
Abstract
One of the difficulties in identifying novel therapeutic strategies to manage central nervous system (CNS) trauma is the need for behavioral assays to assess chronic functional recovery. In vitro assays and/or acute behavioral assessments cannot accurately predict long-term functional outcome. Using data from 13 independent T9 moderate contusive spinal cord injury (SCI) studies, we asked whether the ratio of acute (24–72 h post-injury) changes in the levels of neuron-, oligodendrocyte-, astrocyte-specific and/or endoplasmic reticulum stress response (ERSR) messenger ribonucleic acids (mRNAs) could predict the extent of chronic functional recovery. Increased levels of neuron, oligodendrocyte, and astrocyte mRNAs all correlated with enhanced Basso Mouse Scale (BMS) scores. Reduced levels of the ERSR mRNAs Atf4 and Chop correlate with improved chronic locomotor function. Neither neural or ERSR mRNAs were predictive for chronic recovery across all behavioral changes. The ratio of oligodendrocyte/ERSR mRNAs, however, did predict “improved,” “no change,” or “worse” functional recovery. Neuronal/ERSR mRNA ratios predicted functional improvement, but could not distinguish between worse or no change outcomes. Astrocyte/ERSR mRNA ratios were not predictive. This approach can be used to confirm biological action of injected drugs in vivo and to optimize dose and therapeutic window. It may prove useful in cervical and lumbar SCI and in other traumatic CNS injuries such as traumatic brain injury and stroke, where prevention of neuronal loss is paramount to functional recovery. Although the current analysis was directed toward ERSR whose activity was targeted in all but one study, acute mRNA markers for other pathophysiological cascades may be as predictive of chronic recovery when those cascades are targeted for neuroprotection.
Introduction
One of the difficulties in identifying therapeutic strategies for the management of central nervous system (CNS) trauma is the need for behavioral assays to unequivocally assess the extent of chronic functional recovery. In vitro assays and/or acute behavioral assessments cannot accurately forecast long-term outcome. In spinal cord injury (SCI) studies, the commonly used open field locomotor assessments in rat and mouse, Basso, Beattie, Bresnahan (BBB) 1 and Basso Mouse Scale (BMS), 2 respectively, can require up to eight weeks of analyses. If one needs to establish optimal dose and/or therapeutic windows, the numbers of animals and the time required becomes unwieldy, especially in a matrixed experimental design. We asked whether an acute in vivo assay could be used to predict chronic therapeutic efficacy of a given intervention.
The pathophysiology of SCI involves the primary injury and a secondary injury cascade that progresses hours to months post-SCI. 3,4 Neural (neurons, astrocytes, and oligodendrocytes) cell loss occurs both acutely in the injury epicenter as well as progressively in the injury penumbra. 3 –7 Depending on the level (cervical, thoracic, lumbar), type (contusive, compressive, hemi-section, total transection), and severity of SCI, the contribution to functional recovery of the remaining specific neural cell populations is of more or less importance. For example, in cervical and lumbar contusive injuries, neuronal loss contributes significantly to the functional deficits, while in thoracic contusive SCI, white matter sparing (axons and oligodendrocytes) is the major determinant of the extent of functional recovery. 8 –13
While hemi-section and transection injuries result in the loss of axons of passage, contusive and compressive SCIs result in the loss of all neural cell types in the immediate area of injury. We hypothesized that functional recovery after these latter injury types could be predicted by the extent of loss of the specific neural cell type(s) important for that lesion model utilized.
The proteostasis (protein homeostasis) network includes all proteins with an immediate role in protein synthesis, folding, disaggregation, or degradation. 14 This network includes all proteins necessary for translation, chaperone proteins needed for proper folding, as well as the ubiquitin proteasome system (UPS) and autophagy systems that degrade proteins. Integral to proteostasis are the stress response pathways, the heat shock response (HSR), the endoplasmic reticulum stress response (ERSR), the integrated stress response (ISR), and the unfolded protein response (UPR), whose functions are to restore homeostasis.
