
Abstract
Traumatic spinal cord injury (SCI) is a devastating condition for which effective neuroregenerative and neuroreparative strategies are lacking. The post-traumatic disruption of the blood–spinal cord barrier (BSCB) as part of the neurovascular unit (NVU) is one major factor in the complex pathophysiology of SCI, which is associated with edema, inflammation, and cell death in the penumbra regions of the spinal cord adjacent to the lesion epicenter. Thus, the preservation of an intact NVU and vascular integrity to facilitate the regenerative capacity following SCI is a desirable therapeutic target. This study aims to identify a therapeutic window of opportunity for NVU repair after SCI by characterizing the timeframe of its post-traumatic disintegration and reintegration with implications for functional spinal cord recovery. Following thoracic clip-compression SCI or sham injury, adult C57BL/6J mice were followed up from one to 28 days. At one, three, seven, 14, and 28 days after SCI/sham, seven-Tesla magnetic resonance imaging (MRI), neurobehavioral analysis (Basso mouse scale, Tally subscore, CatWalk® gait analysis), and following sacrifice immunohistochemistry were performed, assessing vessel permeability via Evans blue (EVB) extravasation, (functional) vessel density, and NVU integrity. Thy1-yellow fluorescent protein+ mice were additionally implanted with a customized spinal window chamber and received longitudinal in vivo two-photon excitation imaging (2PM) with the injection of rhodamine-B-isothiocyanate-dextran for the combined imaging of axons and vasculature up to 14 days after SCI/sham injury. Post-traumatic edema formation as assessed by MRI volumetry peaked at one to three days after injury, while EVB permeability quantification revealed a thoroughly injured BSCB up to 14 days after SCI. Partial regeneration of functional vasculature via endogenous revascularization was detected after one to four weeks, however, with only 50–54% of existing vessels regaining functional perfusion. Longitudinal in vivo 2PM visualized the progressive degeneration of initially preserved spinal cord axons in the peri-traumatic zone after SCI while displaying a rarefication of functionally perfused vessels up to two weeks after injury. Neurobehavioral recovery started after one week but remained impaired over the whole observation period of four weeks after SCI. With this study, a therapeutic window to address the impaired NVU starting from the first days to two weeks after SCI is identified. A number of lines of evidence including in vivo 2PM, assessment of NVU integrity, and neurobehavioral assessments point to the critical nature of targeting the NVU to enhance axonal preservation and regeneration after SCI. Continuous multifactorial therapy applications targeting the integrity of the NVU over the identified therapeutic window of opportunity appears promising to ameliorate functional vessel perseverance and the spinal cord’s regenerative capacity.
Introduction
Traumatic spinal cord injury (SCI) is a devastating condition that is one of the leading causes of disability and a major socioeconomic burden for health care systems. 1 –4 Despite advances in medical, surgical, and rehabilitation treatments, effective neuroregenerative and neuroreparative strategies to treat SCI are lacking.
The post-traumatic disruption of the blood–spinal cord barrier (BSCB) is one major factor in the complex pathophysiology of SCI. 5 –7 Following SCI, a disruption of the BSCB occurs with the consequence of edema formation and tissue inflammation, leading to an ischemic environment aggravating and spreading the injury along to formerly non-injured spinal cord regions. 6 –8 One consequence of this ischemic environment is cell death in the neurovascular unit (NVU), consisting of endothelial cells, astrocytic foot processes, and pericytes, as well as tight and adherens junction proteins sealing the NVU. 6 –11 As a result, ischemic apoptosis and necrosis of other central nervous system cells occurs in and around the injury site. 10,12 –14
Previous studies dealing with post-traumatic BSCB disruption have discovered a time course of secondary disruption peaking at one hour to one week after injury, with subsequent, but partial, endogenous regeneration. 15 –18 However, some studies revealed significant BSCB disruption for a much longer period. 19 –22 As the post-traumatic disruption of the BSCB presents a disruptive event on the one hand, but also a chance for potential therapies to pass this physiological barrier on the other hand, a thorough understanding of this pathophysiology and its regenerative time course is key to identifying potential protective and regenerative therapies and the most promising timeframe for their application. 23 –25
Moreover, the regeneration of the NVU and thus of functionally perfused vessels seems essential for eliminating local hypoxia–ischemia and preserving tissue integrity and function, as was previously shown in SCI and stroke. 26 –30 In both entities, vascular regeneration is associated with neurogenesis and ameliorated neurological function, as functional vessels can provide trophic support for regenerating tissue. 31,32 To preserve an intact NVU and vessel function and to ameliorate the regenerative capacity following SCI is therefore a desirable therapeutic target to restore spinal cord integrity and function. 24,25
This study aims to identify a therapeutic window for applications targeting the pathologically hyperpermeable neurovascular unit after SCI. To this end, the spatial and temporal distribution of the disruption of the BSCB and the cellular parts of the NVU, as well as its endogenous regeneration after SCI, are characterized and paralleled with the recovery of neurological function over the acute into the chronic injury phase, using an established clip-compression injury model for murine mid-thoracic SCI. 23,33 –35
Materials and Methods
Animal experiments were approved by the local and governmental animal care committee (G0314/17) and the study is registered on the platform animaltestinfo.org (NTP-ID 00019011–1-7). Animals were randomly assigned to the sham or SCI group and to all respective subgroups. The experimental design is illustrated in Figure 1.
