
Abstract
Human-induced pluripotent stem cell-derived neural stem/progenitor cell (hiPSC-NS/PC) transplantation during the acute phase of spinal cord injury (SCI) is not effective due to the inflammatory response occurring immediately after SCI, which negatively impacts transplanted cell survival. Therefore, we chose to study the powerful chemoattractant complement C5a as a method to generate a more favorable transplantation environment. We hypothesized that suppression of the inflammatory response immediately after SCI by C5a receptor antagonist (C5aRA) would improve the efficacy of hiPSC-NS/PCs transplantation for acute phase SCI. Here, we evaluated the influence of C5aRA on the inflammatory reaction during the acute phase after SCI, and observed significant reductions in several inflammatory cytokines, macrophages, neutrophils, and apoptotic markers. Next, we divided the SCI mice into four groups: 1) phosphate-buffered saline (PBS) only; 2) C5aRA only; 3) PBS + transplantation (PBS+TP); and 4) C5aRA + transplantation (C5aRA+TP). Immediately after SCI, C5aRA or PBS was injected once a day for 4 consecutive days, followed by hiPSC-NS/PC transplantation or PBS into the lesion epicenter on Day 4. The C5aRA+TP group had better functional improvement compared with the PBS only group. The C5aRA+TP group also had a significantly higher cell survival rate compared with the PBS+TP group. This study demonstrates that administration of C5aRA can suppress the inflammatory response during the acute phase of SCI, while improving the survival rate of transplanted hiPSC-NS/PCs, as well as enhancing motor functional restoration. Human-induced pluripotent stem cell-derived neural stem/progenitor cell transplantation with C5aRA is a promising treatment during the acute injury phase for SCI patients.
Introduction
A proportion of individuals who sustain a spinal cord injury (SCI) will suffer from lifelong and severe motor and sensory dysfunction, due to the body's limited ability for neurologic recovery post-injury. 1 To enhance recovery, much attention has been focused on cell transplantation therapies for SCI, especially the use of human-induced pluripotent stem cell (hiPSC). Indeed, we and other groups have demonstrated the efficacy of hiPSC-derived neural stem/progenitor cell (hiPSC-NS/PC) transplantation for conferring functional recovery in SCI, 2 -4 and these advancements are expected to reach clinical application in the near future. 5
However, critical issues remain surrounding the timing of cell transplantation and optimization of the injury site milieu. Currently, the optimal timing for transplantation is the subacute phase of SCI, which is about 7-14 days after SCI in rodents, 6 -8 after neuroinflammation subsides. Since the inflammatory reaction does not allow transplanted cells to survive in the acute phase of injury, which was considered to be within a week after trauma, 7,9 researchers have been forced to wait until the subacute phase to transplant cells. 9,10 During the acute phase of injury, several neurotoxic cytokines are upregulated and inflammatory cells intrude into the lesion area. 11,12 As a consequence of this heightened inflammatory response, there is significant host cell death and a diminished ability for the spinal cord to recover, and thereby, functional improvement is limited even after cell transplantation. However, if this harmful inflammatory response were to be suppressed immediately after the injury, a favorable environment for the grafted cells could be created, and moreover, it may be possible to perform cell transplantation in the acute phase.
The complement system has a very important role in initiating secondary damage during the acute phase in SCI. The complement proteins activate various neurotoxic cytokines and inflammatory cells, and this complex inflammatory cascade leads to the exacerbation of neural damage. 13 –16 Thus, the inflammation process is principally triggered by the complements, and among these proteins, C5a has recently received attention as a target of anti-inflammatory treatment.
C5a is a small glycoprotein (74 amino acids, about 11kDa) and generated by the cleaving of complement C5. 15 C5a is an anaphylatoxin, which causes inflammatory cytokine activation and leukocyte infiltration through the C5a receptor (C5aR). 17 Consequently, C5aR antagonist (C5aRA) could be a target as a treatment for inflammatory reaction after SCI. Among C5aRA, PMX205 (hydrocinnamate-(OPdChaWR)) is often used in the field of central nervous system disease experiment because PMX205 is able to penetrate blood–brain barrier and blood–spinal cord barrier, 18,19 and it has been shown that PMX205 administration after SCI suppresses inflammatory cytokines and macrophage infiltration into the lesion, decreases secondary damage, and improves the recovery of locomotor function. 20 These studies indicate that administration of C5aRA can inhibit the inflammatory reaction during the acute phase of SCI and prepare the injured microenvironment for receiving transplanted NS/PCs.
The purpose of the current study is to investigate whether C5aRA improves the inflammatory environment and enables increased survival of grafted cells when transplanted during the acute phase after SCI. We also evaluated the impact of a combined therapy of hiPSC-NS/PCs transplantation and C5aRA on locomotor functional recovery.
This study is particularly important as it is the first study to our knowledge that demonstrates immunosuppressant enables efficient hiPSC-NS/PCs transplantation during acute phase after SCI by increasing survival of grafted cells.
Methods
Animals
Adult female SCID-Beige mice (8-10 weeks, 17-22 g), which are deficient in lymphocytes and natural killer cells, were provided by Charles River Laboratory. All animals were housed in a temperature- and humidity-controlled environment. All experimental procedures were approved by the ethics committee of Keio University (Assurance No. 13020) and were in accordance with the Guide for the Care and Use of Laboratory Animals (National Institutes of Health, Bethesda, MD). All the studies were performed in accordance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines. An ARRIVE guidelines checklist and time-line diagrams of the study is included in Supplementary Additional File S1 and Supplementary Figure S1. 21
Spinal cord injury
The mice were anesthetized with ketamine (60 mg/kg) and xylazine (10 mg/kg) intraperitoneally. Laminectomy was performed at the 10th thoracic spinal vertebra (T10), and the dorsal surface of the dura matter was exposed. Moderate (70 kdyn) bilateral contusion injury was induced at the level of T10 using an Infinite Horizon impactor (Precision Systems and Instrumentation, Fairfax Station, VA), as previously described. 22 The muscles were sutured and the skin was closed with wound clips. After spinal cord injury, ampicillin (12.5 mg/kg) was administrated subcutaneously.
C5a enzyme-linked immunosorbent assay
A total of 25 mice were used in this experiment; after spinal cord injury, 6 mm of spinal cord, centered at the lesion epicenter, were dissected, as mentioned previously. 23 Detailed methods are described in the Supplementary Material Supporting Information.