The ERSR/UPR/ISR involve overlapping signaling modules. The ERSR consists of three signaling pathways that are activated in response to enhanced levels of misfolded proteins in the ER lumen: (1) ERSR-activated protein kinase ribonucleic acid (RNA) (PKR)-like kinase (PERK), (2) activating transcription factor 6 (ATF6), and (3) inositol-requiring protein-1α (IRE-1α), which signals through X-box binding protein 1 (XBP1). 14,15
ATF6 and IRE1/XBP1 make up the pro-homeostatic UPR signaling pathway. PERK signaling, as well as that of the three ISR kinases (general control non-derepressible [2-GCN2], protein kinase R-PKR, and heme-regulated inhibitor [HRI]) all phosphorylate eukaryotic initiation factor 2α (eIF2α), the main driver of eukaryotic translation, to alleviate cellular stress by inhibiting general protein translation. There is a concomitant increase in ERSR-specific translation to increase ER luminal protein folding capacity and enhance degradation of misfolded proteins. Growth Arrest and DNA Damage-Inducible protein 34 (GADD34), a downstream ERSR effector, dephosphorylates peIF2α to restore homeostasis.
All CNS injuries initiate some aspects of the proteostasis response in all affected cells, and the greater the insult, the stronger the response. If cellular homeostasis cannot be restored, the ERSR/ISR/UPR initiates apoptosis in those cells, driven predominantly by activating transcription factor 4 (ATF4) and CHOP (C/EBP (CCAAT enhancer binding protein) homologous protein). 16 –18
Our 19 –24 and others' 25 –30 works, as well as our unpublished data, have demonstrated a functional role of the ERSR and other aspects of the proteostasis network in the acute responses to SCI and, importantly, chronic locomotor recovery. Here, using both our published and unpublished data from 13 independent studies, we ask whether acute (24–72 h post-injury) changes in the levels of neural- and ERSR-specific messenger RNAs (mRNAs) could predict the extent of functional recovery (6-week post-injury BMS scores) after SCI.
Current data do have inherent bias because all but one of these studies directly modulate individual aspects of the ERSR, which is also used as one output measure. We define a novel analytical approach, however, that may be used with the ERSR and other pathophysiological signaling pathways to simplify optimizing variables of efficacy, dose, bioavailability, and therapeutic window that would otherwise necessitate extensive chronic behavioral studies.
Methods
SCI
As reported previously, 50 kdyn IH (Infinite Horizons, Lexington, KY) T9 contusive injuries were performed. 19,22,31 In all studies, wild type (WT), genetic variant, vehicle, or drug-treated animals were assigned blindly and randomly for surgical order. Injury displacement values for all animals used in all studies ranged 400–600 μm.
Data analyzed
For the quantitative polymerase chain reaction (qPCR) studies, mRNA levels were determined in a 5-mm segment of the contused spinal cord tissue spanning the injury epicenter and the penumbra. This was consistent for all 13 studies. The qPCR and BMS data (Table 1) from the following published studies were used: Chop-/- 19 , salubrinal (Sal, an inhibitor of peIF2α dephosphorylation), 22 guanabenz (Gnb, an inhibitor of peIF2α dephosphorylation) and Gadd34-/-, 21 Atf6-/- , 24 Plp-creERT2:Xbp1fl/fl , 63 and phosphodiesterase 4B null mice (Pde4b-/- ), 32 as well as unpublished data from five others: GSK2656157 (GSK, a PERK phosphorylation inhibitor), Hri-/-, Pkr-/-, Gcn2-/-, Plp-creERT2:Perkfl/fl (OL-Perk-/-), (OL-Xbp1-/-), and adaptaquin (AQ, an inhibitor upstream of ATF433). The Plp-creERT2 (proteolipid protein) promoter cre construct will delete the floxed gene specifically in all cells of the later oligodendrocyte lineage after treatment with tamoxifen. Details can be found in the cited articles.
Table 1. Spinal Cord Injuries Studies Evaluated
, SCI, spinal cord injury; ERSR, endoplasmic reticulum stress response; mRNA, messenger ribonucleic acid; BMS, Basso Mouse Scale.