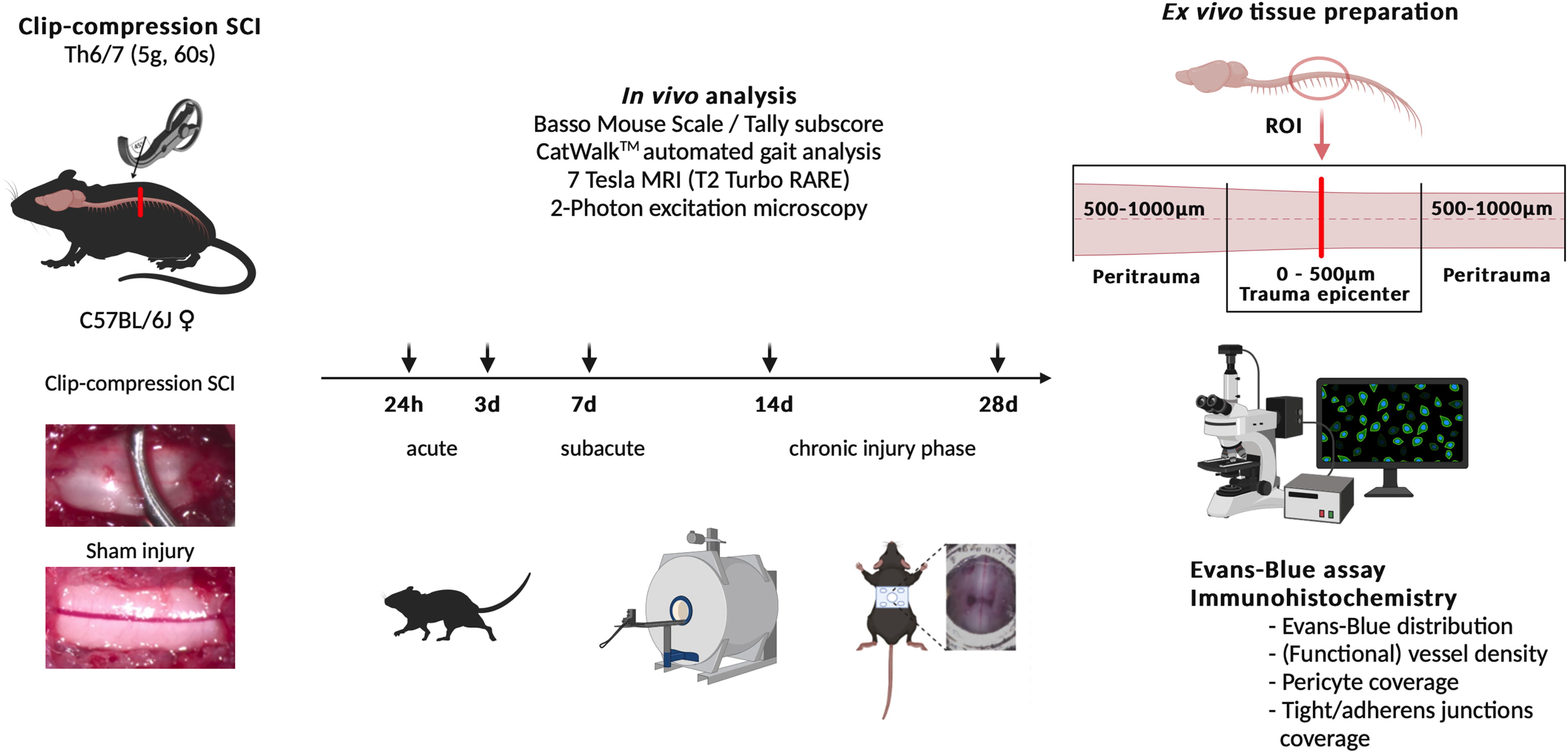
Illustration and overview of the experimental setup. MRI, magnetic resonance imaging; RARE, rapid acquisition with relaxation enhancement; ROI, region of interest; SCI, spinal cord injury; Th, thoracic vertebra.
Mid-thoracic clip-compression SCI
A total of 144 adult (age 12–14 weeks, 20–35 g body weight) C57BL6/J (wild-type) mice (Charles River, Germany) were placed under general anesthesia using injections of ketamine and xylazine (9 mg ketamine, 1 mg xylazine/100 g body weight). The incision area was shaved and disinfected. Under deep anesthesia, a midline incision in the mid-thoracic area was applied, followed by a microsurgical two-level laminectomy at Th6/7 with microscissors. The spinal cord space between Th6/7 was subsequently compressed for a duration of 60 sec using a 5.0-g modified aneurysm clip (FEJOTA™), 23,33 –35 generating a standardized moderately severe bilateral compression-contusion SCI (n = 88 total SCI animals/all groups). After cleaning the situs with saline and Serasept solution, the skin was closed in a layering fashion. In sham animals (n = 56 total sham animals/all groups), a two-level laminectomy was performed at Th6/7 without SCI application. Post-operatively, animals were housed in standard mouse cages, with injured and sham-injured animals housed separately. Clavamox antibiotic (amoxicillin/clavulanic acid) was applied via drinking water for 14 consecutive days peri-operatively (seven days pre- to seven days post-surgery, 25 mg/100 mL). Access to food and water was given ad libitum. Animal’s bladders were manually emptied every eight–12 h until recovery and body weight was measured daily. Post-operative pain medication was administered using buprenorphine injections (0.05 mg/kg body weight, intramuscularly) every eight hours for three days post-operatively (see Supplementary Fig. S1 for detailed animal care procedures, Supplementary Fig. S2 for exemplary intraoperative images, and Supplementary Table S1 for an overview of group sizes).
Basso mouse scale and tally subscore
Mice were placed in an open field and locomotion was assessed by a blinded observer using the established Basso mouse scale (BMS; 0–9 points). 36 In case of regained or preserved hindlimb function, the cumulative Tally subscore was applied (0–11 points). 37 Neurobehavioral analyses were performed pre-operatively (baseline) and at timepoints one, three, seven, 14, and 28 days post-operatively (n = 10/group).
CatWalk™ automated gait analysis
Gait was automatically analyzed using the CatWalk automated gait analysis system 38,39 (CatWalk TM, Noldus Information Technology, CatWalk XT Version 9.1) at timepoints one, three, seven, 14, and 28 days post-operatively (n = 10/group). Pre-operatively, three consecutive training runs/animal were performed to build an individual baseline. Successful performance required three complete runs/animal/timepoint. Incompletely assessed paw print data was manually labeled by an experienced observer in a blinded fashion and data cleaned in a standardized fashion following every run.
7-Tesla small animal magnetic resonance imaging
Magnetic resonance imaging (MRI) was performed before surgery (baseline) and at timepoints one, three, seven, 14, and 28 days post-operatively under volatile anesthesia (isoflurane) using a seven-Tesla (7T) small animal MRI scanner (7T PharmaScan 70/20USR, Bruker Corp.) with a modified rat brain surface coil (RapidBiomed). 40 Standardized T2-Turbo rapid acquisition with relaxation enhancement sequences were acquired in every animal using the software Paravision 6.0.1 (Bruker) following scout scans to center the acquisition around the lesion epicenter. Edema expansion was assessed using the OsiriX Lite software (OsiriX Lite 2022, Pixmeo) in sagittal MRI slices (16 sagittal slices/measurement, 300 µm slice thickness) and compared at every given timepoint to the animal’s non-injured baseline (n = 5). Regions of interest (ROIs) were defined as hyper-intense regions indicating edema in the spinal cord in each slice and selected with the free-hand tool in OsiriX Lite.
