
Abstract
The neurocognitive impairments associated with mild traumatic brain injury (TBI) often resolve within 1–2 weeks; however, a subset of people exhibit persistent cognitive dysfunction for weeks to months after injury. The factors that contribute to these persistent deficits are unknown. One potential risk factor for worsened outcome after TBI is a history of stress experienced by a person early in life. Early life stress (ELS) includes maltreatment such as neglect, and interferes with the normal construction of cortical and hippocampal circuits. We hypothesized that a history of ELS contributes to persistent learning and memory dysfunction following a TBI. To explore this interaction, we modeled ELS by separating Sprague Dawley pups from their nursing mothers from post-natal days 2–14 for 3 h daily. At 2 months of age, male rats received sham surgery or mild to moderate parasagittal fluid-percussion brain injury. We found that the combination of ELS with TBI in adulthood impaired hippocampal-dependent learning, as assessed with contextual fear conditioning, the water maze task, and spatial working memory. Cortical atrophy was significantly exacerbated in TBI animals exposed to ELS compared with normal-reared TBI animals. Changes in corticosterone in response to restraint stress were prolonged in TBI animals that received ELS compared with TBI animals that were normally reared or sham animals that received ELS. Our findings indicate that ELS is a risk factor for worsened outcome after TBI, and results in persistent learning and memory deficits, worsened cortical pathology, and an exacerbation of the hormonal stress response.
Introduction
Traumatic brain injury (TBI) is a major public health problem, with an estimated 1,600,000–3,800,000 concussions occurring annually in the United States. 1 –3 Mild TBI and concussion are the most common types of TBI, comprising 80% of all reported TBIs. 4 Although the majority of people with mild TBI recover within 10–14 days, a subset of patients exhibit persistent neurological symptoms, which could be caused by baseline differences at the time of injury. 5 –8 The factors that contribute to prolonged symptoms after mild TBI remain an area that needs investigation. One factor that may affect the degree of cognitive dysfunction experienced by a person after TBI is a history of stress.
Early life stress (ELS) is associated with learning and memory problems that emerge in middle age, increases susceptibility to psychopathology in adulthood, and alters the physiological response to subsequent stressors. 9 Chronic ELS is highly prevalent in the United States, and there are ∼700,000 reports of child maltreatment each year. 10,11 Cumulatively from 2004 to 2011, this represents a total of 5,689,000 confirmed reports of child maltreatment. 10 Of these reports, 75% of the children were neglected. 10 Chronic ELS in childhood, such as neglect, alters brain development, results in a higher risk for general and mental health problems, elevates levels of risky behavior, and increases the incidence of TBI. 12 –16
Clinical studies have clearly demonstrated several risk factors that are associated with worsened outcome after TBI, but whether exposure to chronic stress is a risk factor for worsened outcome after TBI has not been well established. 17 –21 Known risk factors for a poorer outcome after TBI include older age at the time of injury, history of a previous concussion, lower education level, low socioeconomic status, low pre-injury resilience, and prior psychiatric problems such as depression, anxiety, or post-traumatic stress disorder (PTSD). Some of these comorbid psychiatric problems may actually be related to early stressful life experiences. 22,23
To directly address whether stress is a risk factor for worsened outcome after TBI, a few clinical studies have investigated the association of stressful life events and mild TBI outcome. In a study of pediatric mild TBI patients, a history of prior life stress, but not depression, was associated with prolonged post-concussive symptoms. 24 Similarly, in a study of adult mild TBI patients admitted to an emergency room, a history of stressful life events was associated with worsened physical and mental health quality of life at 3 months post-injury. 25 This finding was replicated in a retrospective study of 165 United States military service members with mild TBI, which found that exposure to chronic stressful life experiences significantly correlated with persistent post-concussion symptoms. 26 Thus, chronic ELS and TBI may interact to result in significantly worsened consequences after TBI in this vulnerable population.
To model these clinical studies that have described an interaction between stress and TBI, most pre-clinical studies have utilized a paradigm of stress exposure immediately prior to and/or after the brain trauma in adulthood to model the relevant situation of a TBI encountered while in a stressful environment. However, it is unknown if stress encountered during early development is a predisposing factor to a worsened outcome after TBI. This is an important clinical question, given that the response to chronic stress in adulthood is different than the response to chronic stress in early development when the hypothalamic pituitary adrenal (HPA) axis is still developing. 27,28 The response to chronic stress in early development when the HPA axis and brain structures are still developing results in long-term alterations in neuronal circuitry as well as in microglia functioning. 27 –30 This suggests that understanding the interaction of ELS and TBI is important for a vulnerable patient population who may be at risk for persistent cognitive dysfunction. Therefore, this study is aimed at exploring the effects of chronic ELS on outcome after TBI. To determine if stress experienced during early development alters outcome after TBI, maternal separation, a well-characterized model of ELS in rodents, was used to model childhood neglect. This model of ELS increases measures of anxiety, impairs maternal care, and diminishes spatial navigation learning. 31 –34 This protocol of ELS has validity as a model of early childhood neglect in humans by recapitulating several features. 28 Both early childhood neglect in humans and this model of ELS in rodents resulted in a heightened immune response to stressors encountered in adulthood, 35 –37 increased corticotropin-releasing hormone activity, 38,39 elevated HPA axis response to acute stress in adulthood, 40 and impaired glucocorticoid feedback because of decreased glucocorticoid receptor levels in the hippocampus. 41,42 Therefore, in this study, we determined whether exposure to stress early in life limited recovery after TBI by evaluating its effect on learning and memory, cortical and hippocampal atrophy, and the hormonal stress response.