E/C, Experimental/Control; 2N, neurons; OL, oligodendrocytes; A, astrocytes.
p < 0.05 BMS week 6 experimental vs. control.
qPCR markers
The qPCR was performed on an ABI 7900HT real-time PCR instrument (Applied Biosystems, Foster City, CA). Briefly, diluted complementary DNAs (cDNAs) were added to Taqman universal PCR master mix (Applied Biosystems) and run in triplicate. Target and reference gene PCR amplification was performed in separate tubes with the following Assay on Demand™ primers (Applied Biosystems). Neural qPCR markers included claudin 11 (Cldn11, Mm00500915_m1), Olig2 (Mm01210556_m1), and myelin basic protein (Mbp, Mm01266402_m1) (oligodendrocytes), neuron-specific enolase (Nse/Eno2, Mm00469062_m1) and microtubule-associated protein 2 (Map2, Mm00485230_m1) (neurons), and glial fibrillary acidic protein (Gfap, Mm01253033_m1) and glutamine synthetase (Glul, Mm00725701_s1) (astrocytes).
The ERSR mRNAs quantified include: Atf4 (Mm00515324_m1, downstream of PERK), Chop/Ddit3 (Mm01135937_g1), Xbp1 (Mm00457359_m1), Gadd34/Ppp1r15a (Mm00435119_m1), and 78 kDa glucose regulated protein (Grp78/Hspa5, Mm01333323_g1, an ER luminal chaperone protein that binds to PERK, ATF6, and IRE1 in the absence of unfolded proteins).
The qPCR assessments of multiple neural and ERSR mRNAs were compiled from the 13 studies performed acutely at 24, 48, or 72 h post-SCI. Specifically, mRNAs were isolated at 24 (Atf6-/- ), 48 (GSK, OL-Perk-/- ), and 72 (Chop-/- , Salubrinal, Hri-/- , Gnb, Gcn2-/-, Pkr-/-, Gadd34-/- , AQ, and OL-Xbp1-/- ) hours post-SCI. Initial experiments showed significant upregulation across all of these acute time points. 19
BMS
Locomotor ability was assessed at weekly intervals post-SCI using the BMS locomotor rating scale2 as reported previously. 19,22 Mice were assessed in an open-field setting and rated by two trained observers, both blinded to experimental design and treatment. Mean left and right hindlimb scores from six weeks were used. Based on the BMS outcome at week 6, the 13 studies were grouped into three categories reflecting functional outcome. The first included studies with significant “improved” differences in the BMS scores between the experimental and control groups (five studies), the second encompassed those with non-significant differences between the groups at week 6 (“no change,” six studies), and the third included studies with significant poorer outcomes in which BMS averages of controls were higher than those of the experimental group at week 6 (“worse,” two studies).
Correlation analyses
The means of the week 6 BMS scores, each of the neural and ERSR mRNAs in the experimental group were divided by the mean of the control group (E/C). Correlations between E/C Atf4 and Chop with their corresponding E/C BMS were calculated in the following manner: For the Atf4 E/C correlation with BMS E/C in the Improved Chop study, (n = 4 control, n = 4 experimental), the average for the four control = 3.30 and the average for the four experimental = 1.26. The Improved Chop study E/C of Atf4 = 0.383 (3.30/1.26). An Atf4 E/C was calculated for each of the 13 studies. Each of the 13 Atf4 E/C values were correlated with their respective E/C BMS value.
Individual markers were pooled when averaged into neural, oligodendrocyte, astrocyte, and ERSRs for each of the 13 studies. For example, in the salubrinal study, the neuronal markers were calculated from the mean of the experimental group (n = 4) for Map2 divided by the mean of the control group (n = 3) mean Map2 = E/C ratio of Map2. The same calculation was repeated for the Nse/Eno2 mRNAs in the study. The mean of the Map2 and Nse/Eno2 = Neural E/C. These values were calculated for each of the 13 studies. For oligodendrocytes, this mean was calculated from Cldn11, Olig2, and Mbp E/C mRNA ratios, and for astrocytes Glul and Gfap for each of the 13 studies.
Statistical analysis
Averaged E/C neuron (Map2, Nse), oligodendrocyte (Olig2, Claudin11, Mbp), and astrocyte (Gfap, GS) mRNA ratios then were divided by E/C ERSR mRNAs Atf4 and Chop only to obtain the ratio of neural to ERSR ratios. Ratios were compared among the three BMS outcome study categories using one-way analysis of variance, followed by Tukey HSD post hoc t tests for multiple comparisons. Median tests were performed comparing neural markers (E/C) among the outcome groups. The relationship between the neural markers with the ERSR and BMS E/C ratios was performed using parametric or nonparametric (Pearson [r], Spearman rank [rS], respectively) correlations, depending on sample sizes. An astrocyte outlier (>3 standard deviations from the mean) 19 was removed from correlations, because outliers will inflate coefficients artificially and reduce p values.
