
Abstract
Individuals living with chronic spinal cord injury (SCI) often exhibit impairments in cognitive function, which impede their rehabilitation and transition into the community. Although a number of clinical studies have demonstrated the impact of impaired cardiovascular control on cognitive impairment, the mechanistic understanding of this deleterious relationship is still lacking. The present study investigates whether chronic disruption of cardiovascular control following experimental SCI results in cerebrovascular decline and vascular cognitive impairment. Fourteen weeks following a high thoracic SCI (at the third thoracic segment), rats were subjected to a battery of in vivo and in vitro physiological assessments, cognitive-behavioral tests, and immunohistochemical approaches to investigate changes in cerebrovascular structure and function in the middle cerebral artery (MCA). We show that in the MCA of rats with SCI, there is a 55% (SCI vs. control: 13.4 ± 1.9% vs. 29.63 ± 2.8%, respectively) reduction in the maximal vasodilator response to carbachol, which is associated with reduced expression of endothelial marker cluster of differentiation 31 (CD31) and transient receptor potential cation channel 4 (TRPV 4) channels. Compared with controls, MCAs in rats with SCI were found to have 50% (SCI vs. control: 1.5 ± 0.2 vs. 1 ± 0.1 a.u., respectively) more collagen 1 in the media of vascular wall and 37% (SCI vs. control: 30.5 ± 2.9% vs. 42.0 ± 4.0%, respectively) less distensibility at physiological intraluminal pressure. Further, the cerebral blood flow (CBF) in the hippocampus was reduced by 32% in the SCI group (SCI vs. control: 44.3 ± 4.5 mL/100 g/min vs. 65.0 ± 7.2 mL/100 g/min, respectively) in association with impairment of short-term memory based on a novel object recognition test. There were no changes in the sympathetic innervation of the vasculature and passive structure in the SCI group. Chronic experimental SCI is associated with structural alterations and endothelial dysfunction in cerebral arteries that likely contribute to significantly reduced CBF and vascular cognitive impairment.
Introduction
Debilitating cardiovascular impairments are among the leading causes of morbidity and mortality in individuals with spinal cord injury (SCI). 1 Disruption of sympathetic cardiovascular control in individuals with upper thoracic or cervical SCI results in severe bidirectional fluctuations in systemic blood pressure (BP), which occur on a daily basis. 2 Chronic concomitant exposure to these extreme hypotensive and hypertensive events (called orthostatic hypotension and autonomic dysreflexia, respectively) following SCI can alter cerebrovascular structure and function, leading to long-term changes such as reductions in blood flow, reactive hyperremia, and cognitive decline. Not surprisingly, SCI is associated with a three- to four-fold higher risk of stroke and a significantly greater risk of cognitive impairment. 3,4 From our review of 70 studies, up to 60% of individuals with SCI exhibit impairments in one or more cognitive domains such as memory, attention, concentration, or executive function. 4 Although cognitive impairment following SCI can be attributed to a number of comorbidities, considerable evidence substantiates the relationship between cerebrovascular dysfunction and cognitive impairment. Further, the dysfunctions could be more pronounced following SCI as the cardiovascular disease progression is accelerated in this population owing to the amplification of various physical, physiological, and environmental factors. 5
Multiple studies from our group and others have presented a relationship between cerebrovascular dysfunction and cognitive decline in individuals with SCI. 6 –8 Intriguingly, cerebrovascular function is only partially restored in response to normalization of systemic BP, indicating that other factors, for example, vascular remodeling, fibrosis, and endothelial dysfunction, may contribute to the observed vascular impairment associated with SCI. We have previously used a rat model to demonstrate that high-thoracic SCI leads to significant fibrosis and increased stiffness of the cerebrovasculature at the sub-acute stage. 9 Understanding the progression of these early deleterious effects is of great clinical significance as this will potentially reveal novel therapeutic targets to prevent or mitigate chronic cerebrovascular dysfunction and cognitive decline following SCI.
The purpose of this study is to test if vascular-cognitive impairment occurs in a chronic experimental model of SCI with potential applications for the development of therapeutic interventions. Here, we used vascular physiological and immunohistochemical assessments to investigate the structural and functional correlates of cerebrovascular dysfunction after SCI. We also evaluated the impact of SCI on resting cerebral blood flow (CBF) and short-term memory in this model. We hypothesized that chronic high-thoracic SCI might produce cognitive deficits associated with diminished vascular reactivity, structural changes in the middle cerebral artery (MCA), and reduced CBF.