Methods
Animal groups
Experimental procedures were compliant with the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals and approved by the University of Miami Animal Care and Use Committee. For this study, 10 pregnant Sprague Dawley rats were obtained at E15 (Charles River Laboratories) and maintained on a 12/12 h light/dark cycle with food and water ad libitum. The pups were born in the facility and the day of birth was designated as post-natal day 0 (P0). Dams with litters were left undisturbed until P2, and litter size was culled to 10 pups to ensure standardized nutrition, and culled to an equal number of pups for each sex when possible. Dams produced a range of four to eight males per litter (median six males/litter). At P21, pups were weaned, and only males were used. A total of 47 male rats were used in this study (n = 13 Sham + Control [Ctl], n = 12 Sham + ELS, n = 11 TBI + Ctl, n = 11 TBI + ELS rats). For behavioral and histology assessments, eight Sham + Ctl, seven Sham + ELS, six TBI + Ctl, and six TBI + ELS rats were used. For corticosterone measurements, five rats per group were used.
Early life stress
To model ELS, maternal separation was used. 43 Animals were randomly assigned prior to the experiment to ELS or normal rearing. For ELS, from P2 to P14, dams were removed from their original cage and placed in a clean cage. Litters were left undisturbed in their home cage for 3 h daily between 9:00 a.m. and 12:00 p.m. in a separate room while maintained at normothermia using a heating pad. The litters in the control group remained undisturbed. All rats were socially housed until surgery.
Surgical procedures
At P60, male rats were a priori randomized to receive either parasagittal mild to moderate fluid-percussion injury (FPI) or sham surgery as described previously. 44 Anesthesia was induced with 3% isoflurane, 70% N2O, and 30% O2, and animals were maintained with 1–2% isoflurane, 70% N2O, and 30% O2. A 4.8 mm craniotomy (-3.8 mm bregma, 2.5 mm lateral) was prepared by trephination over the right parietal cortex, and an 18 gauge syringe hub was secured to the craniotomy window with cyanoacrylate and dental cement. At 16–18 h after the craniotomy surgery, the animals were re-anesthetized (induction with 3% isoflurane, 70% N2O, and 30% O2 and maintained with 1–2% isoflurane, 70% N2O, and 30% O2), intubated, and mechanically ventilated (Stoelting, Wood Dale, IL). A catheter was placed in the tail artery to monitor mean arterial blood pressure (MABP). Arterial blood samples were obtained to measure blood gases (pO2 and pCO2) and blood pH. After stabilization of physiological parameters, animals received a mild to moderate parasagittal FPI (1.4–1.6 atm) or sham injury. Penicillin/benzathine (20,000 IU/kg, intramuscular) was given once prior to the brain injury surgery, and buprenorphine (0.01 mg/kg, subcutaneously) was given once at the completion of the surgery. Physiological variables of MABP, blood pO2, pCO2, pH, and head and body temperature were controlled throughout the surgery (Table 1). A significant interaction of animal treatment × time was observed for pO2 (F [3,43] = 3.91, p = 0.015), with a slight decrease observed in all animal treatment groups at the end of surgery. Additionally, a significant main effect of time was observed for pCO2 (F [1,43] = 42.39, p < 0.001) and pH (F [1,43] = 13.92, p < 0.001). However, blood pO2, pCO2, and pH remained within normal physiological ranges throughout the duration of the surgery for all animal treatment groups. A priori exclusion criteria were: mortality, >15% loss of body weight, non-resolving infection at a surgical site, inability to feed or drink, motor paralysis, listlessness, self-mutilation, excessive grooming leading to loss of dermal layers, excessive spontaneous vocalization when touched, or poor grooming habits. 45 For the first 2 days following surgery, animals were monitored daily, then weighed and evaluated every 2 weeks until decapitation or perfusion. As expected, there was a main effect of time for body weight (F [1,42] = 1984.34, p < 0.001), with all animal treatment groups having a significant weight gain at the time of perfusion compared with the start of surgery. There were no significant differences in body weight among non-stressed and ELS animals at the time of surgery (P60) or at the time of perfusion (P120).
Physiological Data
p < 0.05, *** p < 0.001 versus time at surgery or 15 min prior to fluid-percussion injury (FPI), repeated measures two-way analysis of variance (ANOVA) with Tukey's honestly significant difference (HSD) post-hoc test. Mean ± standard error of the mean (SEM), n = 13 Sham+Ctl, n = 12, Sham+ELS, n = 11 TBI+Ctl, n = 11 TBI+ELS.
ATM, atmospheres of pressure; MABP, mean arterial blood pressure; pO2, partial arterial oxygen pressure; pCO2, partial arterial carbon dioxide pressure; TBI, traumatic brain injury; ELS, early life stress.
Contextual fear conditioning
At 2 weeks post-surgery, contextual fear conditioning was assessed as previously described. 46 On day 1, the animals were habituated to the fear conditioning apparatus for 10 min. At 24 h after habituation, animals were fear conditioned by pairing a tone (75 dB, 2.8 kHz, 30 sec) with a modest foot shock (1 mA, 1 sec) occurring at 120 sec, and then the animals remained in the apparatus for an additional 60 sec. The total duration of the training trial was 3 min. Contextual fear conditioning was assessed at 24 h as well as at 1 month after training, with a 5 min trial duration. Freezing was quantified by video analysis (Freeze Frame 3.32, Coulbourn Instruments, Holliston, MA).