Results
We initially examined the correlation between the E/C ratio of individual acute ERSR mRNA levels and the ratio of E/C chronic BMS scores among our completed 13 SCI studies (Fig. 1). For all genes evaluated, those with improved (•) or worse (▴) BMS scores tended toward lower or higher ERSR ratios, respectively, while those with no change (□) were intermediate. Individually, regression analysis showed Atf4 (Fig. 1d) and Chop (Fig. 1e) to be significant while Grp78 (Fig. 1a), Gadd34 (Fig. 1b), and Xbp1 (Fig. 1c), were not.
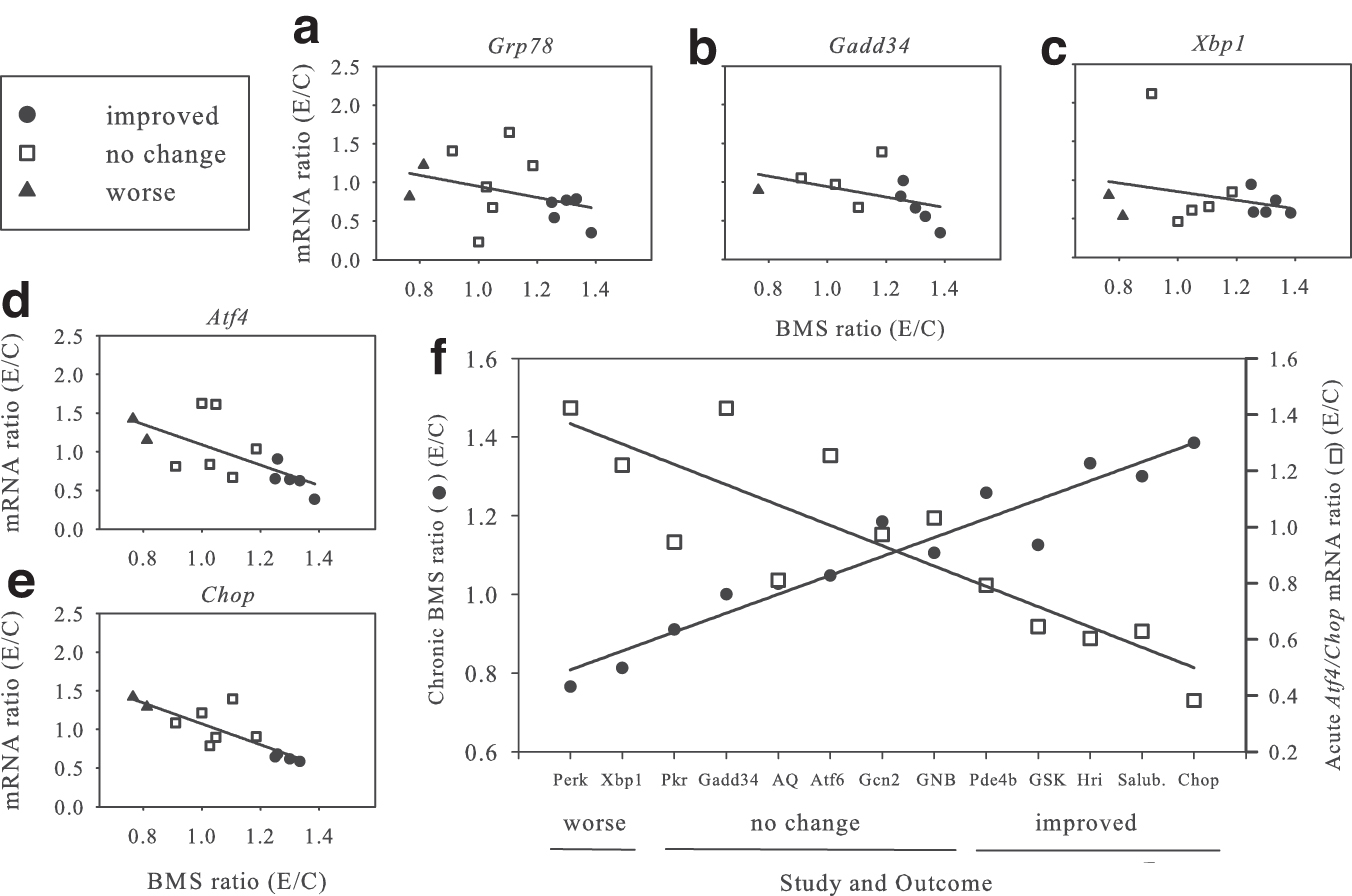
Correlation of acute endoplasmic reticulum stress response (ERSR) messenger ribonucleic acid (mRNA) levels with chronic six-week Basso Mouse Scale (BMS) ratios. (
When all 13 studies were aligned based on increasing BMS score (Fig. 1f) (•, left y axis), linear regression analysis showed a highly significant correlation. Similarly, if the respective ratios of the significant E/C ERSR mRNAs (Atf4, Chop) values (□, right y axis) for each of those studies were plotted, linear regression analysis showed a significant negative relationship. The inverse relationship between E/C BMS score and Atf4/Chop mRNA levels was highly significant (r 2 = 0.66, p < 0.001).