Methods
Experimental design
All experimental procedures were carried out in accordance with the Guide for the Care and Use of Experimental Animals established by the Canadian Council on Animal Care and were approved by the University of British Columbia Animal Care Committee (approval certificate number: A18-0183). Twenty-eight adult (250–300 g) male Wistar rats were assigned into either a naïve control (CON, n = 14) or T3-complete spinal cord transection group (SCI, n = 14). Fourteen weeks post-injury, all rats were assessed for short-term memory using a novel object recognition test (NORT, Fig. 1). Following the NORT, the groups were subdivided equally (n = 7 each). Each subgroup was assigned for either in vitro physiological assessments using pressure myography or for magnetic resonance imaging (MRI) followed by immunohistochemistry.
Experimental timeline. The rats were divided into two experimental groups (n = 14 each): CON (uninjured) and SCI (complete transection at T3 spinal segment). At 14 weeks post-SCI (and age-matched controls), all rats underwent NORT. Following NORT, the groups were subdivided equally and assigned to: 1) in vitro physiological assessments using pressure myography, and 2) MRI followed by immunohistochemistry. MRI, magnetic resonance imaging; NORT, novel object recognition test; SCI, spinal cord injury. Color image is available online.
T3 Transection surgery and animal care
Surgery and animal care were conducted as described previously. 9 Rats were pre-treated with enrofloxacin (Baytril; 10 mg/kg, s.c., Associated Veterinary Purchasing [AVP], Langley, British Columbia, Canada) for 3 days. The animals were fasted the night before surgery. On the day of surgery, anesthesia was induced with 5% isoflurane (AErrane; AVP) in 100% oxygen at a flow rate of 1 L/min and maintained at 2–3% during the surgery. Enrofloxacin (10 mg/kg, s.c.), buprenorphine (Temgesic; 0.02 mg/kg, s.c., University of McGill Animal Resources Centre), and lactated Ringer's solution (5 mL, s.c.) were administered pre-operatively.
A dorsal midline incision was made through the skin and superficial muscle layer, as well as a blunt dissection of deep muscles overlying T1-4 vertebrae. Deep muscles around T2-3 vertebrae were carefully dissected to expose the intervertebral gap. The dura at the gap was then opened, and the spinal cord was transected with microscissors. Complete transection was confirmed by two surgeons via visual separation of the rostral and caudal spinal cord stumps under the microscope. Absorption sponges and Gelfoam (Pharmacia & Upjohn Company, Pfizer, New York, USA) were used to achieve hemostasis. The muscle and skin of the surgical site were closed with 4–0 Vicryl and 4–0 Prolene sutures, respectively. Animals received warmed lactated Ringer's solution (5 mL, s.c.) and were allowed to recover from anesthesia in a temperature-controlled environment (Animal Intensive Care Unit, HotSpot for Birds, Los Angeles, CA, USA). Enrofloxacin (10 mg/kg, s.c., once per day) and buprenorphine (0.02 mg/kg, s.c., twice per day) were administered for 3 days post-surgery.
Animals with SCI were housed in specialized cages, with rubber matting to facilitate movement, low-reaching water bottles, and food scattered on the cage bottom to encourage foraging. Animals were supported with an enriched diet, including meal replacement shake (Ensure, Abbott Laboratories, Abbott Park, IL, USA), fruit, spinach, HydroGel (Clear H2O, Westbrook, ME, USA), peanuts, and kibble (LabDiet, Rodent Diet 5001; PMI International, St. Louis, MO, USA). The bladder was manually emptied three times daily until spontaneous bladder function returned (∼10 days after injury). Health status was monitored and recorded daily for the first 2 weeks after surgery and on alternative days thereafter. All experiments and analyses were performed with the investigators completely blinded of the group assignments.
NORT
Short-term memory was evaluated using the NORT, a widely implemented test to assess retention memory and interest in exploring novel objects. 10 One day prior to the testing session, animals were individually placed in an empty container for 1 h to encourage habituation. One hour before the test, animals were exposed to two identical objects for 10 min in the same container, and then one object was replaced with a novel object. 11 –14 During the test, animals were allowed to explore the open field freely for 5 min while being videotaped. Discrimination index (D-index) was calculated by dividing the difference between the exploration time devoted to novel (TN) and Object 1 (T1) divided by the total exploration time (DI = [TN – T1]/[TN + T1]*100). 11
CBF
CBF maps were acquired using arterial spin labeling (ASL) MRI as previously described. 15 Briefly, water molecules from inflowing blood were magnetically excited by a spin inversion. The overall magnetization of a downstream area of the brain was modified upon the arrival of these labeled molecules. The resultant signal describes the amount of arterial blood delivered to the imaging slice within the transit time of the experiment. On the day of imaging, rats were sedated with isoflurane and maintained on a ventilator (Harvard Apparatus Inspira ASV, USA) to maintain expired CO2 at 35 mm Hg. Body temperature was maintained at 37°C. Assessments were performed on a 7 Tesla MRI scanner (Bruker Biospec 70/30 USR, Bruker, Ettlingen, Germany) using a quadrature volume transmit coil and 4-channel phased-array receivers.