Water maze
At 3 weeks post-surgery, water maze training and testing were performed. 46 The animals were trained over the course of 4 acquisition days to find a hidden platform in a constant location (four trials/day, 4 min inter-trial interval, 1 min maximum trial duration). Retention was assessed with a probe trial (1 min duration) at 24 h after the last acquisition day. Performance was assessed through video tracking and analysis (EthoVision XT 11.5, Noldus Information Technology, Leesburg, VA). Path length to reach the platform and time spent in each quadrant of the pool were quantified.
Spatial working memory
At 4 weeks post-surgery, animals were assessed for spatial working memory. This was performed for 2 days using a delayed match-to-sample task in the water maze. 46 Animals were trained to locate a hidden platform in the water maze that remained constant between two paired trials separated by 5 sec (1 min maximum trial duration). Four paired trials were averaged (4 min inter-trial interval). Animals were video tracked and path length differences between the first sample trial and subsequent match trial were measured (EthoVision XT 11.5). Data shown are from day 2 of testing.
Histology
At 8 weeks post-surgery, animals were anesthetized (3% isoflurane, 70% N2O, 30% O2, 5 min) and transcardially perfused with saline and 4% paraformaldehyde in 0.1M phosphate buffer, pH 7.4. The brains were embedded in paraffin and sectioned in a stereological series (10 μm thick, 150 μm apart) using a microtome (Leica RM2125 RTS, Buffalo Grove, IL) as previously described. 47 Sections were stained with hematoxylin and eosin (H&E) plus Luxol fast blue. Using NeuroLucida software (11.11.2, MicroBrightField) the ipsilateral and contralateral parietal cortex and hippocampus were contoured between bregma levels -3.3 to -6.8 mm. Atrophy was evaluated by quantifying and comparing contralateral and ipsilateral volume differences and normalizing to the contralateral volume.
Corticosterone measurements
At 8 weeks post-surgery, a separate set of animals that did not undergo behavioral testing were assessed for basal and stress-induced corticosterone levels in blood serum. Animals received 30 min of restraint stress by immobilization in Plexiglas tubes. Plasma from the tail vein was obtained from animals, and corticosterone was measured using ELISA (Enzo Life Sciences, Ann Arbor, MI). Baseline levels prior to restraint, immediately after 30 min of restraint, and 90 min after recovery from restraint were compared.
Statistical analysis
Data presented are mean ± standard error of the mean (SEM) and significance was designated at p < 0.05. Statistical comparisons were made using GraphPad Prism 7.0 and SigmaPlot 14.0. Two-way repeated measures analysis of variance (ANOVA) or two-way ANOVA were used to determine significance between the factors animal treatment × trial/day/quadrant/time or animal surgery × stress treatment. Tukey's post-hoc tests were used when significant interactions were present. A Student's t test was used to analyze atmospheres of pressure (ATM) levels between TBI+Ctl and TBI+ELS animal groups.
Results
Effect of ELS on learning and memory deficits after TBI
To determine if ELS is a factor contributing to worsened outcome after TBI, animals received ELS from P2 to P14, and then sham surgery or mild to moderate FPI in young adulthood. During the subacute recovery period from 2 weeks to 1 month post-surgery, animals were assessed on a series of behavioral tasks.
Beginning at 2 weeks post-surgery, we assessed for the presence of hippocampal-dependent learning deficits using contextual fear conditioning. Similar to what has previously been reported, contextual fear conditioning was normal in non-stressed TBI rats 44 as well as in ELS sham rats (Fig. 1). 48,49 However, in rats that received the combination of ELS and TBI, retention of contextual fear conditioning at 24 h after training was significantly impaired (animal treatment × trial interaction F [6,46] = 3.80, p = 0.004). These contextual fear conditioning deficits were persistent and also present when re-assessed 1 month after training. These results demonstrate that the interaction of ELS and TBI impairs hippocampal-dependent learning and memory retention at 24 h and 1 month after acquisition.
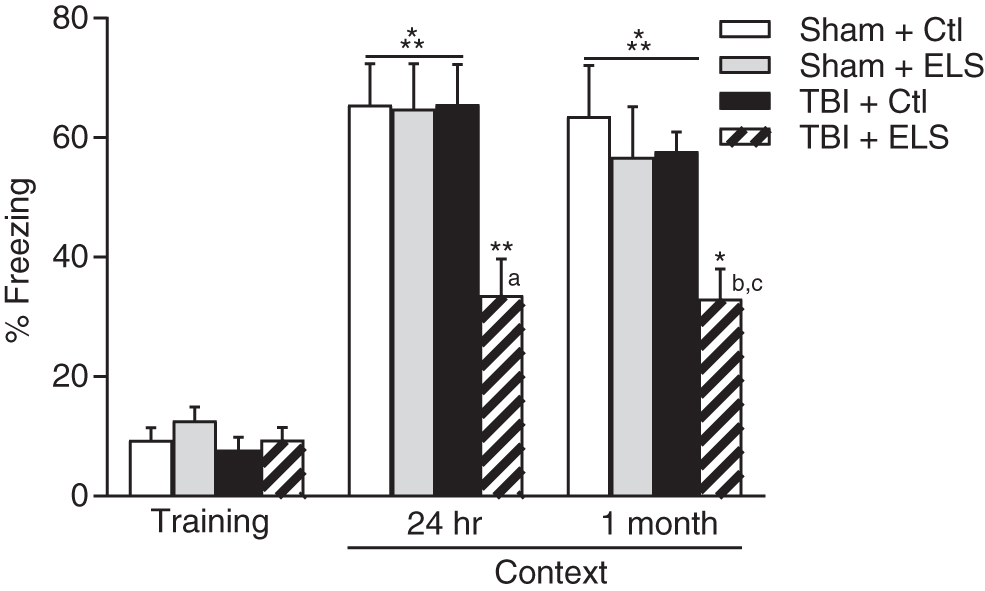
Deficits in contextual fear conditioning after early life stress (ELS) and traumatic brain injury (TBI). TBI+ELS animals exhibited significantly less contextual fear conditioning than Sham+Ctl, Sham+ELS or TBI+Ctl animals when assessed at 24 h or 1 month after training. *p < 0.05, **p < 0.01, ***p < 0 .001 versus training; a p < 0.01 versus Sham+Ctl, Sham+ELS, TBI+Ctl; b p < 0.01 versus Sham+ELS, TBI+Ctl; c p < 0.001 versus Sham+Ctl; repeated measures two-way analysis of variance (ANOVA) with post-hoc Tukey's test. Mean ± standard error of the mean (SEM), n = 8 Sham+Ctl; n = 7 Sham+ELS; n = 6 TBI+Ctl; n = 6 TBI+ELS.