We queried whether assessing acute E/C ERSR mRNA levels could be used to predict chronic locomotor recovery. Pooling of these experimental to control averaged mRNA ratios by outcome (improved, no change, worse) showed that the ERSR mRNAs were significantly higher in the worse (p < 0.05) and no change (p < 0.01) groups compared with improved but were not different from each other (data not shown), making their predictive value inaccurate across all behavioral recovery levels.
We undertook a similar analysis for each of the E/C ratios of neuronal (Map2, Nse/Eno2) (Fig. 2a,b), oligodendrocyte (Olig2, Mbp, Cldn11) (Fig. 2c–e), and astrocyte (Gfap, Glul) (Fig. 2f,g) mRNAs to the ratio of E/C BMS scores. Note that not all studies tested every single mRNA. Data are presented for improved studies (•), those with no behavioral change (□), and those with worse behavioral recovery (▴). For all mRNAs evaluated, except Cldn11 where p = 0.06, regression analyses showed significant (p < 0.05) correlations. Averaging the values for the E/C mRNA ratios for neurons (Fig. 2h), oligodendrocytes (Fig. 2i), or astrocytes (Fig. 2j) showed similar significant correlations.
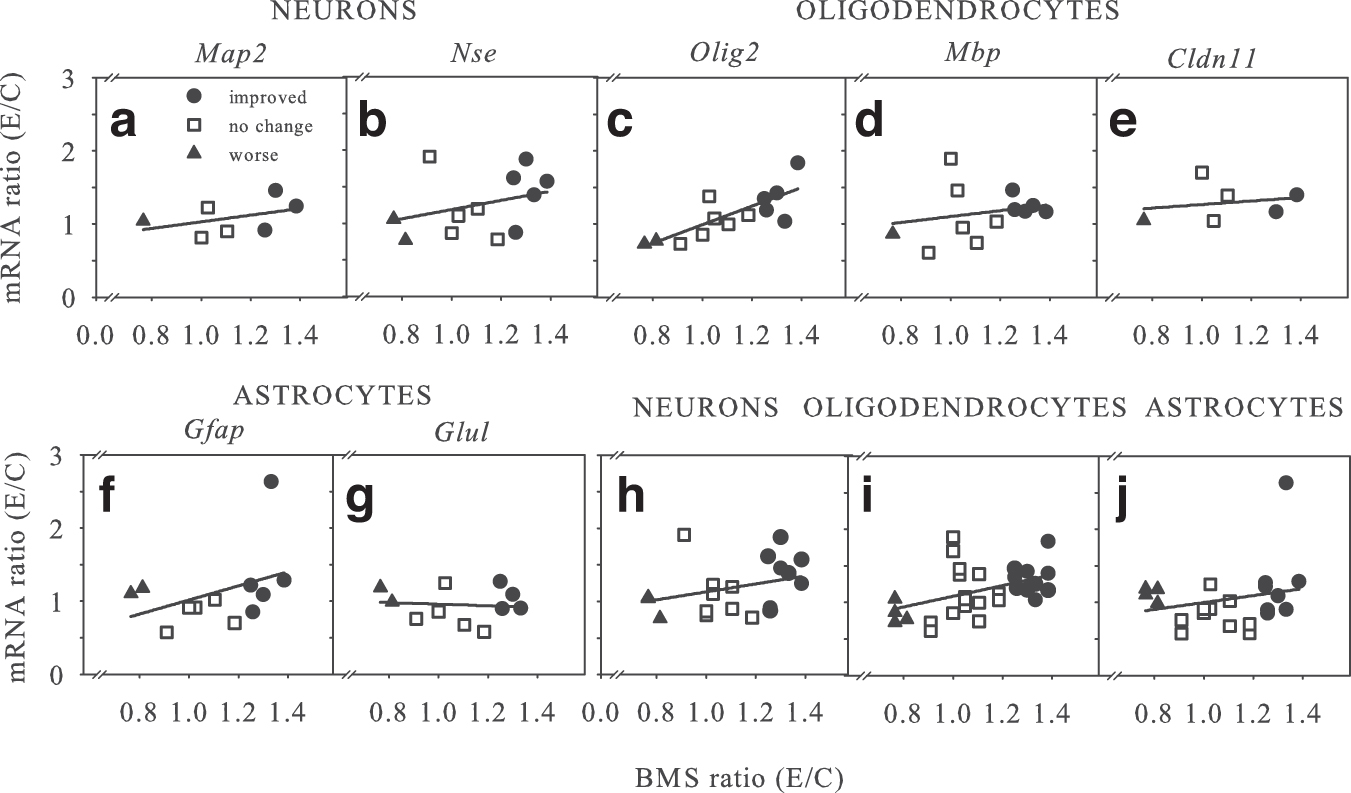
Correlation of acute neural messenger ribonucleic acid (mRNA) levels with chronic six-week Basso Mouse Scale (BMS) ratios. For all studies (Table 1), the ratio of individual acute neuron (a,b,h), oligodendrocyte (c-e,i), and astrocyte (f,g,j) mRNA levels for experimental/control conditions were graphed versus their respective ratio of E/C chronic BMS scores. For all graphs, studies with ‘improved’ (•), ‘no change’ (