Multiple inversion recovery echo-planar imaging (EPI) scans were acquired to obtain calibration data (tissue T1, inversion efficiency, and proton density) for the ASL scan (recovery time = 10 sec, echo time = 11.47 msec, 22 inversion times between 30 and 8000 msec spaced geometrically, field of view = 40 × 40 mm, matrix = 64 × 64, slice thickness = 2 mm, 9 slices, single shot). Flow-sensitive alternating inversion recovery EPI with a pre-saturation preparation 16 was used to acquire the ASL data at five inversion times to allow correction for variable transit times using same geometry as the calibration scans. CBF maps were calculated by fitting the full perfusion model, including non-zero arterial transit time and bolus outflow. 17
Pressure myography
MCA function was assessed using pressure myography (Living Systems, St. Albans, VT, USA) as described previously. 9,18,19 A 5 × 3-mm section of brain tissue containing the MCA was removed and placed in Ca2+ free physiological salt solution (PSS) at 4°C, with 1% bovine serum albumin. A branch-free M2 segment of MCA was dissected. The M2 segment of MCA is the branch-free segment of the MCA and is a widely accepted segment for the study of pressure-diameter relationships because the other branched segments cannot hold the perfusion pressure. 20 –22 The vessels were mounted on two glass micropipettes in the pressure myograph, and the diameter was measured using a digital CCD camera. MCAs were equilibrated at 80 mm Hg for 45 min in PSS containing (in millimoles) 141.9 NaCl, 4.7 KCl, 1.12 KH2PO4, 1.7 MgSO4.7H2O, 2.8 CaCl2, 10 HEPES, 5 dextrose, and 0.5 EDTA (pH = 7.4) solution. The baseline of the inner diameter of the MCA was measured, and only vessels that developed >20% spontaneous myogenic tone were used for the experiment. Passive distensibility was assessed in Ca2+-free PSS containing 0.002 M EGTA as intraluminal pressure was increased from 20 to 180 mm Hg in 20 mm Hg increments.
Endothelial function was assessed by measuring the response to administration of carbachol in the bath as previously described. 23,24 The MCA was first bathed in PSS and pressurized at 80 mm Hg to generate a stable degree of spontaneous myogenic tone. The vasodilator response to increasing concentrations of carbachol (10−10 to 10−5 M) was then determined. 25 The vessel was pre-incubated with a transient receptor potential cation channel 4 (TRPV 4) antagonist, HC067047 (10−6 M), for 15 min and the responses to carbachol (10−10 to 10−5 M) were recorded to investigate the mechanism involved. 26 Finally, the vessels were bathed in Ca2+-free PSS, and a passive pressure diameter curve was recorded. Myogenic tone was calculated using the following formula: % tone = (1−[active lumen diameter/passive lumen diameter]) × 100.
Perfusion and tissue collection
Animals were euthanized with isoflurane followed by intraperitoneal injection of chloral hydrate. The cerebral vasculature was fixed by transcardial perfusion with 100–150 mL of phosphate-buffered saline (PBS; at 80–90 mm Hg) and 400 mL of 4% paraformaldehyde at 120–140 mm Hg. The brain was harvested and placed in 4% paraformaldehyde for 24 h at 4°C and then transferred to 20% sucrose for 3 days. A 2- to 3-mm long segment of MCA was gently dissected from each hemisphere and used for histological analyses.
Immunohistochemistry
The M2 segments of MCA with a length of approximately 5 mm and nearly uniform cylindrical shape were isolated from the animals, and were embedded in Cryomatrix®. We systematically randomly sampled a series of sections for stereology as previously described. 21 Using a cryostat, 10-μm frozen transverse sections were cut and were placed on Superfrost Plus slides. The sections were then randomly selected, and stored at −80°C until processed for immunohistochemistry.
Briefly, MCA sections were washed in PBS and 0.1 M PBS containing 0.3% Triton X-100 (PBST). They were blocked for 1 h in 10% normal donkey serum in PBST. Sections were then incubated with primary antibodies to: collagen 1 (COL 1, 1:500, Abcam, ab34710), collagen 3 (COL 3, 1:300, Abcam, ab6310), TRPV 4 (1:300, generous gift from Dr. Stefan Heller's lab at Stanford), cluster of differentiation 31 (CD31, 10 μg/mL, R&D systems, AF3628), or smooth muscle actin (SMA, 1:400, Abcam, ab5694) overnight at room temperature. On the second day, the sections were washed in PBST and incubated with species-specific secondary antibodies: Alexa™-fluor 488 donkey anti-mouse (1:1000, Molecular Probes©, USA), Alexa™-fluor 488 donkey anti-goat (1:1000, Molecular Probes©), or Cy3 conjugated donkey anti-rabbit IgG antibody (1:1000, Jackson ImmunoResearch) for 1 h. Finally, the sections were washed with PBST, coverslipped with Vectashield mounting medium (Vector Laboratories, Inc.), and images were captured using a confocal microscope with identical acquisition parameters.