To determine the effects of ELS and TBI in another hippocampal-dependent learning paradigm, animals were trained in the water maze task to find a hidden platform using spatial cues at 3 weeks post-surgery (Fig. 2A). Neither TBI nor ELS resulted in a significant difference in the distance traveled to locate the hidden platform across all 4 acquisition days. Memory retention was evaluated using a probe trial. At 24 h after the last training day, the hidden platform was removed and animals were placed back into the water maze to search for the missing platform (Fig. 2B). Both sham and TBI animals that received ELS exhibited significantly reduced time spent in the target quadrant whereas non-stressed sham and TBI animals exhibited significant memory retention of the platform location (animal treatment × quadrant interaction F [9,84] = 7.96, p < 0.001).
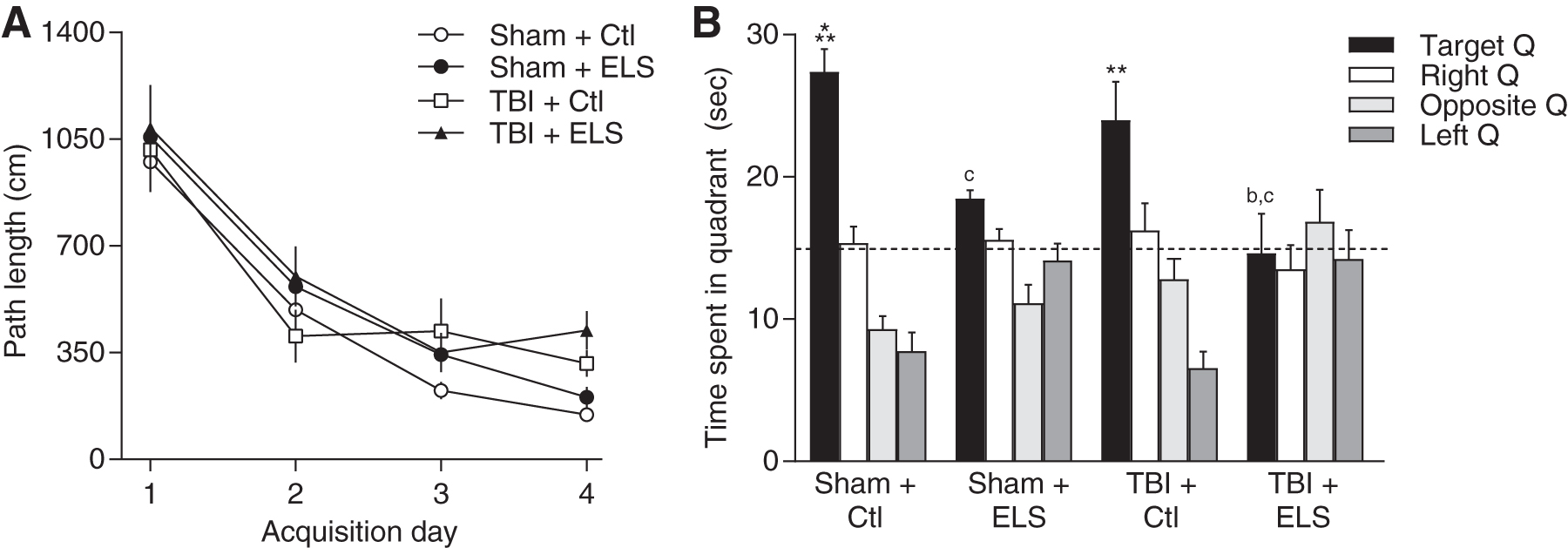
Water maze deficits after traumatic brain injury (TBI) and early life stress (ELS).
To assess the effects of ELS on spatial working memory after TBI, we used a delayed match to sample version of the water maze task. At 4 weeks post-surgery, the path length to find the platform in the sample trial versus the match trial was measured (Fig. 3). A positive difference between the sample and match trials in path length to find the platform is indicative of working memory ability. 50 Both non-stressed and ELS sham animals exhibited a significant difference in path length to find the platform in the sample versus the match trial. However, there was a significant impairment resulting from TBI. Both non-stressed and ELS TBI animals exhibited significant impairment in spatial working memory (main effect of surgery F [1,23] = 11.72, p = 0.002).