Spinal window chamber implantation
Following thoracic SCI or sham injury, a spinal window chamber adapted by Farrar et al. 41 fabricated from stainless steel was implanted over the surgically exposed spinal cord region in a total of 16 (n = 8 SCI, n = 8 sham) male heterozygotous Thy-yellow fluorescent protein (YFP)+ C57BL/6J transgenic mice 42 to image dorsal column axons and associated vasculature repeatedly in vivo, without the necessity for repetitive surgery. Due to higher body weight and increased robustness, male animals were used for this procedure. Pre- and post-operative animal care was carried out as described and mice were housed individually to prevent damaging of the implant (Supplementary Fig. S2).
Longitudinal in vivo microscopy
To image the spinal cord in vivo, mice were anesthetized with ketamine/xylazine and placed in a stereotaxic frame to secure the top plate of the implant horizontally. Mice were hereby slightly elevated to reduce breathing artifacts as described before. 41 Body temperature as measured by a rectal probe was maintained at 37°C during imaging by a heating pad and mice were allowed to recover on a heated surface after imaging. For vessel visualization, mice received intravenous injection of 100 µL rhodamine-B-isothiocyanate-dextran (RD) 70 kDa (Sigma) right before each imaging session. For two-photon excitation fluorescence microscopy (2PM), a custom-designed multiphoton microscope (LaVision Biotec GmbH, Bielefeld) was used combined with a tunable femtosecond pulsed Ti:Sa laser (wavelength range 700–1020 nm, 100 fs, 80 MHz, MaiTai, Spectra Physics, Germany), controlled by ImSpector software 208 (LaVision BioTec GmbH, Bielefeld, Germany). For high-resolution imaging, the microscope was equipped with a 20× water-immersion objective lens (numerical aperture = 0.95; Olympus, Hamburg, Germany). The laser was tuned to 920 nm for excitation of YFP, rhodamine-B-dextran, and second-harmonic generation (SHG) or to 850 nm for excitation of rhodamine-B-dextran individually. Emission filters were used at 525/50, 593/40, and 466/40 nm to isolate fluorescence from YFP, rhodamine-B-dextran, and SHG, respectively.
2PM image processing
For image processing and visualization, Imaris software (Bitplane AG, Zurich, Switzerland) was used. 2PM was used to acquire 3D images at the lesion epicenter and at the peri-traumatic lesion directly adjacent rostrally and caudally. Each image was collected as z-stack (1–4 µm/step size) in a depth of 150–250 µm. The acquisition time for an image with a field of view of 500 × 500 μm2 and a digital resolution of 505 × 505 pixel was 800 ms. Imaging depth was measured by identifying the dura as starting point via the SHG signal. The image data was converted using ImarisFileConverter 9.9.1 and rendered and visualized within ImarisViewer 9.9.1.
Post-mortem tissue processing
Phosphate-buffered saline perfusion for immunohistochemistry
Mice were perfused with 40 mL ice-cold phosphate-buffered saline (PBS). Brain and spinal cord were consecutively dissected and a 1 cm section of the spinal cord surrounding the injury site was defined as ROI and separated. Tissue was snap-frozen in nitrogen and stored at −80°C.
Evans blue perfusion for BSCB integrity analysis
For later quantification of Evans blue (EVB) extravasation, mice were injected with 100 µL EVB (64 kDa, 2%) intravenously before transcardial perfusion with 40 mL PBS as described above. EVB was allowed to circulate 20 min prior to sacrifice with the animal under deep anesthesia. ROI tissue (1 cm) was snap-frozen and processed for either fluorescence assay or histological processing.
Fluorescein isothiocyanate -lectin perfusion for functional vessel analysis
For the post-mortem quantification of functionally perfused vessels, 100 µL fluorescein isothiocyanate (FITC)-lectin in 150 µL PBS was infused intracardially 2 min prior to sacrifice via exsanguination. ROI tissue (1 cm) was snap-frozen and processed for histological analysis.
EVB assay for BSCB integrity
EVB extravasation was quantified in a total of 50 mice (n = 5/group/timepoint) using an automated fluorescence assay. 23,43 Following the above-described EVB perfusion, ROI tissue (1 cm) was weighed and homogenized via pulse-sonification in N,N-dimethylformamide (Merck), then centrifuged at 21.000 rpm/30 min (Sigma). After collection and aliquotation of the supernatant into a 96-well glass plate (Greiner Bio-One), fluorescence was measured between 620 and 680 nm (Tecan microplate reader). EVB concentration was calculated based on a standard curve and samples were normalized to tissue weight (µg/g).
Histochemistry and immunohistochemistry
For histological processing, ROI tissue (1 cm) was embedded in gelatine and serial cryosections were cut axially (10 µm slice thickness) in two peri-traumatic regions (epicenter: ±500 µm caudal-rostral, peri-trauma: ±500–1000 µm caudal-rostral) and mounted on Superfrost charged glass slides (Superfrost™ Plus Adhesion Microscope Slides). Single additional spinal cords (one per timepoint after SCI, one after sham) were additionally cut longitudinally (20 µm slice thickness) following EVB perfusion to visualize EVB extravasation around the trauma epicenter (centered medially).
Four serial axial slices/region/animal (two regions, epicenter: ±500 µm caudal-rostral, peri-trauma: ±500–1000 µm caudal-rostral, n = 4/group/timepoint in SCI, n = 4/sham, totaling in 24 spinal cords/staining group) were co-stained with the following goals: To assess vessel density and pericyte coverage, co-staining was performed with CD31 (endothelial cells, BD Pharmingen 550274, 1:100) and desmin (pericytes, Invitrogen MA1-06401, 1:100). Functional vessel density was assessed in animals previously injected with FITC-lectin via co-staining with CD31. To assess tight junctions and adherens junctions proteins, co-staining was performed with CD31 and claudin-5 (tight junctions, Invitrogen 35–2500, 1:100) or vascular endothelial (VE)-cadherine (adherens junctions, Abcam ab205336, 1:50).
Singular longitudinal sections of animals previously perfused with EVB (n = 6) were also co-stained with CD31 to display the spatial and temporal distribution of EVB extravasation.
(Functional) blood vessel quantification
Vessel quantification was performed in four standardized areas per slice (ventral and dorsal horns, containing both gray and white matter) in four slices/region/animal (two regions) in n = 4 animals/group/timepoint under 20× magnification to assess the temporal and spatial distribution of vascular damage after SCI. Vessel quantification was assessed by the count of CD31 signal and perfused vessels were quantified by the count of FITC-lectin signal and declared as functional. Following, perfused–vessel ratio was obtained.