Quantification
The intensity of immunoreactivity of MCAs was assessed using Fiji Software (
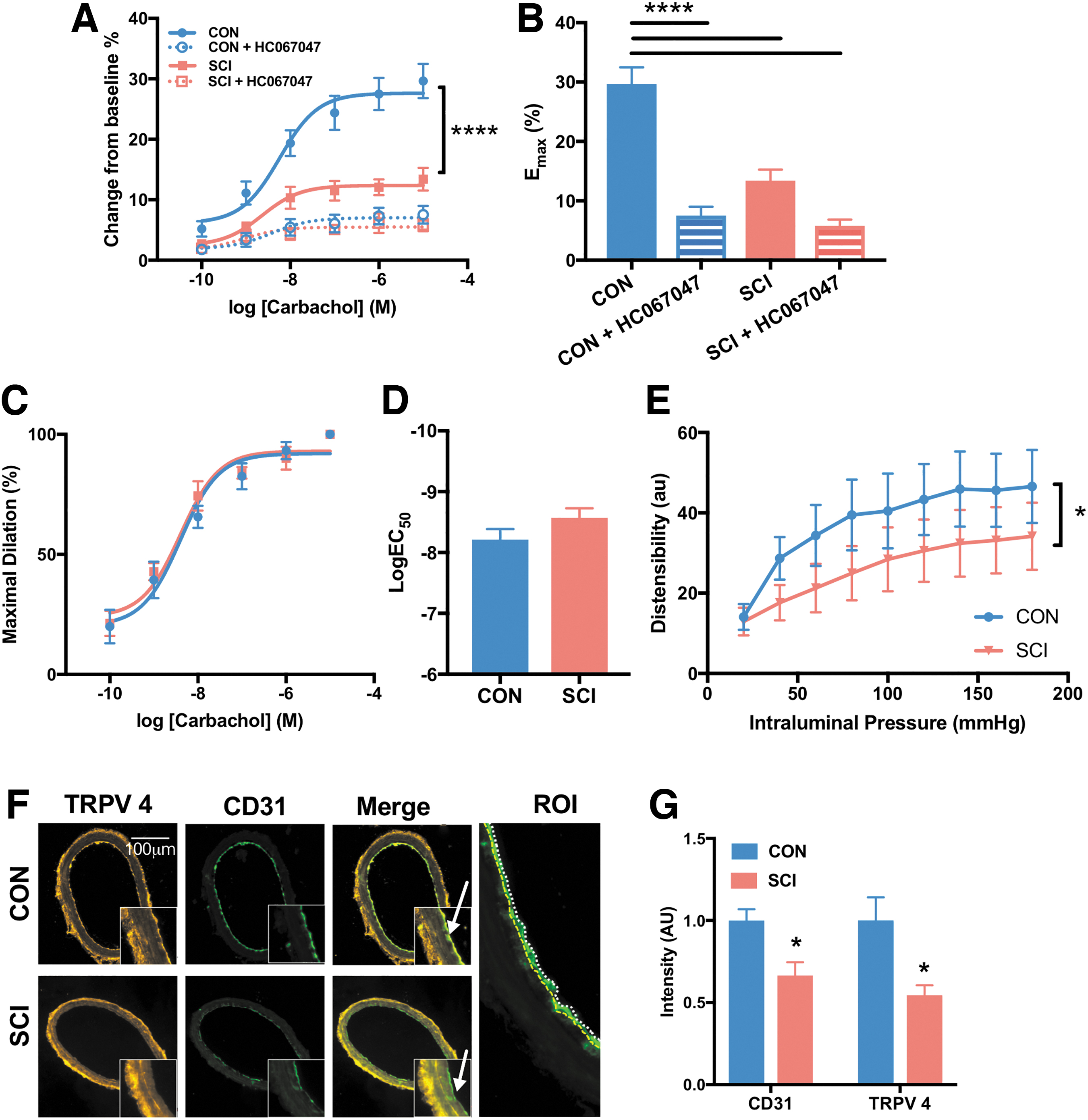
Carbachol-induced relaxation is diminished in MCA after chronic SCI, which is closely associated with the loss of TRPV 4 and CD31 staining in the endothelium.