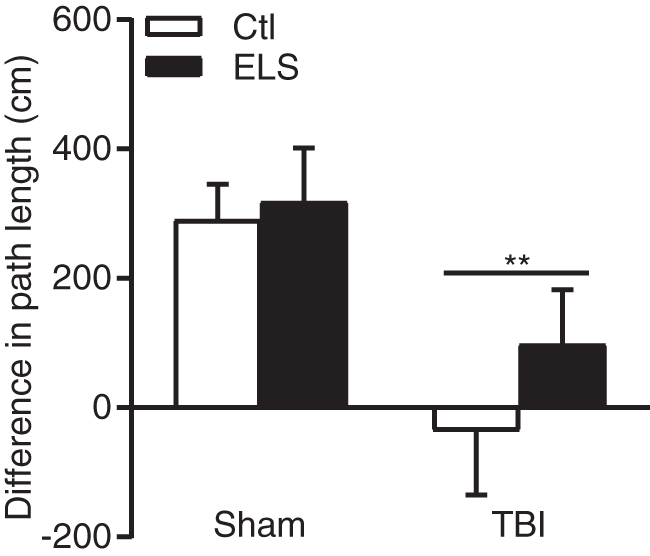
Spatial working memory impairment in traumatic brain injured (TBI) animals. Neither TBI+Ctl nor TBI+early life stress (ELS) animals exhibited a decrease in path length to find the platform in the match trial versus the sample trial, indicating impaired working memory after TBI. **p < 0.01 versus Sham+Ctl, Sham+ELS, repeated measures two-way analysis of variance (ANOVA) with post-hoc Tukey's test. Mean ± standard error of the mean (SEM), n = 8 Sham+Ctl; n = 7 Sham+ELS; n = 6 TBI+Ctl; n = 6 TBI+ELS.
Effect of ELS on atrophy after TBI
At the completion of the behavioral testing, cortical and hippocampal atrophy was assessed (Fig. 4). This model of mild to moderate parasagittal FPI results in modest cortical atrophy and has no significant effect on hippocampal atrophy. 51 In this current study, there was a small, significant increase in hippocampal atrophy in both non-stressed and ELS animals that received TBI (main effect of surgery F [1,23] = 4.52, p = 0.045). Additionally, there was a small, but significant increase in hippocampal atrophy in both sham and TBI animals that received ELS (main effect of stress F [1,23] = 5.15, p = 0.033). However, there was no significant interaction of surgery and ELS for hippocampal atrophy (surgery × stress interaction F [1,23] = 2.76, p = 0.111). In contrast, there was a significant increase in cortical atrophy after TBI (main effect of surgery F [1,23] = 93.63, p < 0.001) that was exacerbated by the exposure to ELS (surgery × stress interaction F [1,23] = 6.23, p = 0.020). These results suggest that cortical atrophy was significantly increased by a combined effect of ELS and TBI.
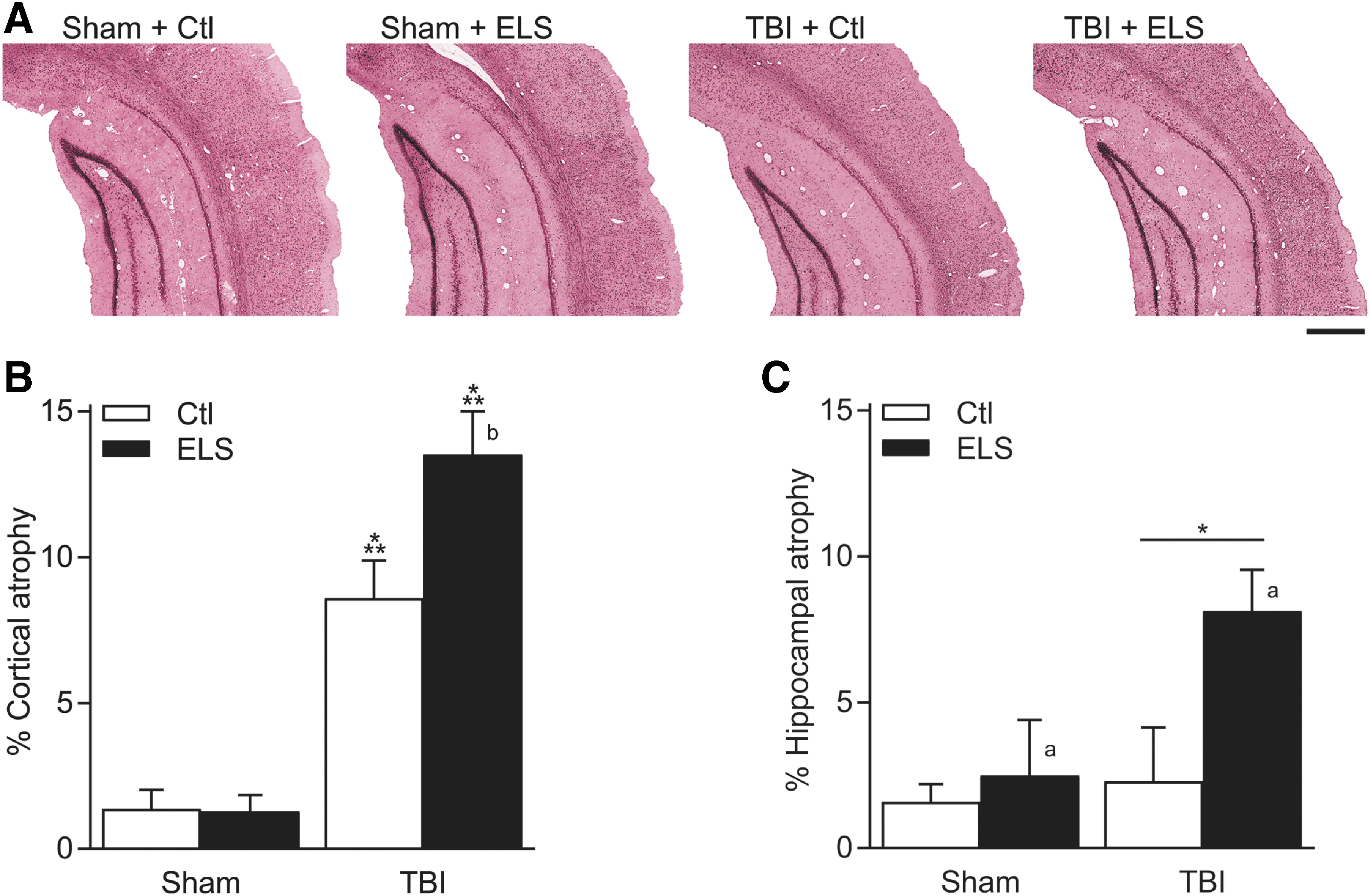
Exacerbation of cortical atrophy after traumatic brain injury (TBI) by early life stress (ELS).
