for intracranial hemorrhage in TBI with contusion. Early treatment of vepoloxamer restores coagulation after TBI by reducing brain thrombosis formation.
Our previous study suggests that brain thrombosis starts 1–4 h, peaks at 1–3 days, then declines at 8 and 15 days in rat after CCI-TBI. 44 A recent study demonstrates that microthrombi start to form in the parenchymal microcirculation within the first few hours and occlude more than 60% of all cerebral vessels 24 h in mice after CCI-TBI. 45 It is reported that microthrombosis formation in the brain after TBI is associated with hemodynamic depression, and resultant ischemic damages within areas surrounding the contusion. 46 Patients with moderate TBI are at serious risk of developing brain intravascular microthrombosis 47 and there is a strong link between intravascular microthrombosis and neuronal death in human with fatal TBI. 48 Our present data demonstrate that early administration (2 h post-injury) of vepoloxamer exhibits neuroprotection by normalizing the bleeding time, and reducing brain hemorrhage, microthrombosis formation, the lesion size, and neuroinflammation.
A recent study indicates that P-188 may be protective in TBI by inhibiting megachannel opening (pannexin channels), 49 which may be related to its membrane sealing action. In addition, Miller and colleagues compared P-188 with kolloidon VA64 in a mouse CCI model and found few protective effects of P-188. 50 The difference between strong protective effects of purified P-188 vepoloxamer in the current study and less effects of P-188 by Miller and colleagues is not clear and may result from the different purity of the compounds (purified P-188 vepoloxamer vs. unpurified P-188), species (rat vs. mice), administration time post-injury (2 h vs. 5–10 min), dose (low dose of purified P-188 vs. high dose of P-188), and outcome measurements (brain tissue loss at 35 days vs. membrane sealing action at 2 h). The ability of vepoloxamer to reduce brain tissue loss and functional deficits supports the idea that other mechanisms in addition to membrane resealing properties also may contribute to its observed beneficial effects in rat after TBI.
In conclusion, we have demonstrated for the first time that intravenous administration of vepoloxamer 2 h after TBI improves functional recovery and reduces neuroinflammation in rats, suggesting that this novel agent has potential value for treatment of TBI. Further investigation of the optimal dose and therapeutic window of vepoloxamer treatment for TBI and the mechanisms underlying its therapeutic effects is warranted.
Footnotes
Acknowledgments
This work was supported by internal funding from HFH. The authors would like to thank Susan MacPhee-Gray for editorial assistance.
Author contributions to the study and manuscript preparation include the following: conception and design—Xiong, Chopp, ZG Zhang; acquisition of data—Y Zhang, Xiong; analysis and interpretation of data—Lu, T Zhang, Xiong, ZG Zhang, Chopp; drafting of the manuscript—Y Zhang, Xiong; critical review of the manuscript—Xiong, Chopp, ZG Zhang, L Zhang, Emanuele, Mahmood; review of final version of the manuscript and approval for submission—all authors.
Author Disclosure Statement
Martin Emanuele is an employee of LifeRaft Technologies, Inc., San Diego, CA. For the other authors, no competing financial interests exist.