Morphometric assessment of the MCA
The arterial wall thickness and lumen diameter were measured using the transverse sections of the MCA immunostained with SMA (four sections per rat). Within each section, four sites were selected, representing a quadrant of the vessel, and a line was drawn across the media of the vascular wall perpendicular to the tangent line, and the length of this line was measured. The average of the four measurements was used to denote wall thickness for each section. Lumen diameter was measured by taking the average of the widest and the narrowest part of the lumen.
Tyrosine hydroxylase (TH) staining of the MCA whole mounts
A small segment (3–4 μm) from the middle third of the MCA was stained for tyrosine hydroxylase (TH, 1:1000, anti-rabbit, Millipore, AB152) to compare the degree of sympathetic innervation. The density of TH-positive (TH+) nerve fibers was measured using Fiji Software (using ImageJ,
Statistical analysis
Mean values ± standard error are presented. The significance of differences between mean values in the control and spinal cord transected groups was determined using a Student's t test or a non-parametric alternative when the data were not normally distributed. Repeated measures analysis of variance (ANOVA) was used for the concentration-response curves for carbachol, which were further analyzed by non-linear regression (Hill equation) to calculate EC50 and Emax values. All the statistical analyses were carried out using GraphPad Prism 7.0 software (GraphPad, San Diego, CA). A p-value of less than 0.05 was considered to be statistically significant.
Results
Altered endothelial function and mechanical properties of the MCA after chronic SCI
The endothelial function of the MCA was impaired in the rats with SCI (Fig. 3). The maximal vasodilation response to carbachol was 55% (SCI vs. CON: 13.4 ± 1.9% vs. 29.63 ± 2.8%, respectively) less in the SCI group than in the uninjured controls (Fig. 3A,B), but the EC50 was not significantly different (Fig. 3C,D). Further, when tested across a range of intraluminal pressures in Ca2+free solution, the distensibility of the MCA was significantly reduced, by about 37% (SCI vs. CON: 30.5 ± 2.9% vs. 42.0 ± 4.0%, respectively) over a range of pressures from 80 to 180 mm Hg (Fig. 3E).
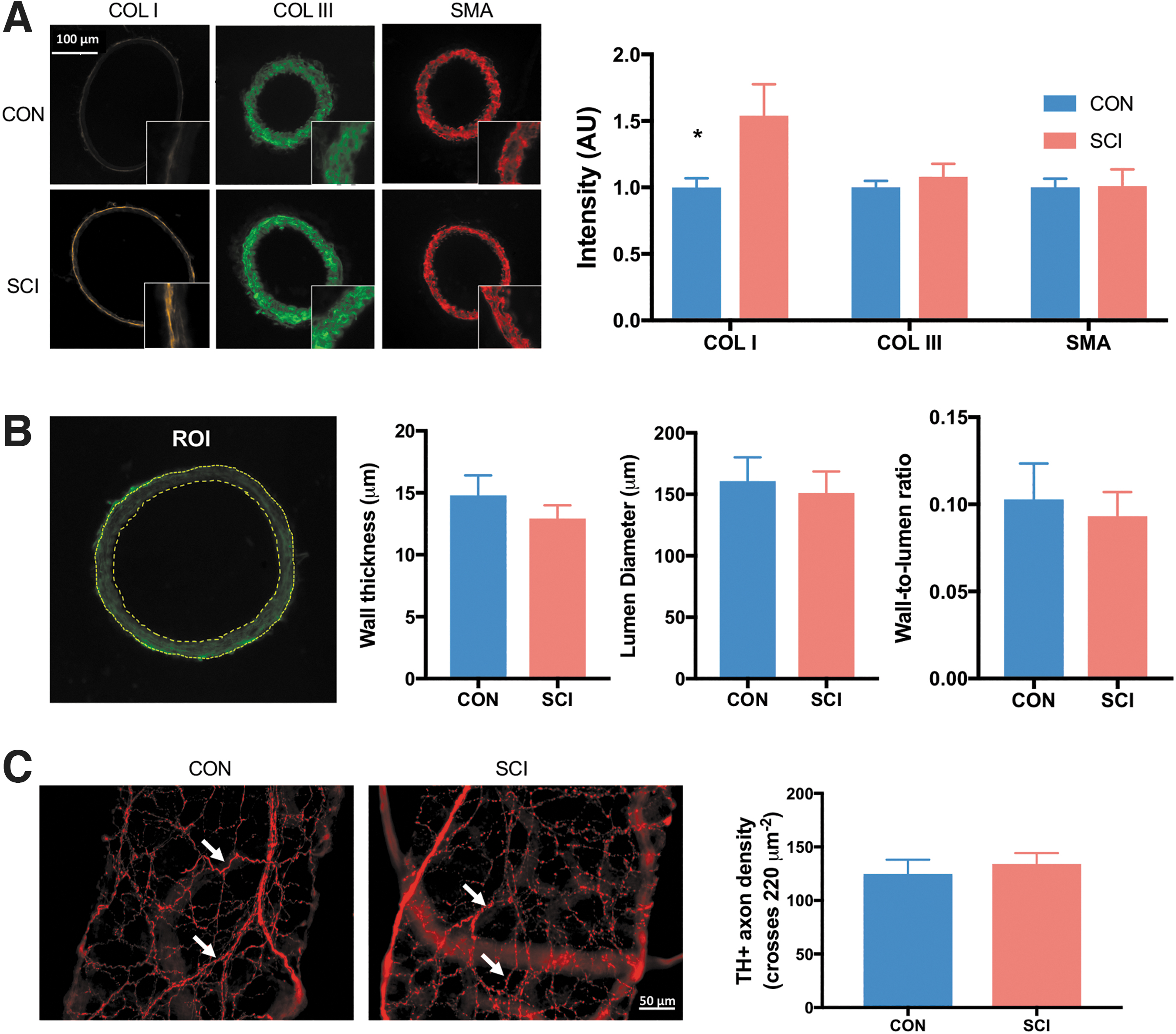
Chronic SCI leads to increased collagen deposition in the arterial wall.