Corticosterone expression in the interaction of ELS and TBI
ELS results in hyperactivity of the HPA axis in adulthood as exemplified by increased levels of corticosterone in response to stress. 52,53 TBI alone, like ELS, potentiates the increase in stress-induced corticosterone. 54 –57 Similar to these studies, when we evaluated the HPA axis using restraint stress at 8 weeks post-surgery, we found that basal levels of corticosterone in the plasma were similar for all groups (Fig. 5). Likewise, the rise in corticosterone during restraint stress was comparable among all groups. When non-stressed sham animals returned to their home cage, their corticosterone levels began to decrease. In both ELS sham animals and non-stressed TBI animals, there was a persistent elevation of plasma corticosterone during the recovery period after the restraint stress. However, in animals that received ELS and TBI, corticosterone levels continued to rise significantly when they were returned to their home cage, compared with ELS alone or TBI alone (animal treatment × time interaction F [6,30] = 5.88, p < 0.001). At 90 min after recovery from restraint stress (Fig. 5B), corticosterone was significantly elevated in both non-stressed and ELS animals that received TBI (main effect of surgery F [1,16] = 9.32, p = 0.008). Additionally, a significant increase in corticosterone was measured in both sham and TBI animals that received ELS (main effect of stress F[1,16] = 23.76, p < 0.001). These results suggest that the combined effect of ELS and TBI result in a prolonged elevation of glucocorticoids in response to stress.
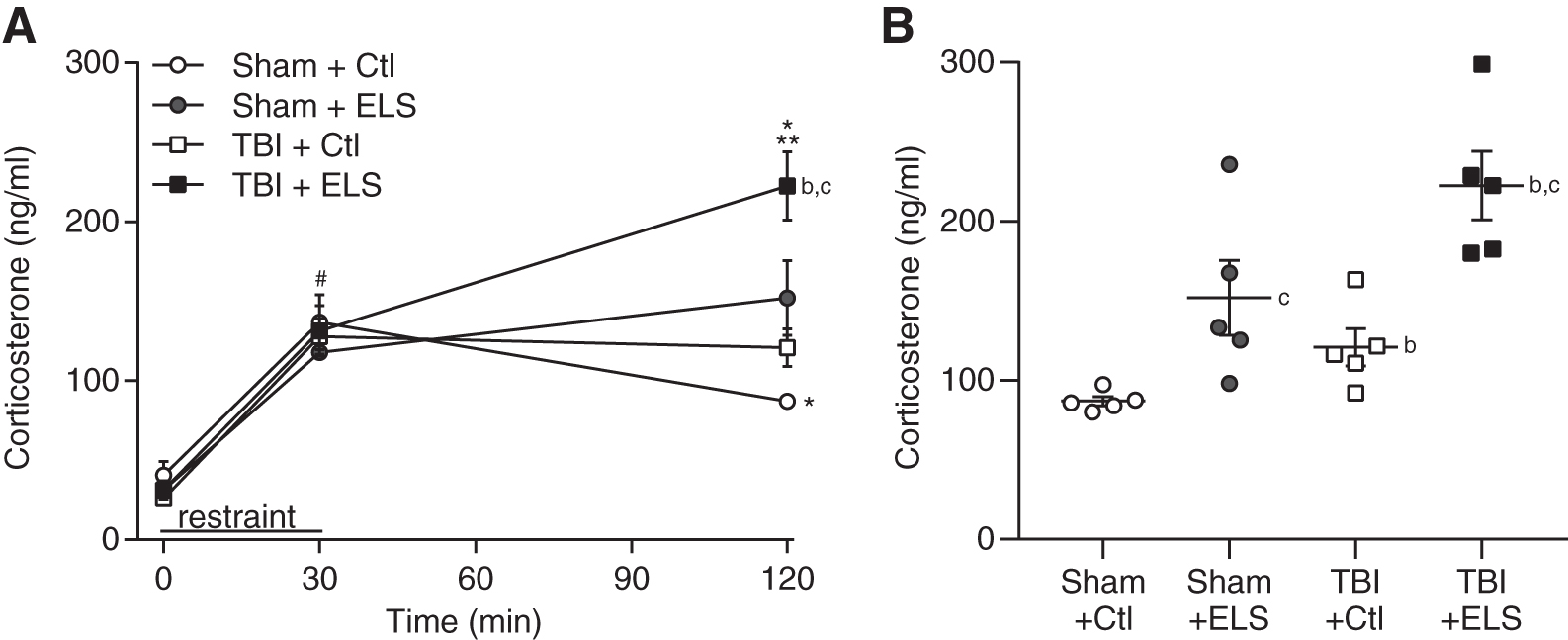
Prolonged elevation of corticosterone after traumatic brain injury (TBI) and early life stress (ELS).