Quantification of pericyte coverage
Pericyte coverage was assessed by the blinded count of blood vessels (CD31) covered with pericytes (desmin) compared with blood vessels without associated pericytes. Assessment was also performed in four standardized areas (ventral and dorsal horns) per slice in four slices/region/animal in n = 4 animals/group/timepoint under 20× magnification.
Assessment of tight and adherens junction coverage
Tight junctions were assessed via blinded evaluation of blood vessel coverage (CD31) with claudin-5, while adherens junctions were assessed via evaluation of vessel coverage with VE-cadherine, both in four standardized areas per slice (ventral and dorsal horns) in four slices/region/animal in n = 3 animals/group/timepoint under 20× magnification. All cell quantification was performed by blinded observers using ImageJ (version 1.53).
Statistical planning and analysis
Data were analyzed with GraphPad Prism (GraphPad Software Inc., version 9.1) and SPSS (IBM SPSS software, version 28). Sample size calculation was performed with nQuery and nTermin (version 4.0). For behavioral analysis, a power of 80% was calculated to detect differences in means with an effect of Δ0.45 on a significance of α = 5% with n = 8/group. Fluorescence analyses, cell count analyses, and MRI edema volume assessments were planned exploratory. Statistical differences between groups and timepoints regarding neurological function were calculated using one-way analysis of variance with Bonferroni’s post-hoc test or unpaired t tests, as appropriate. Cell counts, EVB extravasation, and edema volume assessed by 7T MRI were calculated using Kruskal–Wallis test with Dunn’s post-hoc analysis and mixed model statistics, following averaging of every specimen’s results. Statistical significance was set at p < 0.05.
Results
Post-traumatic spinal cord edema assessed by MRI
First, the time course and extent of post-traumatic edema formation as assessed by MRI were evaluated. Following SCI or sham injury, small animal MRI (n = 5/timepoint/group) revealed significant edema formation around the injury site within the first seven days as compared with baseline before surgery (1–3 days: p < 0.01, 7 days: p < 0.05, SCI vs. baseline). Edema volume peak was reached on the first day after injury and decreased steadily over the following week. At days 14 and 28 after SCI, edema ceased to be significantly increased as compared with baseline (Fig. 2).
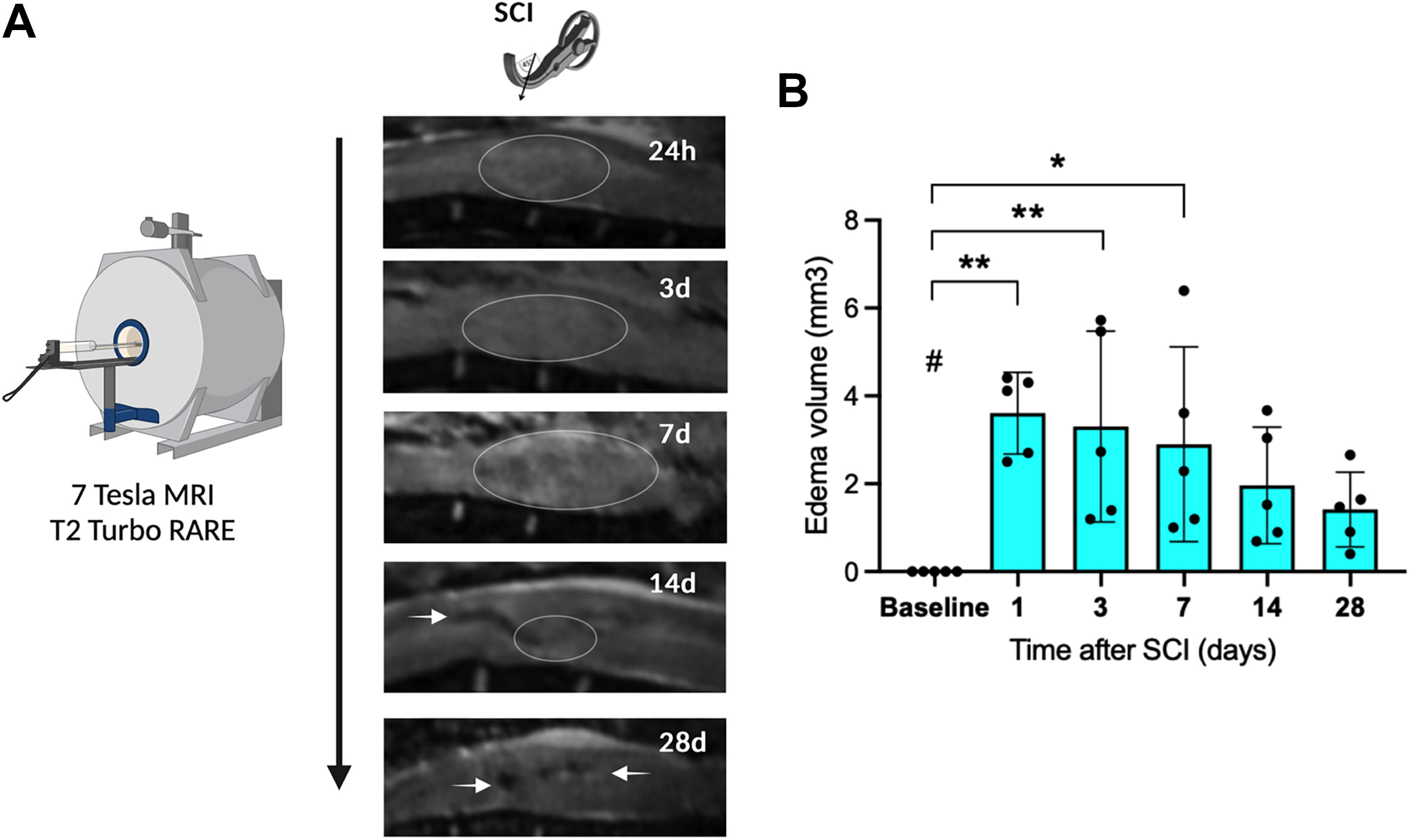
Edema formation analysis.
BSCB permeability assessed by EVB extravasation
After spinal cord explantation following EVB (64 kDa) injection, extravasation was macroscopically most prominent in the first three to seven days after SCI and less visible over a longer period (Fig. 3A).