The difference in endothelium-mediated vasodilation response between the two groups was likely mediated via endothelial TRPV 4, as the response to carbachol was abolished in both groups following treatment with the TRPV 4 channel antagonist HC067047 (Fig. 3A,B). This was further supported by immunohistochemical analysis, which demonstrated that endothelial TRPV 4 expression was 46% (SCI vs. CON: 0.5 ± 0.1 a.u. vs. 1 ± 0.1 a.u., respectively) lower in rats with SCI (Fig. 3G). Further, a 34% (SCI vs. CON: 0.7 ± 0.1 a.u. vs. 1 ± 0.1 a.u., respectively) lower expression of endothelial CD31 in the SCI is suggestive of loss of endothelial cells or changes in their function (Fig. 3G).
Morphological and structural changes in the MCA after SCI
Immunohistochemical analyses revealed there is a marked 50% (SCI vs. CON: 1.5 ± 0.2 a.u. vs. 1 ± 0.1 a.u., respectively) increase in COL 1 deposition in the media of the MCA after SCI (Fig. 4A). This is associated with a significant reduction in SMA staining. However, there was no significant difference in wall thickness, lumen diameter, or wall to lumen ratio (Fig. 4B). Perivascular sympathetic innervation of MCAs, as revealed by TH+ axonal density, was also not affected by SCI (Fig. 4C).
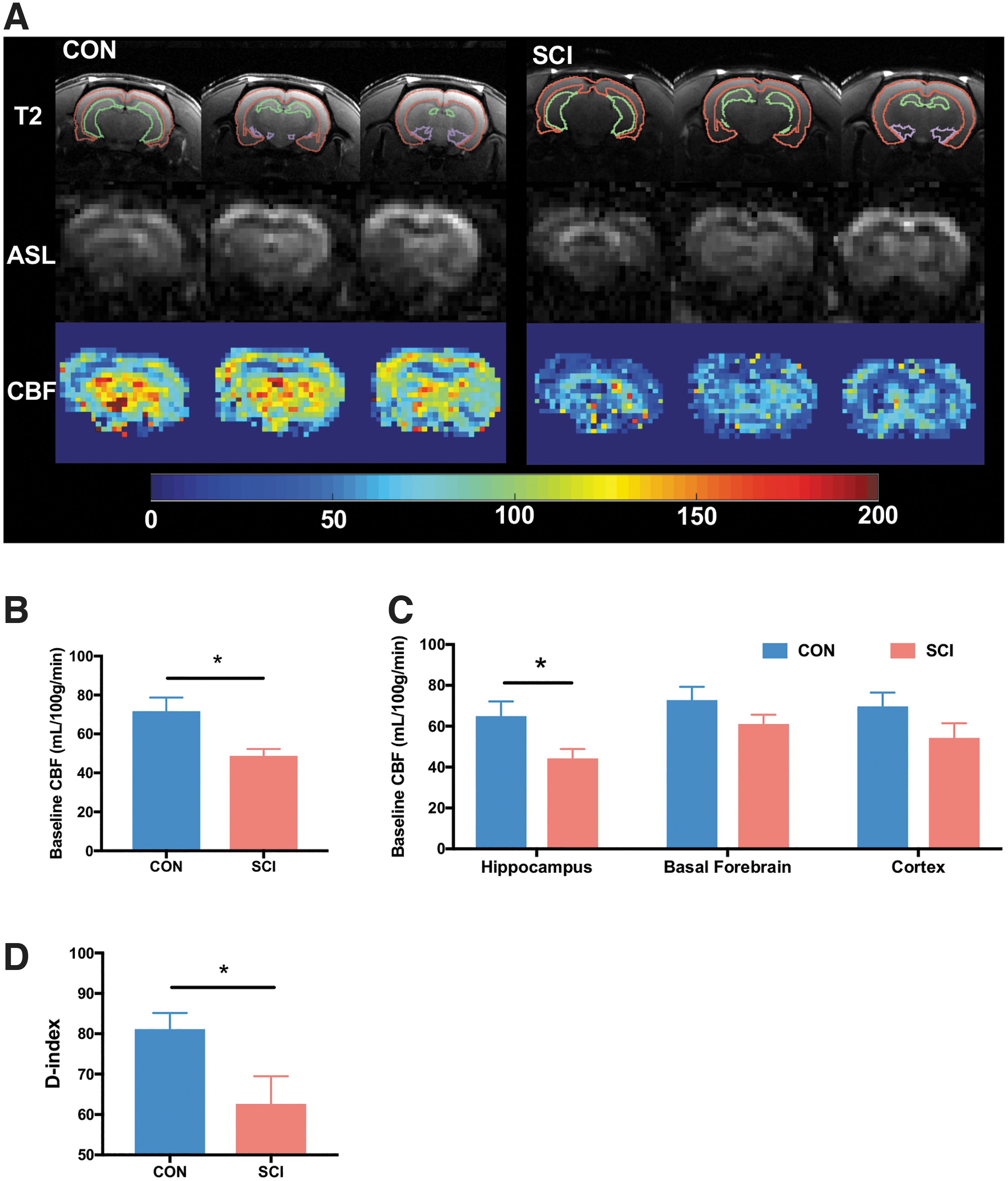
Reduced baseline CBF and impaired spatial short-term memory are observed after prolonged high-thoracic SCI.
Chronic SCI is associated with a reduction in CBF
Resting CBF was 32% (SCI vs. CON: 48.7 ± 3.5 mL/100 g/min vs. 71.7 ± 7.0 mL/100 g/min, respectively) lower in T3-SCI (Fig. 2A,B), indicating reduced brain perfusion following chronic SCI. Moreover, regional flow in SCI was significantly reduced in the hippocampus (32%, SCI vs. CON: 44.3 ± 4.5 vs. 65.0 ± 7.2 mL/100 g/min, respectively, Fig. 4C). No effects were seen on CBF in the basal forebrain or cortex.
SCI disrupts short-term memory
The control rats showed a higher preference for the novel object (D index = 81.1 ± 13.9%). In contrast, the rats with chronic SCI had significantly reduced preference for the novel object (D index = 62.6 ± 19.2%), suggesting impairment in short-term retention memory (Fig. 2D).
Discussion
This study provides novel mechanistic evidence of vascular cognitive impairment in a model of SCI that consistently results in highly labile systemic BP. Although a number of clinical studies in recent years have demonstrated associations between disrupted cardiovascular control and cognitive impairment following SCI, the majority of them are unable to rule out the contribution of a concomitant traumatic brain injury (reviewed in a previous article by Sachdeva and collegues 4 ). The present results indicate that chronic SCI results in structural and functional impairments in the MCA, namely: endothelial dysfunction, elevated expression of COL 1, the major collagen isoform in the media of vascular wall, and significantly impaired MCA distensibility. Consistent with the clinical literature, 6,29 we found that experimental high-thoracic SCI results in significant cerebral hypoperfusion. Our comprehensive review has concluded that regulation of CBF following SCI is drastically impacted via a number of factors, including endothelial dysfunction. 30 Interestingly, the reduction in CBF in the present study was observable in the hippocampus, a region associated with memory, and likely more susceptible to ischemic damage due to its circulatory pattern. 31 As hypothesized, a high-thoracic SCI that is known to cause detrimental cardiovascular and cerebrovascular impairments resulted in significant impairments in memory. Retention memory was assessed using the NORT, which is relatively less dependent on locomotion compared with other tests of cognitive function that have been employed previously in rats with SCI. 28