Discussion
Mild TBI alone results in modest behavioral symptoms that typically resolve within 10–14 days. 5 However, a subset of individuals report long-term symptoms that take weeks to months to resolve. 5 –8 The aim of this study was to determine if ELS during development limits the recovery trajectory after a TBI that occurs in adulthood. Although several pre-clinical studies have investigated the interaction of stress and TBI when the stressors occur during adulthood, 24,25 it is still unknown if stress encountered during early development is a predisposing factor to a worsened outcome after TBI. Having an understanding of the effects of the interaction of ELS and TBI is important for a vulnerable patient population who may be at risk for persistent cognitive dysfunction following brain injury. Using maternal separation in rats to model ELS, we found that a mild FPI in adulthood after ELS impaired contextual fear conditioning retention as well as memory retention in the water maze and spatial working memory, resulted in cortical atrophy, and exacerbated the hormonal stress response.
Maternal separation is a well-characterized model for ELS in rodents. 27 This protocol of brief, daily maternal separation from P2 to P14 was selected because this is the period of HPA axis maturation in rodents. 58,59 The HPA axis in rodents matures during the first 3 post-natal weeks and is adult-like by P30. 60,61 This period is comparable in human brain development to the last 1–2 months of gestation until ∼4–5 years post-natally. 45,62 We also chose this model because this is the age when both the cortex and hippocampus are developing, as are the circuits involved in the learning and memory tasks evaluated in this study. 63,64 Validity for this model of ELS is supported by similar behavioral impairments in both rodents and humans. This paradigm of ELS impairs several types of hippocampal-dependent learning and memory tasks in adulthood. 43,65 Comparably in humans, early child maltreatment is associated with cognitive impairments in adulthood, affecting attention, working memory, executive functioning, processing speed, and visual episodic memory. 66 In measures of stress and anxiety, ELS in rodents increases anxiety-like behavior in adulthood, such as reducing time spent in the open arms of the elevated plus maze. 27,43 This phenotype is similar to what has been observed in human studies, which report an increased relative risk for developing anxiety disorders in adulthood after childhood maltreatment. 40 Thus, this model of ELS results in behavioral changes in adulthood that have commonalities to what has been observed in humans.
It is important to note that previous studies using this same model of ELS have found that body weight is lower in maternally separated pups than in those with normal rearing from P1 to P15; however, this difference becomes non-significant by P30. 52,53 In order to limit the effects of additional handling, we did not measure pup weights during the maternal separation period in our study. However, there were no significant differences in body weight between non-stressed and ELS animals at the time of surgery (P60) or at the time of perfusion (P120). This suggests that weight was not a significant factor in the differences to the response of the TBI surgery. A limitation of this study is that only male rats were evaluated. We chose to focus on males to highlight the effects of ELS on outcome after TBI, as female rats are known to be less sensitive to the effects of ELS for some learning tasks. 27 ELS impairs water maze retention and spatial working memory in male rats, but not in female rats. 67 For contextual fear conditioning, sex differences between males and females after ELS are not as apparent. 68,69 Future studies are needed to determine whether ELS also worsens outcome in females after TBI.
Several studies have investigated the interaction of stress and TBI utilizing a paradigm of stress exposure immediately prior to and/or after the brain trauma in adulthood to model the clinically relevant situation of a TBI encountered while in a stressful environment. These studies have found that exposure to stress immediately prior to TBI increases microglia cell numbers in the cortex and hippocampus, decreases CA3 neuronal survival, impairs hippocampal long-term potentiation, and increases corticosterone levels. 54,70 –78 One developmental study investigated the effects of intermittent social isolation and a high-fat diet during adolescence on outcome after TBI and reported impaired novel object recognition. 79 However, whether stress encountered during early development is a predisposing factor to a worsened outcome after TBI is important to address. This is a relevant clinical question because the response to chronic stress in adulthood is very different than the response to chronic stress in early development, when the HPA axis is still developing. 27,28 For example, stress encountered during the first 5 years of life, but not during middle childhood or adolescence, interacts with stress encountered in adulthood to dysregulate diurnal cortisol levels. 61 Therefore, although several studies have investigated the interaction of stress at the time of injury and TBI in adulthood, the impact of ELS on outcome after TBI remains an area that greatly needs investigation.
To evaluate whether ELS results in prolonged cognitive deficits following a TBI in adulthood, animals received ELS during the first 2 weeks post-natally and then a mild to moderate TBI in young adulthood. At 2 weeks post-injury, the animals were trained and tested on contextual fear conditioning. This learning task was selected because ELS in uninjured animals has no effect on the acquisition or retention of contextual fear conditioning. 49,80 Similarly, our model of TBI at this injury severity does not result in any observable contextual fear conditioning deficits, nor are there shock sensitivity changes. 46,51,81 ELS also does not result in alterations in foot shock sensitivity. 82 For general activity and anxiety levels, mild TBI or ELS alone does not alter distance moved in the open field at this recovery time point. 27,43,83 –85 However, when ELS occurred prior to TBI, contextual fear conditioning retention at 24 h after training was significantly impaired. A clinically relevant finding of our study is that the TBI animals that received ELS showed persistent impairments in fear conditioning even 1 month after training. Therefore, our results suggest that ELS could be a contributing factor for prolonged cognitive symptoms after TBI.