C5a receptor Western blotting
A total of 16 mice were used in this experiment; after spinal cord injury, 6 mm of spinal cord, centered at the lesion epicenter, were dissected, as mentioned previously. 23 Detailed methods are described in Supplementary Material Supporting Information.
C5a receptor antagonist administration
We used PMX205 (Tocris Bioscience, U.K.) as a C5a receptor antagonist, as previously reported. 20 Immediately after SCI, the mice randomly received PMX205 (1 mg/kg body weight) 20 diluted in 1% ethanol or PBS containing 1% ethanol intraperitoneally once a day for 4 consecutive days or until sacrificed.
Quantitative reverse transcription polymerase chain reaction
A total of 50 mice were used in this experiment; the mice randomly received PMX205 or PBS intraperitoneally immediately after SCI once a day until sacrifice, and the injured mice were sacrificed at four time-points with naive controls. The times included 6 h, 12 h, 1 day, and 4 days after SCI. Detailed methods were described in the Supplementary Material Supporting Information.
mRNA-Seq
A total of 22 mice were used in this experiment; the mice randomly received PMX205 or PBS intraperitoneally immediately after SCI once a day until sacrifice, and the injured mice were sacrificed at 4 time-points with naive controls. The times included 3 h, 6 h, 12 h, 1 day, and 4 days after SCI. Detailed methods were described in the Supplementary Material Supporting Information.
Flow cytometry
A total of 12 mice were used in this experiment; the mice randomly received PMX205 or PBS intraperitoneally immediately after SCI once a day until sacrifice, and the injured mice were sacrificed at 4 days after SCI. Detailed methods are described in the Supplementary Material Supporting Information.
Cell culture, lentivirus transduction, neural differentiation, and immunohistochemistry
Cell culture of hiPSC (414C2) was performed as described previously, with subtle modifications. 24 Neural differentiation and immunohistochemistry were performed as described previously. 25 Detailed methods are described in the Supplementary Material Supporting Information.
Cell transplantation
Immediately after SCI, the mice were randomly divided into C5aRA administration or PBS administration intraperitoneally once a day for 4 consecutive days. Four days after SCI, the mice were randomly divided into two groups depending on whether or not hiPSC-derived NS/PCs transplantation was performed in each of the C5aRA-administered group, and the PBS-administered group based on their BMS score to ensure equivalent deficits across the groups. In brief, all mice were divided into four groups (PBS only group; C5aRA only group; PBS + transplantation (PBS+TP) group; and C5aRA + transplantation [C5aRA+TP] group), and re-anesthetized with isoflurane. Human-induced pluripotent stem cell–derived NS/PCs (5 × 105 cells/2 μL) or PBS were transplanted into the lesion epicenter using a metal needle at a rate of 1 μL/min. After transplantation, the skin was closed with wound clips and ampicillin (12.5 mg/kg) was injected subcutaneously. The mice were sacrificed 42 days after the SCI.
Bioluminescence imaging
Bioluminescence imaging (BLI) was performed as described previously. 26 Detailed methods were described in the Supplementary Material Supporting Information.
Assessment of grafted cell death
A total of 16 mice were used in this experiment; the C5aRA or PBS were administrated immediately after SCI for 4 days as mentioned above. Four days after SCI, hiPSC-derived NS/PCs were transplanted into lesion center. The mice were sacrificed at 3 and 7 days after transplantation, and immunohistochemistry was performed as described below.
Tissue immunohistochemistry
Tissue immunohistochemistry was performed as described previously. 27 Detailed methods are described in the Supplementary Material Supporting Information. Briefly, the following primary antibodies were used: anti-CD88 (rat, 1:400; Biorad, MCA2456); anti-C5R1 (rabbit, 1:1000; Abcam, ab59390); anti-Iba1 (rabbit, 1:400; WAKO, CDP0133); anti-LY6G (rat, 1:1000; Novusbio, NBP200441); anti-cleaved caspase-3 (rabbit immunoglobulin G [IgG], 1:500; Cell Signaling, 9661); anti–green fluorescent protein (GFP; rabbit, 1:2000; MBL); anti-human nuclear antigen (mouse IgG, 1:100; Millipore, MAB4383); anti-human nestin (rabbit, 1:200; IBL: 18741); anti-Ki67 (rabbit IgG, 1:200; Leica, NCL-Ki67p); anti-pan-ELAVL (human IgG, 1:1000, a gift from Dr. Robert Darnell; Rockefeller University, New York) 28 ; anti-APC (mouse IgG2b, 1:200; Millipore, OP80); anti-glial fibrillary acidic protein (GFAP; rabbit, 1:2000; Proteintech, AG10423); anti-STEM121 (mouse IgG1, 1:200, Takara Bio, Y40410); and anti-Syn1 (rabbit, 1:100, Abcam ab1543). Anti-CD88 and anti-C5R1 antibodies were used as a C5a receptor detection. Hoechst 33258 was used as a nuclear counter-staining. All images were obtained using a fluorescence microscope or a confocal laser scanning microscope.
For evaluation of C5aR expressing cells after SCI, sagittal sections were used and the regions within the area 2 mm caudal to the lesion epicenter were selected and counted under × 63 magnification. For quantification of survival rate of grafted cells, anti-GFP and Hoechst 33258 as a primary antibody and the appropriate secondary antibodies were used. The images were captured at the lesion epicenter and 0.2, 0.4, 0.6, 0.8, 1.0, 1.2, 1.4, 1.6, 1.8, and 2.0 mm rostral and caudal to the epicenter in axial sections using a fluorescence microscope, and the area of survived grafted cells was assessed by measuring the GFP positive areas on each cross section using ImageJ. For quantification of cleaved caspase-3-, APC-, GFAP-, pan-ELAVL-, Ki67-, nestin - and OCT4-positive cells, sagittal sections were used and five regions within the area 2 mm rostral and caudal to the lesion epicenter where human nuclear antigen (HNA)-positive grafted cells existed were selected and counted under × 63 magnification.
Immuno-electron microscopy
Immuno-electron microscopic analysis was performed as described previously. 29 Detailed methods are described in the Supplementary Material Supporting Information.
Locomotor function analysis
A total of 70 mice were used in this experiment; all behavioral assessments were performed and analyzed by one independent examiner blinded to the experimental groups. The hindlimb locomotor function analyses were evaluated using the Basso Mouse Scale (BMS) score, Rotor-rod and treadmill gait analysis. Detailed methods were described in the Supplementary Material Supporting Information.