Endothelial dysfunction was identified as a prominent outcome in the chronic SCI, as indicated by the impaired response to carbachol. This is an important observation in light of our previous findings that endothelial function remains unaltered in the sub-acute stage of SCI. 9 This finding may indicate a therapeutic time window for early intervention that may be capable of preventing or impeding the ensuing decline in endothelial function, CBF, and cognitive ability—evident characteristics of chronic SCI. Endothelium-dependent relaxation, as induced by carbachol is primarily mediated via three pathways: nitric oxide (NO), prostacyclin and/or endothelial-derived hyperpolarization factor (EDHF). 32 Particularly in the MCA, carbachol activates the M3 muscarinic receptor in endothelial cells that opens TRPV 4 channels, resulting in calcium influx, which increases the formation of NO and EDHF. 33 Therefore, blockade of TRPV 4 channels suppresses NO- and EDHF-mediated vasodilation. 34 –36
Interestingly, the administration of a TRPV 4 channel inhibitor had a greater effect to reduce the vasodilator response to carbachol in control animals than SCI animals, whereas the residual response to carbachol remained similar in both groups. This is consistent with endothelial dysfunction and reduced production of NO in the SCI rats. Therefore, the underlying mechanism behind SCI-derived loss of endothelial reactivity may be related to a reduction in TRPV 4 channel activity. This argument is further supported by our immunohistochemical data, where the expression of endothelial TRPV 4 is reduced following SCI. Because TRPV 4 channels are sensitive to shear-stress, the abnormal expression of endothelial TRPV 4 receptors after T3-SCI may be associated with the global reduction in CBF. 33 Further, decreased expression of CD31 suggests potential structural damage to the MCA endothelium following SCI, consistent with other reports of reduced CD31 in models of cognitive impairment associated with ischemia. 37
In addition to endothelial dysfunction, SCI resulted in a marked increase in COL 1 deposition in the MCA and reduced distensibility, suggesting stiffer arteries. The reduced distensibility of theMCA after 14-week SCI is consistent with our previous study. 38 The differences of MCA wall thickness and inner diameter between the present and previous study are likely due to the assessment of fixed tissue that causes further constriction and affects vascular structure. The differences in vascular structure in the present study should be interpreted based on the pressure-diameter measurements in the Ca2+ free solution in the pressure myograph system. Although previous literature implies a role of disrupted sympathetic innervation in increasing fibrosis in the aorta, 39 it is unlikely that a similar mechanism is