Interestingly, the interaction of ELS and TBI on performance in the water maze task and spatial memory task yielded mixed results. In accordance with previous studies, neither ELS nor TBI at this injury severity had a significant effect on acquisition of the water maze task. 51,65 Similarly, animals that received ELS and TBI had normal water maze acquisition. When animals were assessed for retention of the platform location, ELS animals from both sham and TBI groups were impaired. Prior studies with this ELS paradigm have revealed similar deficits in retention, but not acquisition of the water maze task. 65 These results suggest that with this learning and memory task, the effects of ELS were predominant in causing a retention deficit. In contrast, spatial working memory was significantly impaired after TBI, but not with ELS alone. Both normal reared TBI animals and animals that received ELS and TBI were impaired, suggesting that spatial working memory is particularly sensitive to the effects of TBI. 51 Together, these results suggest that the interaction of ELS and TBI affects cognitive functioning in a complex manner, requiring several different memory tasks to delineate the deficits accorded by TBI and ELS.
These learning and memory deficits after ELS and TBI were associated with worsened pathology. This mild to moderate parasagittal FPI model results in modest cortical atrophy, which was also observed in our study. 51 However, the combination of ELS and TBI significantly exacerbated cortical atrophy. We also observed a modest increase in hippocampal atrophy caused by ELS as well as TBI, but no significant interaction between TBI and ELS. This result may be reflective of the recovery time point of 8 weeks. A previous study measuring hippocampal volume in ELS animals at a similar time point of 2–3 months of age found no significant hippocampal atrophy caused by ELS. 65 It is possible that the effects on hippocampal atrophy are not fully realized at this assessment time point considering that the animals were young adults. Another limitation of our pathology analysis is that more subtle pathologies within the hippocampus were not assessed, such as neuronal loss or dendritic changes. ELS is well established to decrease hippocampal dendritic complexity and length, which is evident in middle-aged, but not in young adult rats. 86,87 Although moderate FPI decreases synaptic density, 88 mild TBI only transiently decreases dendritic length and complexity. 89 Therefore, it is possible that the combination of ELS and TBI could interact to decrease hippocampal dendritic complexity or length, or alter spine numbers.
In our studies, we found that restraint stress persistently increased corticosterone levels in sham ELS and normal reared TBI animals. This corresponds to prior studies that have reported that ELS results in hyperactivity of the HPA axis in adulthood. 52,53 Although basal corticosterone levels in adulthood are unaltered, stress-induced levels of corticosterone are elevated. 40,90,91 Mild and moderate TBI alone, like ELS, potentiates the increase in corticosterone after stress. 54 –57,78 However, in TBI animals that received ELS, corticosterone levels continued to rise during the recovery period after restraint stress, and this persistent increase was significantly greater than in TBI-only or ELS-only animals. These disturbances in negative feedback regulation of the HPA axis may be a mechanism underlying the increased vulnerability in TBI animals that received ELS. Glucocorticoid receptors play an important role in negative feedback of the HPA axis when circulating glucocorticoid levels are high. 92,93 Given its density of glucocorticoid receptors, the hippocampus is highly involved in negative regulation of the HPA axis. 94 Some studies have reported that ELS decreases hippocampal glucocorticoid receptor levels in adulthood. 95 –97 A reduction in glucocorticoid receptor levels in the hippocampus could decrease negative feedback of the HPA axis to the hypothalamus. 41,42 Therefore, ELS and TBI may have interacted to decrease glucocorticoid receptor expression. This could have attenuated negative feedback on the HPA axis, potentially explaining the persistent elevation of corticosterone in ELS TBI animals after restraint stress.
Glucocorticoids modulate learning and memory processes in animals and humans in a complex manner, with elevated levels of glucocorticoids both enhancing and impairing learning and memory. A transient increase in circulating glucocorticoids in association with a learning task facilitates memory consolidation. 98 –100 In contrast, memory retrieval processes are usually impaired with high circulating levels of glucocorticoids. 34,101 –103 Associated with these memory deficits are impairments in hippocampal long-term potentiation. 101,102,104 This indicates that activity-dependent strengthening of synapses, as is thought to occur during learning and memory formation, is impaired with elevated glucocorticoid levels. 105 Therefore, we speculate that the learning and memory deficits we observed in ELS TBI animals may have been caused by increased corticosterone levels.
A key mechanism linking changes in the HPA axis after ELS to worsened behavior and pathology after TBI may be elevated inflammation. 106 ELS elevates corticosterone in pups during the separation period. 107 –109 Additionally, ELS increases microglia density in the hippocampus, and this increase persists through young adulthood. 110 –112 Microglia express high levels of glucocorticoid receptors 113,114 and respond to increased levels of corticosterone by proliferating and increasing in phagocytic activity and motility. 111,112,115 Although glucocorticoids are anti-inflammatory, if stress occurs prior to a second challenge such as peripheral or central nervous system (CNS) immune stimuli, the neuroinflammatory response is potentiated. 116,117 Therefore, it is possible that there is an additive or synergistic effect between inflammation and stress, resulting in higher microglial activation and expression of proinflammatory molecules after TBI. 95 This suggests that exposure to ELS prior to a TBI may have increased inflammation and contributed to the exacerbation of cortical pathology observed after ELS and TBI.
Conclusion
In summary, our findings indicate that ELS limits the recovery trajectory after a TBI that occurs in adulthood. The combination of ELS and TBI resulted in persistent learning and memory deficits, worsened cortical pathology, and an altered neuroendocrine stress response. These results are important in recognizing that there may be a vulnerable patient population who are at risk for persistent cognitive dysfunction following a TBI. Further understanding of the molecular mechanisms that contribute to this delayed recovery will aid in the development of therapeutics to mitigate the effects of ELS on TBI outcome.
Footnotes
Acknowledgments
The authors thank Akhila Sangadi for critical reading of the manuscript.
Funding Information
This work was supported by the Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development, Rehabilitation Research and Development Service (I21RX003017), The Miami Project to Cure Paralysis, and the Buoniconti Fund.
Author Disclosure Statement
No competing financial interests exist.