Statistical analysis
All data are presented as means ± standard error of the mean. A Mann-Whitney U test was used to identify any significant differences between groups with respect to the results of flowcytometry and immunohistochemistry. One-way analyses of variance (ANOVA) followed by Tukey-Kramer tests for multiple comparisons were used to detect significant differences in stride length, stance angle, and rotarod score between the four groups. Two-way repeated-measures ANOVA followed by Tukey-Kramer tests were used for the others. For all statistical analyses, the significance level was set at p < 0.05. Microsoft Excel 2016 and IBM SPSS Statistics (ver. 25) were used for all calculations.
Results
C5a and C5aR expression in the spinal cord after injury
To investigate expression of C5a in the spinal cord, we evaluated the protein levels of C5a before and after injury at different time-points. The results revealed that C5a protein levels were significantly increased 1 day after injury (8.16 ± 0.83 pg/μg protein) and maintained the high amount of protein compared with the one prior to injury (2.43 ± 0.45 pg/μg protein) at 4 (6.69 ± 0.99 pg/μg protein), 7 (9.72 ± 0.85 pg/μg protein) and 14 days (8.24 ± 0.98 pg/μg protein; Fig. 1A).

Determination of C5a production and C5a receptor expression after SCI
Next, we investigated the level and distribution of C5a receptor expression in the injured spinal cord using Western blotting and immunohistochemistry. Western blotting showed that the level of C5a receptor expression were upregulated at 1 day after injury, then decreased at Days 4 and 7 (1 day: 14.02 ± 01.54; 4 days: 4.18 ± 0.85; 7 days: 2.08 ± 0.16) (Fig. 1B, 1C). To examine what types of cells expressed the C5a receptor, immunohistochemical analysis was performed for the injured spinal cord at 1 and 4 days after SCI. The results revealed that Iba1+ macrophages, activated microglia and some LY6G+ neutrophils expressed C5a receptor at 1 and 4 days after SCI (Fig. 1D, 1E). At 4 days after SCI, some pan-ELAVL+ neuron expressed C5a receptor, but APC+ oligodendrocyte and GFAP+ astrocyte rarely expressed C5a receptor. These results suggest that C5a protein was produced immediately after SCI, and that C5a receptors were expressed in inflammatory cells such as macrophages, activated microglia and neutrophils and neurons during the acute phase after SCI.
To examine what types of cells expressed the C5a receptor, immunohistochemical analysis was performed for the injured spinal cord at 1 and 4 days after SCI. As a result, Iba1+ macrophages and activated microglia and some LY6G+ neutrophils expressed C5a receptor at 1 and 4 days after SCI (Fig. 1D, 1E). At 4 days after SCI, some pan-ELAVL+ neuron expressed C5a receptor, but there were no detectable APC+ oligodendrocytes or GFAP+ astrocytes that expressed C5aR. These results suggest that C5a protein was expressed immediately after SCI, and that C5a receptors were expressed in inflammatory cells such as macrophages, activated microglia, as well as neutrophils and neurons during the acute phase after SCI.
Administration of a C5aR antagonist reduced the inflammatory response after SCI
To investigate the influence of C5aRA on the production of inflammatory cytokines in spinal cord tissue after SCI, C5aRA (PMX205, 1 mg/kg body weight) were injected into the mice immediately after SCI and the expression of cytokines in spinal cord tissue was evaluated using quantitative reverse transcription polymerase chain reaction. IL-1β and IL-6 were significantly reduced by the antagonist administration compared with PBS administration at 6 h (IL-1β: 60.9 ± 11.5 vs. 30.5 ± 5.2; IL-6: 1228.8 ± 380.1 vs. 112.8 ± 25.0; p < 0.05) and 12 h (IL-1β: 108.5 ± 23.1 vs. 36.1 ± 2.4; IL-6: 433.3 ± 102.7 vs. 177.2 ± 3.6; p < 0.05; Fig 2A, 2B). TNFα significantly reduced at 3 h (32.6 ± 6.9 vs. 10.6 ± 2.2; p < 0.05) and 6 h (21.5 ± 4.1 vs. 5.5 ± 1.2; p < 0.05) after SCI (Fig. 2C). Next, mRNA-Seq analysis was performed to analyze the influence of C5aRA on the gene expression profile in the injured spinal cord. mRNA sequencing revealed that the expression of genes associated with inflammatory cytokines was suppressed by administering C5aRA at 12 h after SCI (Fig. 2D). With regard to the apoptotic and necroptotic markers, administration of C5aRA also downregulated several apoptotic (caspase-8 and PIDD1) and necroptotic (RIPK3 and MLKL) markers 4 days after injury (Fig. 2E).