responsible for the stiffening of the MCA in the present study. This is supported by the near-identical TH staining pattern in the MCAs of the two experimental groups. However, this preliminary observation should be further investigated in dedicated structural studies using higher magnification and co-localization analyses. Moreover, considering the chronic increased BP variability following disruption of sympathetic control, the role of neuro-humoral processes in mediating arterial stiffening via increased pro-inflammatory factors should be investigated as a possible underlying mechanism. 40
Future perspectives
We present an experimental high-thoracic SCI model that leads to endothelial dysfunction, structural remodeling of the cerebral arteries, global reduction in CBF, and vascular-cognitive impairment, which has now been identified as a serious disease burden in individuals with SCI. 5,7,15,41 Interestingly, there appears to be a therapeutic time window between the establishment of vascular fibrosis and the onset of cognitive symptoms, offering various directions for upcoming research. Future studies could explore a number of intriguing avenues, such as spatiotemporal changes in expression of specific receptor subtypes to facilitate testing of potential drugs and other interventions for early prevention of vascular-cognitive impairment following SCI. For instance, upregulation of TRPV 4 channel activity may be a viable therapy for mitigating loss of cerebrovascular function. JNc-440, a designed molecule to strengthen the interaction between TRPV 4 and activated potassium channel 3 (KCa2.3), has been recently reported to enhance endothelial-induced vasodilation and exert anti-hypertensive effects. 42 A possible clinically relevant intervention could also be physical exercise, which has been shown to have significant benefits on endothelial function. 43,44 Moreover, a better understanding of SCI-induced structural and functional changes in cerebral microvessels is needed. Alterations in the endothelial health of microvessels will likely provide deeper insight into the underlying mechanisms of vascular-cognitive impairment after chronic SCI and provide novel therapeutic targets. 45
Footnotes
Funding Information
The present study was supported by funds from the Heart and Stroke Foundation (G-16-00012571) and Canadian Institutes of Health Research (389694). Dr. Sachdeva is supported by Postdoctoral Fellowships from the Craig H. Neilsen Foundation (455064), Canadian Institutes of Health Research (415366), Michael Smith Foundation for Health Research (18408) and University of British Columbia (Bluma Tischler Postdoctoral Fellowship). The Phillips Lab is supported by the Wings for Life Foundation (Project Grant 202), Compute Canada (Resources for Research Groups nnu-172), Natural Sciences and Engineering Research Council (Canada; Discovery Grant RGPIN/03771-2018), the Canadian Institutes of Health Research (Project Grant 159573), Alberta Innovates Health Solutions, Campus Alberta Neuroscience, the Libin Cardiovascular Institute of Alberta, the Hotchkiss Brain Institute, and the Rick Hansen Institute. Dr. Fan and Roman Labs were supported by grants AG050049 (F.F.), AG057842 (F.F.) and HL138685 (R.J.R.) from the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.