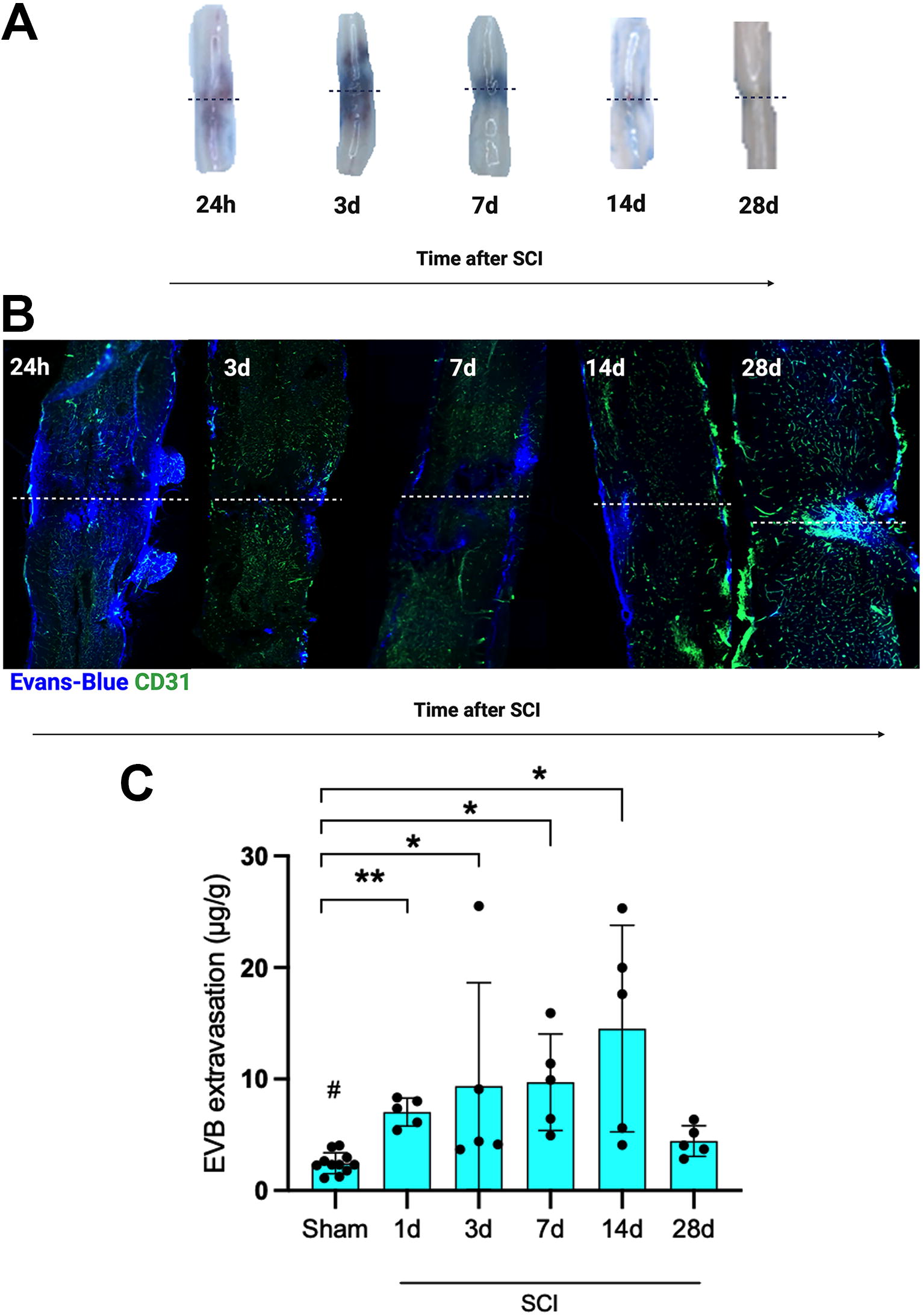
EVB extravasation analysis.
Examining longitudinal spinal cord sections after EVB perfusion and co-stained with CD31 for endothelial cells using fluorescence immunohistochemistry (n = 6), EVB extravasation was displayed most diffusely on the first day after SCI. Over subsequent timepoints, extravasation developed in a more localized pattern (Fig. 3B).
For a more detailed analysis of BSCB disruption, post-traumatic EVB extravasation was quantified. After injection of EVB dye intravenously, sham animals (n = 10) displayed no relevant EVB extravasation. In SCI animals (n = 5/timepoint), EVB extravasation was significantly increased from one to 14 days after SCI compared with sham animals, with EVB extravasation peaking after 14 days. BSCB integrity was largely restored after 28 days (Fig. 3C).
Post-traumatic spinal cord vessel perfusion
To correlate BSCB disruption and restitution with blood vessel perfusion, total and functional blood vessels were assessed (n = 4 animals/group/timepoint, Fig. 4A). Following SCI, the total blood vessel count per visual field at injury epicenter (±500 µm) decreased significantly over the first three days after injury as compared with sham animals (one day: p < 0.01, three days: p < 0.05, SCI vs. sham). After seven to 28 days, total blood vessel count returned almost to initial value or even surmounted it (seven–28 days: p > 0.05, SCI vs. sham). In the region around the injury center (±500–1000 µm, peri-trauma), total vessel count per visual field was reduced non-significantly over the first week after SCI and also slightly surmounted the initial value after 14 and 28 days (1–28 days: p > 0.05, SCI vs. sham) (Fig. 4B).