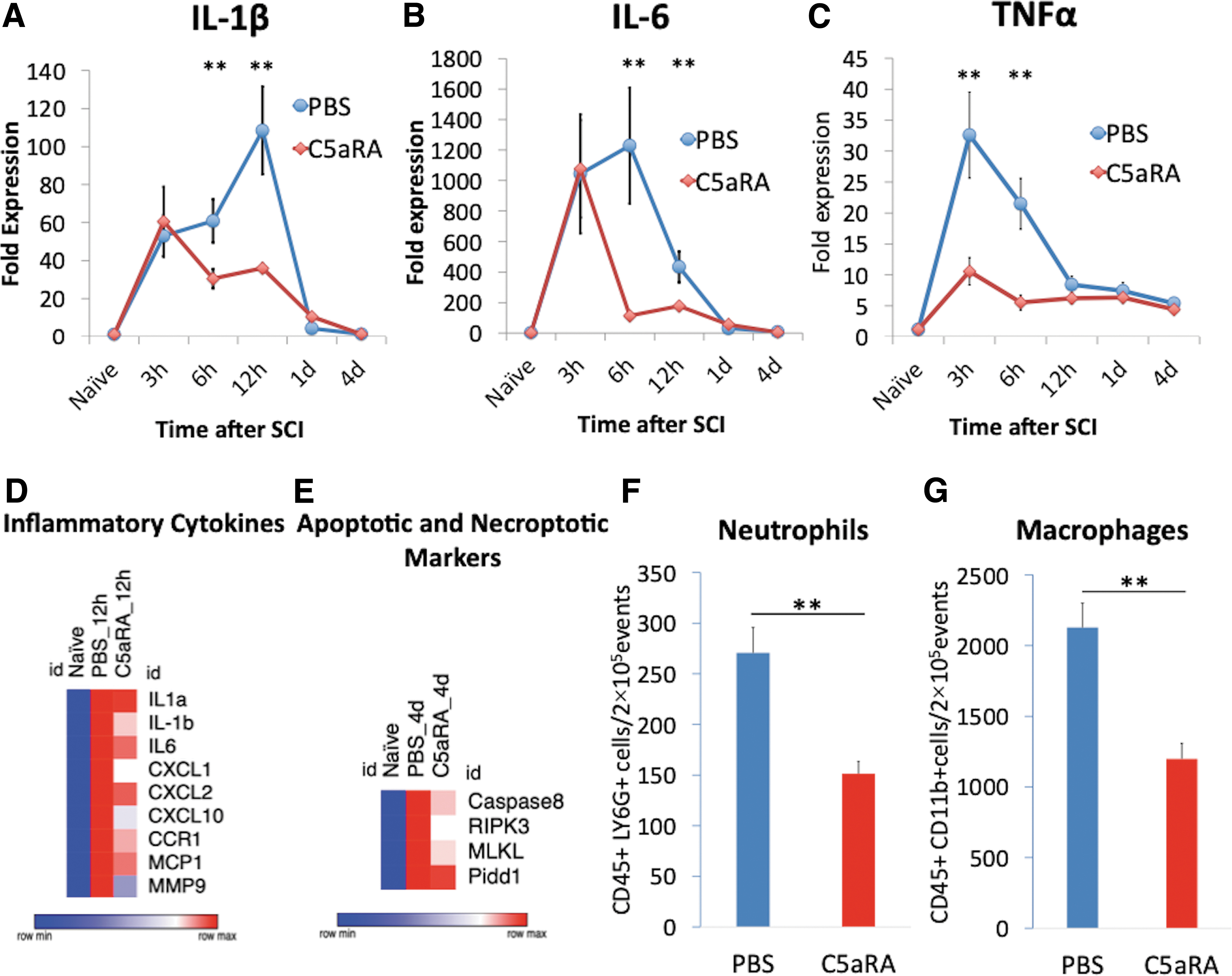
Inflammatory reaction after spinal cord injury with C5aR antagonist.
To investigate whether C5aR inhibition blocked infiltration of neutrophils and macrophages after SCI, flow cytometric analysis was performed. The results showed that administration of C5aRA reduced the number of CD45+/LY6G+ neutrophils and CD45+/CD11bhigh macrophages in the spinal cord compared with administration of PBS (neutrophils: 151 ± 12 cells/2 × 105 events vs. 270 ± 25 cells/2 × 105 events; macrophage: 1198 ± 111 cells/2 × 105 events vs. 2128 ± 173 cells/2 × 105 events; p < 0.05; Fig. 2F, 2G). These findings indicate that the acute inflammatory response after SCI occurs through C5a receptor activation, and inhibition of the C5a-C5aR axis suppressed inflammatory cytokine production and infiltration of inflammatory cells.
C5aR antagonist injection does not affect cell differentiation in vitro
To investigate the influence of C5aRA on hiPSC-NS/PCs, we evaluated the differentiation profiles and expression of the C5a receptor in these cells. The hiPSC-NS/PCs with or without C5aRA presented similar differentiation patterns; nestin+ undifferentiated cells (56.58 ± 4.07 % vs. 53.47 ± 3.06%; p = 0.513), β-III tubulin+ neurons (49.38 ± 3.87% vs. 44.87 ± 5.14%; p = 0.275), GFAP+ astrocytes (1.01 ± 1.01% vs. 2.53 ± 1.29%; p = 0.246), and O4+ oligodendrocytes (0 ± 0% vs. 0 ± 0%; p = 1.00; Fig. 3A-C), and there were no significant differences in all cell types between the groups (Fig. 3C). Some β-III tubulin+ cells and most of GFAP+ cells expressed nestin. In addition, the hiPSC-NS/PCs rarely expressed C5a receptor (Fig. 3D). These findings suggest that C5aR antagonist did not change the differentiation properties of hiPSC-NS/PCs.

hiPSC-NS/PCs differentiation and proliferation in vitro.
C5aR antagonist suppressed the apoptosis of grafted cells
To examine the effect of C5aRA on the apoptosis of grafted cells, we evaluated the expression of cleaved caspase-3, which is an apoptotic marker. As shown in Figure 4A and Supplementary Figure S2, cleaved caspase 3-positive grafted cells were detected in immunohistochemical analyses. Quantitative analyses revealed that the administration of C5aRA significantly reduced the cleaved caspase 3-positive grafted cells at both 3 and 7 days after SCI compared with PBS (Day 3: 9.91 ± 1.38% vs. 5.47 ± 1.05%; Day 7: 5.31 ± 0.32% vs. 1.72 ± 0.60%; p < 0.05; Fig. 4B). This result indicates that the administration of C5aRA significantly reduced the apoptosis of transplanted cells.