To determine whether grouped acute neural E/C mRNA data might be predictive of chronic locomotor recovery, data were analyzed as above for ERSR mRNAs. Combining these data using the worse, no change, and improved BMS outcome grouping shows a significant difference in the improved outcome groups for both neurons (p < 0.05) and oligodendrocytes (p = 0.05) with BMS outcome ratios, but no significant differences in the worse or no change groups; astrocyte data were not significant (data not shown). Thus, as with the ERSR mRNAs, neural mRNA ratios were not accurately predictive across all functional recovery outcome groups.
In all studies that showed chronic behavioral improvement (Chop-/-, Sal, Hri-/- , GSK, and Pde4b-/- ), however, there were increased neural and reduced ERSR acute mRNA levels. Those studies with no improvement (Gnb, Gcn2-/-, Atf6-/-, AQ, Gadd34-/-, and Pkr-/- ) showed no change, while those with worse chronic locomotor recovery (OL-Xbp1 -/- and OL-Perk-/- ) had decreased neural and increased E/C ERSR acute mRNA levels, respectively.
To determine whether both sets of acute data could be combined to generate a predictive measure of chronic behavioral recovery, we plotted the value of neuronal (Fig. 3a), oligodendrocyte (Fig. 3b), or astrocyte (Fig. 3c) E/C mRNA ratio divided by the E/C ratio of ERSR mRNA versus their respective ratios of E/C chronic BMS scores. Regression analyses showed significance (p < 0.05, p < 0.01, p < 0.05) for all three comparisons, respectively. Moreover, the pharmacological/genetic interventions that increased (•), decreased (▴), or had no significant effect (□) on locomotor recovery nicely grouped together, especially for oligodendrocyte mRNAs (Fig. 3b).