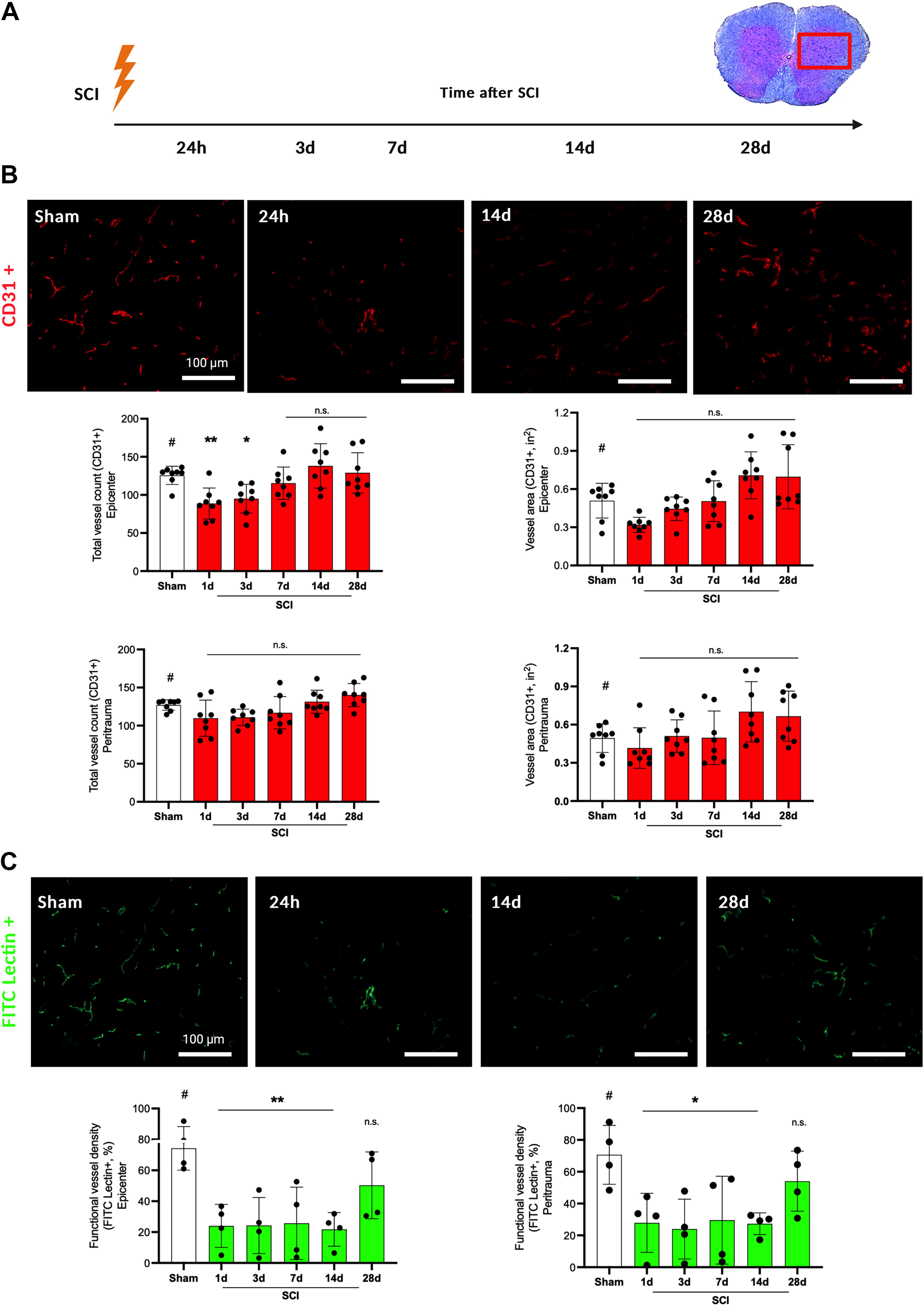
Analysis of post-traumatic blood vessel density.
Blood vessel morphology was assessed analyzing the area covered by CD31+ signals per visual field (square inches, n = 4 animals/group/timepoint, Fig. 4B). At both the injury epicenter and surrounding regions (peri-trauma), no significant differences in the area covered by blood vessels after SCI compared with sham could be observed (one–28 days: p > 0.05, SCI vs. sham). However, the area covered by blood vessels increased by day seven after SCI, surmounting initial values after 14 and 28 days non-significantly. This effect was stronger in the trauma epicenter than in the peri-trauma region.
To measure the amount of functionally perfused vessels, FITC-lectin perfusion was performed (n = 4 animals/group/timepoint) together with co-staining of FITC-lectin with CD31. During the time course from one up to 14 days after SCI, the percentage of functionally perfused vessels per visual field remained significantly diminished at injury epicenter compared with healthy animals (one–14 days: p < 0.01, SCI vs. sham). After 28 days, more vessels were functionally perfused, but without reaching the initial level (28 days: p > 0.05, SCI vs. sham, functionally perfused vessels 50–54%). This effect was similar at the peri-trauma region, however, slightly weaker (one–14 days: p < 0.05, 28 days: p > 0.05, SCI vs. sham) (Fig. 4C, all values: Supplementary Table S2).
Breakdown of the neurovascular unit
To characterize the injury to the BSCB and the NVU on a cellular level (Fig. 5A), co-staining of CD31 (endothelial cells) with desmin (pericytes), claudin-5 (tight junctions), and VE-cadherine (adherens junctions) was examined (n = 4 animals/group/timepoint).
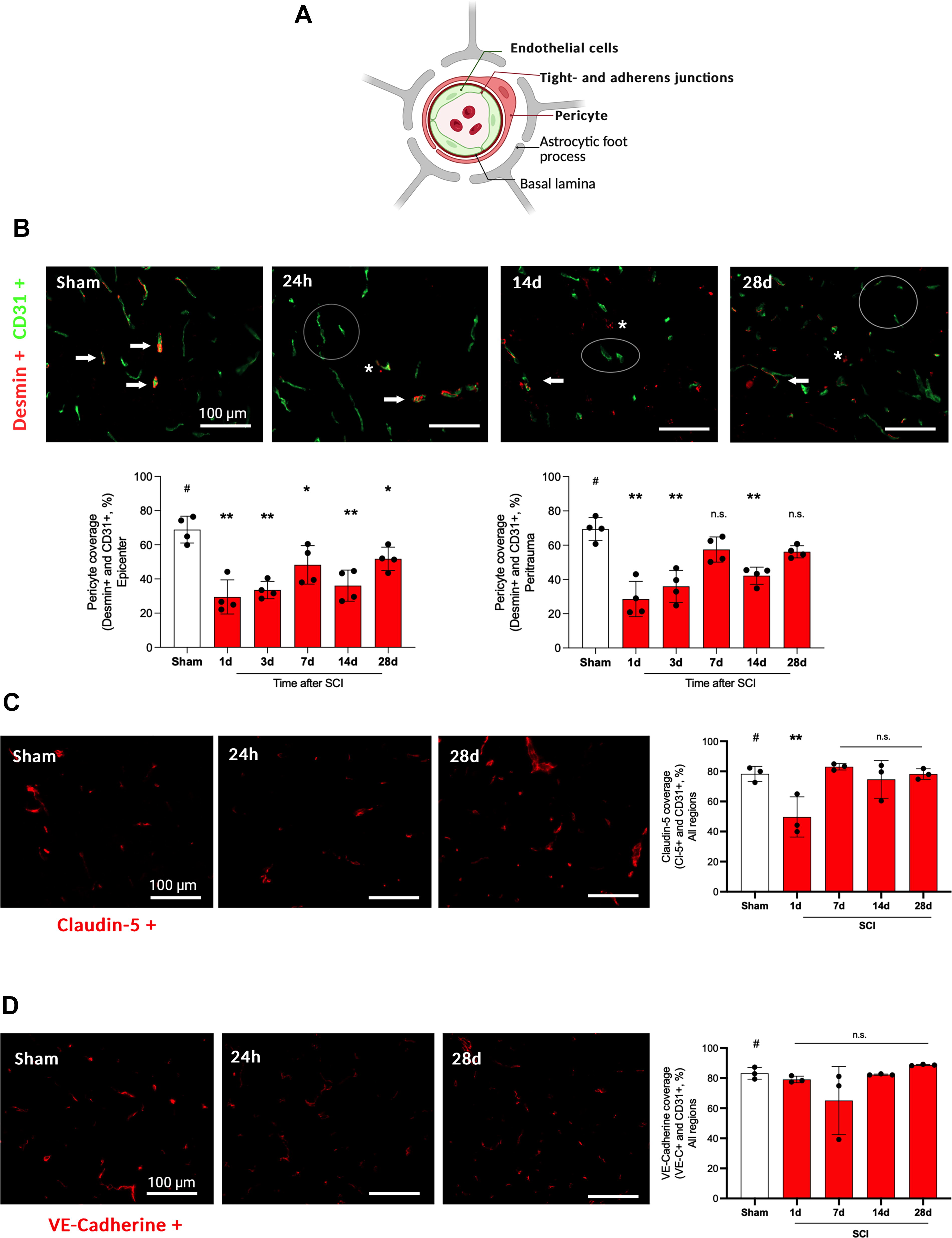
In this figure, the integrity of the NVU is examined via the assessment of blood vessel coverage with pericytes, tight junctions proteins, and adherens junctions proteins.