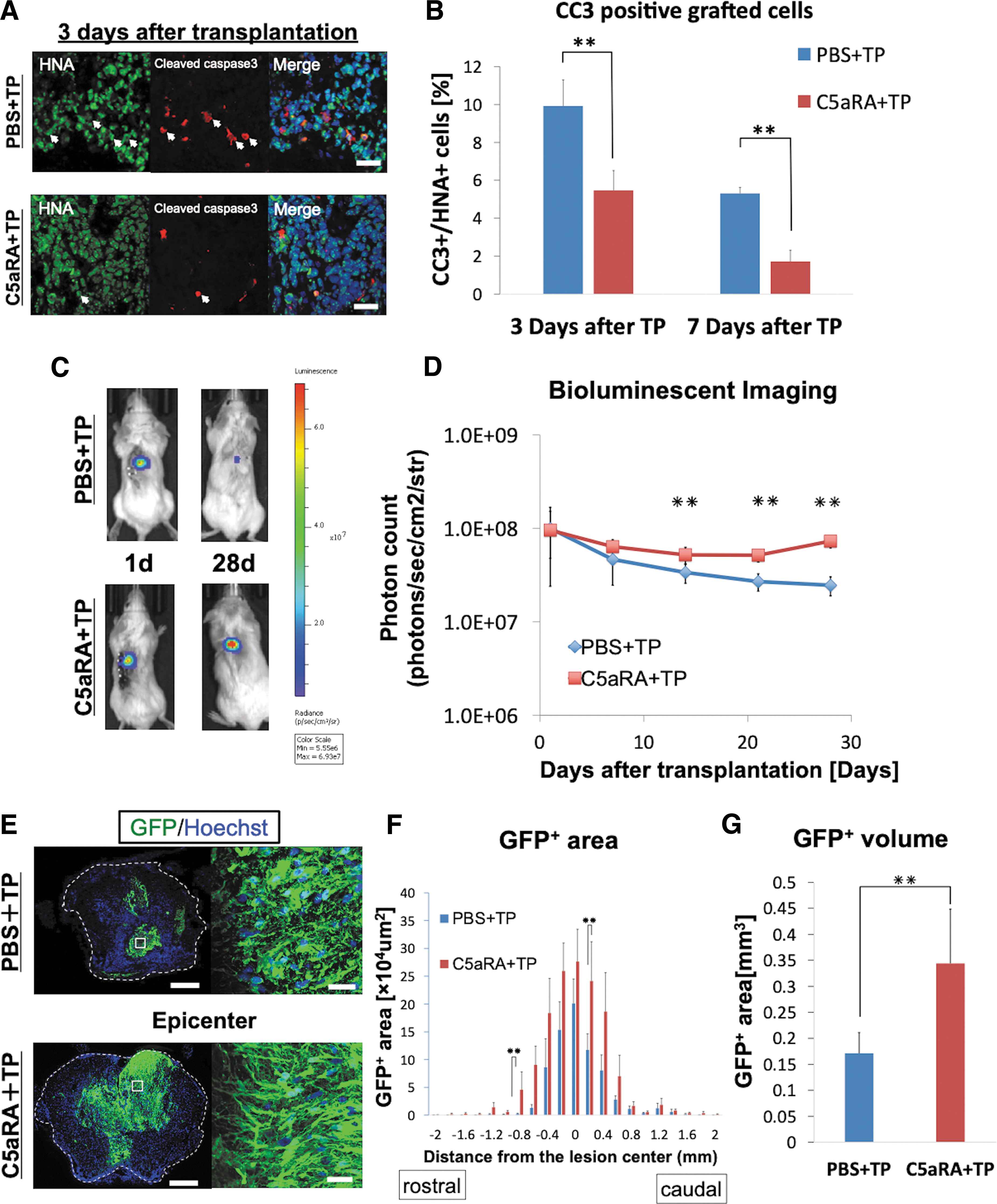
Apoptosis and survival rate of the transplanted cells.
Inhibition of C5aR increases the survival rate of grafted NS/PCs
To examine the effect of C5aRA on the survival rate of grafted cells, we assessed the luminescence of the grafted cells using BLI (Fig. 4C). The BLI analyses demonstrated that the luminescence of the grafted cells in the C5aRA+TP group was significantly higher than that of grafted cells in the PBS +TP group at 21 days post-SCI and thereafter (C5aRA+TP vs. PBS+TP, Day 21: 5.17 ± 1.13 E + 07 vs. 2.71 ± 0.57 E + 07; Day 28: 7.31 ± 1.12 E + 07 vs. 2.47 ± 0.53 E + 07; p < 0.05; Fig. 4D). We also performed the BLI analysis until 2 months after transplantation to observe for a longer period of time. This BLI analysis demonstrated that the luminescence of the grafted cells in C5aRA+TP group continue to be flat from 7 days to 63 days after transplantation (Supplementary Fig. S3).
We also evaluated grafted cell survival using immunohistochemistry (Fig. 4E; Supplementary Fig. S4). In the C5aRA+TP group, GFP+ area had a tendency to increase compared with PBS+TP group in each axial section (Fig. 4F), and total volume of GFP+ areas was significantly larger compared with the PBS+TP group (C5aRA: 0.34 ± 0.10 mm3; PBS: 0.17 ± 0.04 mm3; p < 0.05; Fig. 4G). Moreover, there was no visible tumor-like tissue observed in any of the hematoxylin and eosin–stained images in the both groups (Supplementary Fig. S5). These results indicate that inhibition of C5aR improves the survival and proliferation of hiPSC-derived NS/PCs after the transplantation.
Administration of C5aR antagonist does not affect cell differentiation in vivo
In order to evaluate the neural differentiation and proliferation of the grafted cells in vivo, we performed immunohistochemical analyses using antibodies specific for HNA and cell type specific markers. Immunohistochemistry revealed that the HNA positive grafted cells in the C5aRA+TP and PBS+TP groups differentiated into pan-ELAVL+ neurons (37.0 ± 3.6% and 36.8 ± 3.9%; p = 0.827), GFAP+ astrocytes (46.2 ± 6.0% and 33.1 ± 3.2%; p = 0.127), and APC+ oligodendrocytes (8.0 ± 1.2% and 11.6 ± 1.5%; p = 0.127; Fig. 5A, 5B). The differentiation rates of neurons, astrocytes, and oligodendrocytes did not significantly differ between the C5aRA+TP and PBS+TP groups (Fig. 5C). The proportions of Ki67+ cells (2.5 ± 0.7% vs. 2.7 ± 0.8%; p = 0.43) and OCT3/4+ cells (0 ± 0% vs. 0 ± 0%; p = 1.0) were not significantly different between the two groups (Fig. 5C).

Transplanted hiPS-NS/PCs differentiation and proliferation.
Next, we examined synapse formation between host tissue and neurons derived from grafted cells in the C5aRA+TP group to demonstrate neuronal relay between host and grafted cells. Immunohistochemistry showed that some STEM121 positive grafted cells expressed Synapsin1, which is a synapse marker (Fig. 6A). Immunoelectron microscopy revealed that pre- and post-synaptic structures labeled with STEM121 were detected (Fig. 6B). These results indicate that host-to-graft and graft-to-host neuronal connections were formed in the C5aRA+TP group.