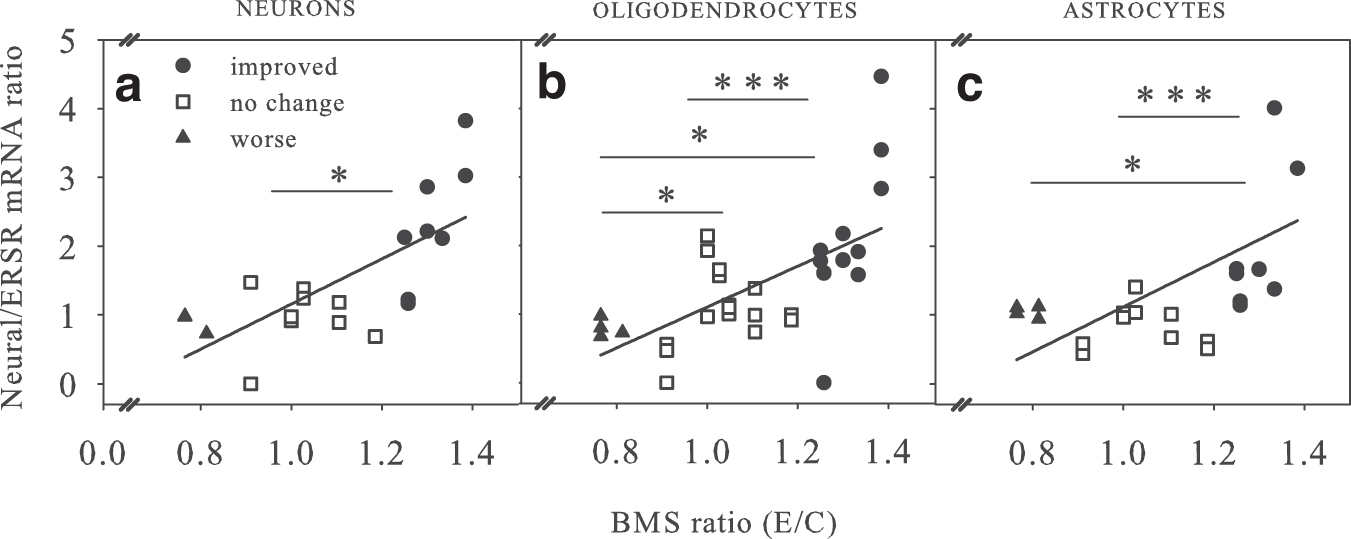
Correlation of the ratio of acute neural/endoplasmic reticulum stress response (ERSR) messenger ribonucleic acids (mRNAs) with chronic Basso Mouse Scale (BMS) scores. For all studies (Table 1), the ratios of individual acute neural mRNA levels – neuron (
Analysis using non-parametric Median tests showed that for neurons (Fig. 3a), the improved was distinct from the no change group, but not the worse group, likely because the group size of the latter was only two. For oligodendrocytes (Fig. 3b), all three groups were significantly different from each other. For astrocytes (Fig. 3c), the improved group was distinct from both no change and worse groups.
Grouping E/C neural/ERSR mRNA ratios into improved (•), worse (▴), or no change (□) data sets and analyzing using parametrical statistics showed that neuron, astrocyte, and oligodendrocyte groups all showed significant differences between improved with the no change groups (p ≤ 0.05), while astrocyte ratios were not significant. The inability of these latter analyses to be predictive across all recovery levels, as discussed below, is likely reflective of the small n in the worse group.
Discussion
In the context of CNS dysfunction, proteostasis has been most thoroughly investigated in aging 14,34 and neurodegenerative disease. 35 –38 Given the accumulation of neurotoxic aggregates in the latter, it is not surprising that the proteostasis network is a target of active and evolving drug development for those disorders. 39 With respect to CNS trauma, the contributions of both autophagy 40,41 and the ERSR 15,16 to neuropathology have been reviewed recently. Pathological activation of the ERSR has also been implicated in traumatic brain injury, 42 –47 intracerebral hemorrhage, 33 and stroke. 48,49 –51 Our work has demonstrated directly that both the ERSR 19 –22,24.32,52 and autophagy 23 are implicated in the acute pathological responses after SCI and that ameliorating those responses can lead to enhanced functional recovery.
That work and a number of unpublished studies led to the hypothesis that abrogation of the acute ERSR concomitant with reduced loss of neural mRNAs could be predictive for chronic locomotor recovery after contusive thoracic SCI. Data presented in Figs. 1–3 support that hypothesis. While we recognize that this assay is predominantly correlative, it has been consistent in all of the evaluations that we have undertaken thus far with thoracic contusive SCI. This assay has evolved over the course of these 13 studies, which represents more than a decade of work. The genes that we now routinely screen are Atf4, Chop, Map2, Nse/Eno2, Olig2, Mbp, Gfap, and Glul.