Looking at the trauma epicenter, pericyte coverage dropped significantly from days one to 28 after SCI with only a trend toward recovery after 28 days (one–28 days: p < 0.05, SCI vs. sham). In the peri-trauma region, no significant differences between SCI and sham-injured animals could be observed from seven to 28 days compared with sham animals (p > 0.05, SCI vs. sham) (Fig. 5B).
A significant reduction of tight junction coverage (CD31+ and claudin-5+ vessels) was displayed one day after SCI compared with sham animals (one day: p < 0.01) with normalization during the later time course from three to 28 days (p > 0.05) as compared with sham (Fig. 5C). Looking at adherens junction coverage (CD31+ and VE-cadherine+ vessels), no significant reduction was displayed over the whole time course (one–28 days: p > 0.05, SCI vs. sham) (Fig. 5D, all data: Supplementary Table S3).
Neurovascular injury assessed by longitudinal in vivo microscopy
To complement the analysis of post-traumatic vessel disruption and recovery, a live insight into the spinal cord after SCI via longitudinal in vivo microscopy was added. With a customized implanted spinal window chamber (adapted by Farrar et al. 41 ), longitudinal in vivo imaging was performed using 2PM up to the chronic phase 14 days after SCI or sham injury (n = 8/group). To examine axonal responses in the posterior columns in up to 250 µm depth, Thy1-YFP+ C57BL/6J transgenic mice were used, and prior to the examination RD (70 kDa) was injected to visualize functionally perfused blood vessels and pathological vessel permeability. Using SHG imaging, the dura mater could be differentiated for orientation purposes. (For more technical details, see Methods, and Fig. 6A–C, with [A] overview of experimental setup with implanted spinal window chamber, [B] setup of in vivo 2PM, [C] achieved microscopy depth with 2PM setup illustrated with axial histological slices with Luxol fast blue staining and Thy1-YFP signal for axons).
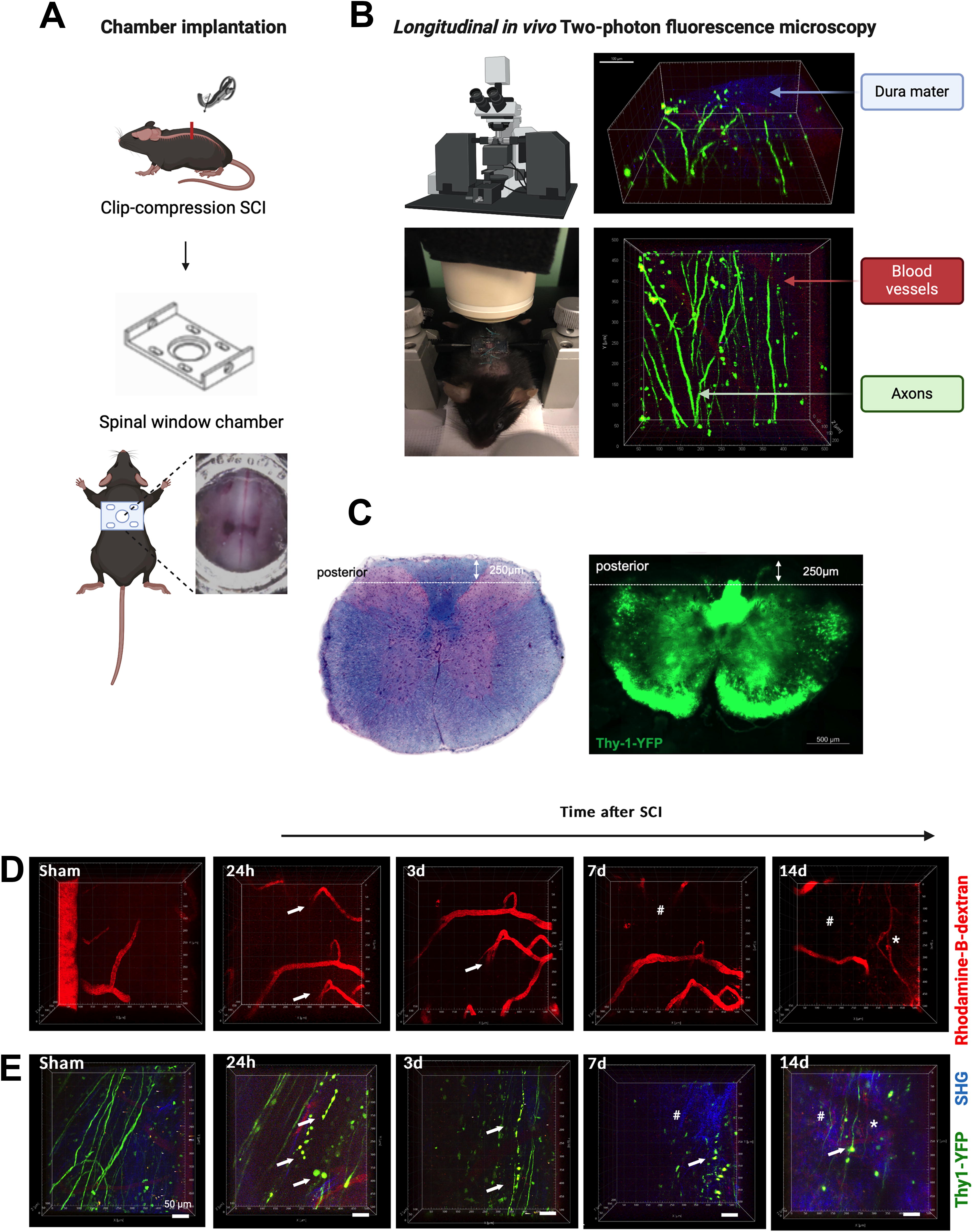
Vessel density in the visual field decreased over the first week after SCI, with slight, but allegedly non-functional revascularization visible on day 14. In sham animals, vessel count remained constant (Fig. 6D). Looking at both Thy1-YFP+ signals and perfused vessels (RD+), axonal degeneration after injury could be visualized starting from the first day, with axonal end bulb retraction visible during ongoing axonal degeneration over time, while associated with functional vessel degradation after seven and 14 days. Thus, a spreading of the initial injury could be observed in assessable regions at farther distance (up to 500 µm in both directions) from the injury epicenter (Fig. 6E). In sham animals, axons and blood vessels remained intact and visible over time.