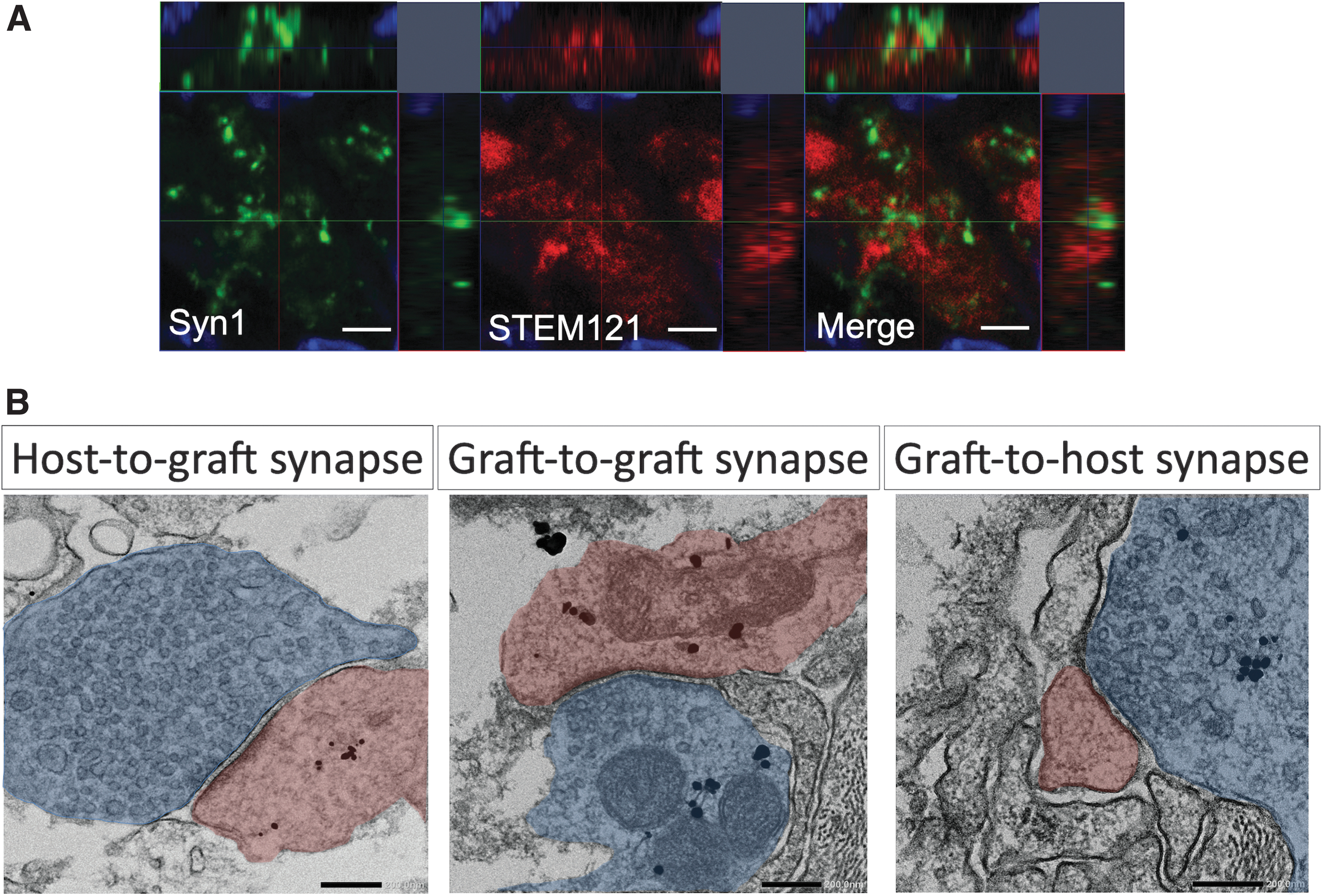
Histological synapse formation in C5aRA+TP group.
C5aR inhibition at the acute phase improved locomotor function
We assessed hindlimb locomotor function using the BMS score, Rotor-rod test, and treadmill gait analysis with the Digigait system. The average BMS scores of the C5aRA+TP group were significantly higher than those of the PBS group at Day 28 (3.5 ± 0.3 vs. 2.4 ± 0.2; p < 0.05) and Day 35 (3.8 ± 0.3 vs. 2.4 ± 0.2; p < 0.05) after SCI (Fig. 7A). In the Rotor-rod test, which was evaluated at Day 35, the running time of mice in the C5aRA+TP group was significantly longer than the time in the other groups (C5aRA+TP:17.5 ± 4.0sec; PBS+TP: 9.7 ± 1.3 sec; C5aRA: 9.5 ± 1.2 sec; PBS: 7.5 ± 0.8 sec; p < 0.05; Fig. 7B). Using the treadmill Digigait system to analyze gait performance, only the mice in the C5aRA+TP groups showed significant improvement in stride length (C5aRA+TP: 4.17 ± 0.16 cm; PBS+TP: 3.74 ± 0.16 cm; C5aRA: 3.65 ± 0.08 cm; PBS: 3.43 ± 0.13 cm; p < 0.05) or paw angle (C5aRA+TP: 23.5 ± 3.3°; PBS+TP: 32.8 ± 3.8°; C5aRA: 29.1 ± 3.7°; PBS: 41.9 ± 4.5°) compared with the PBS group (p < 0.05; Fig. 7C, 7D)
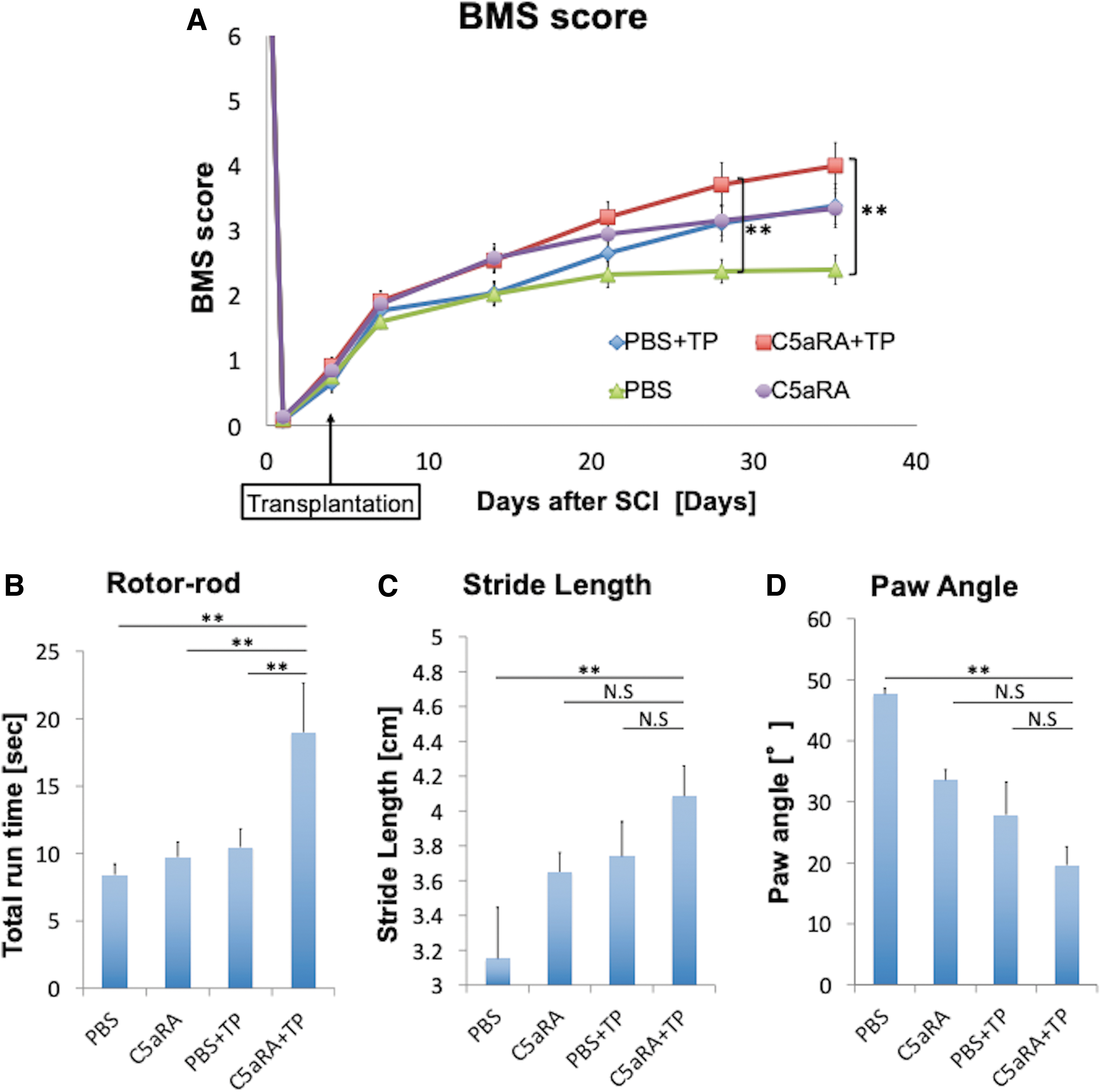
Hindlimb motor function after transplantation.
Discussion
As shown previously, anti-inflammatory treatment is needed to enable efficient cell transplantation therapy during acute phase because transplanted cell rarely survives in an inflammatory environment in spinal cord during acute phase after SCI, but effective immunosuppressant that increase grafted cell survival has not been found. Therefore, we hypothesized that C5aRA could be a novel immunosuppressant which strongly suppressed inflammatory reaction after SCI and enables efficient hiPSC-NS/PCs transplantation during acute phase. The present study demonstrates that C5aRA administration after SCI significantly reduces upregulation of IL-1β, IL-6, and TNFα, as well as the infiltration of neutrophils and macrophages. C5aRA also downregulated the expression of inflammatory cytokines and apoptosis markers, which was demonstrated by RNA sequence analysis. The combined therapy of hiPSC-NS/PCs transplantation and C5aRA decreased the grafted cell death including apoptosis compared with cell transplantation monotherapy. Consequently, this combined therapy significantly improved locomotor function after SCI. Our findings show that C5aRA could be a promising medication to enhance the efficacy of hiPSC-NS/PCs when transplanted during the acute phase of SCI.
It is well known that inflammatory reactions play critical roles in the exacerbation of secondary damage in the SCI microenvironment. 6,9,11,13 For example, IL-1β and TNFα contribute to upregulation of several inflammatory mediators, recruiting neutrophils and macrophages, and resulting in the apoptosis of neurons and oligodendrocytes after SCI. 30 –34 IL-6 also promotes cytotoxic macrophage infiltration into the lesion and increases secondary damage. 