With respect to the statistical analyses used, the data are much more robust for the improved group because the number of studies that showed therapeutic effects was larger (n = 5) than those that showed worse functional improvement (n = 2). The variability of the no change data set by its very nature also added to intrinsic variability. This was evident when neural or ERSR mRNAs were analyzed alone. The overall significance of the combined data sets was robust, but could not be predictive across the full range of locomotor recoveries. Even in the combined data set (Fig. 3), it was only non-parametrical analyses that fully evidenced the predictive ability of this approach.
Given addition of future studies that show significantly worse functional recovery to increase the sample size of the data set, the oligodendrocyte/ and neuronal/ERSR mRNA ratios would likely provide a more accurate forecast across the full range of behavioral outcomes. We are confident in our current data, however, that this acute in vivo analysis is very accurate in predicting which treatments will likely prove to be beneficial for chronic locomotor recovery.
We recognize that in the current data set, we are pharmacologically or genetically modulating the exact signaling pathways (ERSR) that we are using as one of our output measures, thus potentially biasing our results. Proteostasis responses, however, are the end signaling pathway for all pathophysiological cellular stresses in all affected cells. 14 We contend that any pharmacological or genetic intervention that enhances functional recovery after SCI must necessarily restore cellular homeostasis in those cell populations responsible for that recovery that would otherwise initiate apoptotic responses and subsequently die or remain dysfunctional.
Supporting this contention, the Pde4b-/- study 32 was mechanistically independent of initial ISR/ERSR/UPR signaling modulation because it involves TLR4-mediated inflammatory responses both peripherally 53,54 and centrally. 32,55 Yet, in that case, attenuated acute induction of Chop and Atf4 mRNAs was associated with chronically improved BMS. 32 Importantly, the ISR/ERSR affects other components of acute/subacute injury response including neuroinflammation and scar formation. 56 –60 Neuroinflammation is one of the major drivers of delayed white matter injury after SCI while scar formation may reduce the potential for neuronal plasticity-driven recovery. 61,62 Therefore, manipulating the ISR/ERSR may modulate functional recovery after SCI not only by directly influencing acute cell loss but also by affecting delayed white matter damage and/or post-injury plasticity.
We have not tested this approach in cervical or lumbar contusive SCI where neuronal loss is of paramount importance to the extent of locomotor recovery. In our data set that was primarily focused on studies that manipulated ERSR/proteostasis, however, neuronal sparing correlated with functional recovery while being predictive of improved recovery (Fig. 3a). Moreover, we observed upregulation of ERSR/UPR markers in neurons after SCI. 19,20 In those studies of thoracic contusive SCI where neuronal loss does not correlate with functional recovery, 8,10 this likely reflects the critical role of proteostasis for overall neural tissue sparing. Hence, we anticipate that similar changes of neuronal marker and ERSR/UPR mRNA expression would reflect functional improvement in injuries that target lumbar or cervical enlargements.
We have also used this approach to confirm the in vivo biological action of multiple injected drugs after SCI: salubrinal, 19 guanabenz, 21 rapamycin, spautin-1, 23 GSK2656157, and adaptaquin (unpublished data). If changes in neural and/or ERSR mRNAs are seen after drug infusion, one can assume both penetrance across the blood–brain/spinal cord barriers and biological activity in the CNS. We suggest that this assay would also be useful to optimize dose and therapeutic window after SCI. Relative to the time-consuming analysis of behavioral analysis of novel therapeutic pharmacological compounds, this assay provides a relatively quick, yet still in vivo, approach to predict therapeutic efficacy. It should be useful for CNS trauma applications other than thoracic contusive SCI.
Additional studies with therapeutic effectors that do not directly affect proteostasis responses will be required to further validate this assay. We recommend that the effectiveness of acute interventions focused on other components of SCI pathogenesis such as oxidative stress, mitochondrial damage, or neuroinflammation may be best predicted by supplementing current tissue sparing mRNA markers with those that directly monitor the manipulated process.
Footnotes
Acknowledgments
The data presented here were supported by NS108529, NS073584, P20/P30 GM103507, Kentucky Spinal Cord and Head Injury Research Trust 08-15, Norton Healthcare, and the Commonwealth of Kentucky Challenge for Excellence. We thank Christine Yarberry for surgical assistance and Johnny Morehouse and Jason Beare for behavioral analyses.
Author Disclosure Statement
No competing financial interests exist.