35,36 Neutrophil and macrophage infiltration into the lesion area is a detrimental factor for resident cell survival and functional recovery after SCI. 37 –40 C5aR expressed on microglia/macrophages and neutrophils, and the C5a-C5aR axis activates these cells and inflammatory cytokines such as IL-1, IL-6, and TNFα. As reported in the previous study, 20 we demonstrated that administration of C5aRA reduced secretion of various cytokines, infiltration of inflammatory cells, and apoptosis of residual cells in the spinal cord. Further, we presented that some neurons expressed C5aR during the acute phase after SCI. A previous study has demonstrated that C5a-bound C5aR directly induced neuronal apoptosis, 41 indicating that the administration of C5aRA directly inhibited apoptosis in the present study. Thus, an intervention using C5aRA could exert stronger effects than each individual anti-inflammatory drug on suppressing multiple inflammatory mediators, preventing the expansion of secondary damage, and creating a hospitable environment for cell survival after transplantation.
Previous reports have revealed that the acute inflammatory reaction after central nervous system (CNS) injury contributes to rejection of grafted cells in spinal cord tissue. 42 -44 To overcome this situation, several studies have tried to use immunosuppressant drugs when transplanting cells after SCI. 40,45 -47 However, there are few reports that showed successful improvements in cell survival. For example, depletion of neutrophils was found to decrease the astrogliosis of the transplanted cells, but did not change the survival rate. 47 A reduction in macrophages also failed to favorably affect cell engraftment when the drug clodronate was administrated. 40
It is inferred from those results that the blockade of a single inflammatory factor cannot decrease grafted cell death. Therefore, a strong immunosuppressant capable of blocking several inflammatory reactions after SCI is indispensable for the improvement of cell survival. C5aRA was an efficacious drug against rejection of hiPSC-NS/PCs transplantation because this medication was able to suppress multiple inflammatory cytokines and cells, and our results demonstrated the reduction of apoptosis of the grafted cells and amelioration of the cell survival rate. We can deduce from this result that C5aRA is more effective for improving the efficacy of cell transplantation into the injured spinal cord than other immunosuppressants, because the C5a-C5aR axis regulates several inflammatory reactions in the acute phase after SCI. 15
We demonstrated that the differentiation rate of astrocytes in the C5aRA group was relatively lower than the PBS group, yet there was no statistically significant difference (Fig. 5C). Astrogliosis is known to be promoted by some inflammatory cells and cytokines, such as polymorphonuclear cells, TNFα and IL-6. 48 However, previous reports evaluated the role of C5a for astrogliosis after SCI, and demonstrated that administration of C5aRA in the acute phase of injury did not reduce astrogliosis, whereas inhibition of the C5a-C5a receptor axis in the chronic phase suppressed astrogliosis. 20 These results indicate that C5a inhibition in the acute phase did not have a specific impact on astrogliosis. Our study also demonstrated that administration of C5aRA in the acute phase after SCI did not influence the astroglial differentiation of transplanted cells, and this result could be consistent with findings from the previous study.
Our results demonstrated that the intensive inflammatory reaction during the acute phase of SCI did not allow transplanted cells to survive sufficiently and exert their efficacy when C5aRA was not used. Although the in vivo differentiation profiles were comparable between groups with or without the C5aRA (Fig. 5) and the changes to cell survival rate were small between these groups, C5aRA administration did enhance locomotor functional recovery. Our results indicate that the difference in the number of survived transplanted cells is important to result in a significant functional improvement in this transplantation therapy. By using C5aRA, it is inferred that more NS/PCs differentiated into axons to construct functional circuits, or differentiated into oligodendrocytes for remyelination, and these mechanisms led to functional recovery. 2,49,50
Further, our results demonstrated that host-to-graft and graft-to-host neuronal connections were histologically formed in the C5aRA+TP group. Kawai and colleagues demonstrated that transplanted hiPSC-derived NS/PCs following a contusive spinal cord injury formed histologically confirmed synaptic connections with host tissue by immune-electron microscopy, and that these synaptic connections physiologically function as neuronal relays and contribute to functional recovery. 51 Similarly to this article, neurons differentiated from grafted cells physiologically connected with host neurons and contributed to functional recovery in our study. Therefore, we propose that improving the survival of grafted cells leads to an increase in neuronal connections between grafted cells and host tissues, resulted in improved motor function.
If cell transplantation is possible in the acute phase of injury, earlier interventions could become feasible and bring the benefits of regenerative medicine to the SCI field.
For the transplantation studies using human-derived cells, immunodeficient animals were employed to avoid rejection of the grafted cells. For example, NOD-SCID mice which generally used in transplantation experiment lacked lymphocytes and impaired innate immune system such as neutrophils, macrophage and complement. 26,50,52 -54 However, NOD-SCID mice are not suitable for our current study because normal activation of the complement system is necessary to evaluate the effect of C5aRA. Immunocompetent mice are also not appropriate for the present study owing to lymphocytes, which strongly reject xenograft. 44,55 In this study, we selected SCID-Beige mice, which lack lymphocytes but maintain a functional complement system. We demonstrated that the complement system was activated normally after SCI in SCID-Beige mice, and we could perform evaluating the influence of C5aRA on the grafted cells and locomotor function recovery. However, as it is known that lymphocytes express C5a receptor and regulate proliferation and survival of lymphocytes, 56 this study using SCID-Beige mice might not be sufficient to accurately evaluate the effect of C5aRA against cell transplantation after SCI. Therefore, further studies using immunocompetent mice with lymphocyte suppressants are necessary to enhance our understanding.
The complement cascade has an important role in the innate immune system. Particularly, C5a, which is a strong anaphylatoxin, has been attracting attention as a target for drug treatment. Against C5a-C5aR axis, several C5aR antagonists have been developed. Among them, PMX53 has been widely used and provides beneficial effects for several diseases such as arthritis, sepsis, and inflammatory bowel diseases. 57 -59 PMX205 was developed by modifying the extra cyclic phenylalanine residue of PMX53. Compared with PMX53, PMX205 was more efficient in penetrating and infiltrating the brain and spinal cord. This drug has shown efficacy in CNS diseases, including amyotrophic lateral sclerosis, Alzheimer's disease and SCI. 20,25,60 In addition, PMX205 is a likely clinical candidate as it is currently being examined as part of an ongoing Phase1 trial for Alzheimer's disease (Australian Clinical Trials: ACTRN12619001639112). Therefore, we used PMX 205 in this SCI study and demonstrated significant functional recovery. PMX205 represents a promising drug that enhances the efficacy of hiPSC transplantation therapy for SCI.
Conclusion
The present study demonstrates that the administration of C5aRA suppresses the inflammatory response during the acute phase of SCI. This beneficial effect led to improved transplanted hiPSC-NS/PCs survival rates as well as the enhancement of motor functional restoration. These findings suggest that administration of C5aRA makes it possible to transplant hiPSC-NS/PCs during the acute phase of SCI. This work opens a potentially novel strategy wherein the transplantation of neural stem cells could be combined with early decompressive surgery as a one-stage treatment for severe SCI.
Footnotes
Acknowledgments
We appreciate the assistance of O. Tsuji, M. Shinozaki, T. Kondo, T. Iida, T. Okubo, S. Ito, K. Kojima, Y. Tanimoto, Y. Hoshino, Y. Kamata, K. Kajikawa, K. Ago, T. Kitagawa, and M. Kawai who are all members of the spinal cord research team in the Department of Orthopedic Surgery and Physiology, Keio University School of Medicine, Tokyo, Japan. We also thank K. Yasutake, T. Harada, and M. Akizawa for their assistance with the experiments and animal care. Further, we thank T. Worden for his assistance in proofreading.
Authors' Contributions
Reo Shibata: Conception and design, collection and assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript.
Narihito Nagoshi: Conception and design, financial support, administrative support, collection and assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript.
Keita Kajikawa: Collection of data of the animal behavior experiments, and data analysis and interpretation.
Shuhei Ito: Collection of data of the animal behavior experiments, and data analysis and interpretation.
Shinsuke Shibata: Collection of data of the immune-electron microscopic experiments, data analysis and interpretation, and reviewing and editing the manuscript.
Tomoko Shindo: Collection of data of the immune-electron microscopic experiments, and data analysis and interpretation.
Mohamad Khazaei: Conception and design, and reviewing and editing the manuscript.
Satoshi Nori: Reviewing and editing the manuscript.
Jun Kohyama: Collection and assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript.
Michael G. Fehlings: Conception and design, and reviewing and editing the manuscript.
Morio Matsumoto: Reviewing and editing the manuscript.
Masaya Nakamura: Reviewing and editing the manuscript.
Hideyuki Okano: Reviewing and editing the manuscript.
Funding Information
This study was supported by funding from the Research Project for Practical Applications of Regenerative Medicine from the Japan Agency for Medical Research and Development (AMED), Grant No. JP21bk0104114.; JSPS KAKENHI (Grant Number 17H04318 to N.N.), a medical research grant on traffic accidents from the General Insurance Association of Japan, and AO Spine Japan (AOSJP(R)2017-14).
Author Disclosure Statement
H.O. serves as a paid scientific advisor to San Bio Co. Ltd., and K Pharma Inc. M.N. serves as a paid scientific advisor to K Pharma Inc. For the other authors, no competing financial interests exist.
Supplementary Material
Supplementary Additional File S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supporting Information
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.