Neurobehavioral recovery after SCI
Following the application of thoracic clip-compression SCI, mice were paraplegic and required meticulous care (Supplementary Fig. S1). To monitor neurobehavioral recovery, BMS and Tally subscore for recovered hindlimbs were performed in an open field after one, three, seven, 14, and 28 days (n = 10/group, sham/SCI). Hindlimb function as tested with BMS and Tally subscore started to recover slightly after three days, with substantial recovery displayed starting one week after injury, with further increase during follow-up. However, neurological function remained significantly impaired over the whole time course up to 28 days as compared with sham animals (BMS and Tally subscore at one–28 days SCI as compared with sham: p < 0.01) (Fig. 7A).
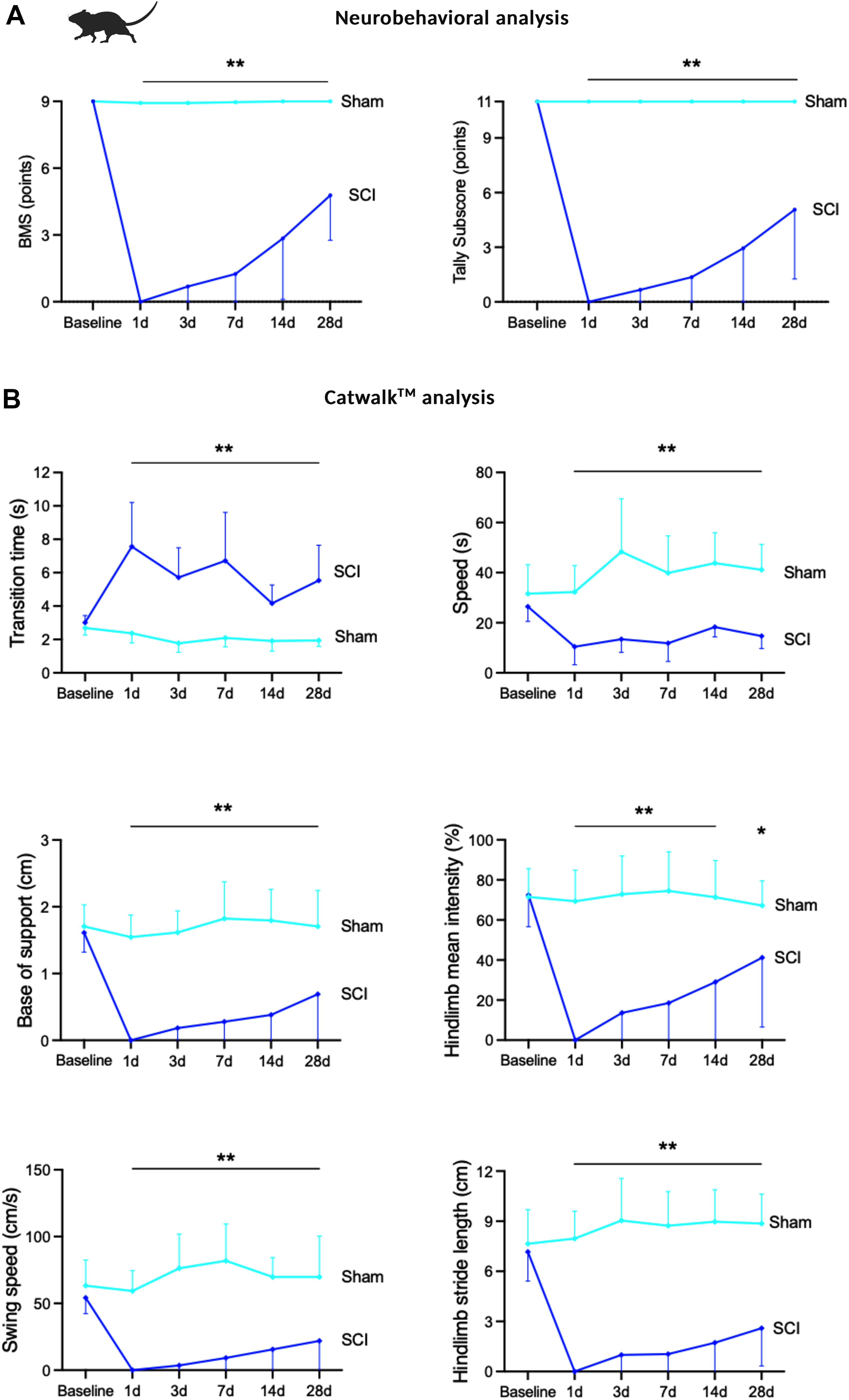
Analysis of neurobehavioral recovery after SCI.
Additionally, CatWalkTM automated gait analysis was performed at the given timepoints in ten animals per timepoint and group, further displaying substantial hindlimb recovery starting at post-traumatic days seven until the end of the examination (day 28) in all examined parameters, comprising of parameters for speed analysis, gait analysis, and stride analysis (transition time, speed, base of support, hindlimb mean intensity, swing speed, and hindlimb stride length). However, residual significant impairment was at all times apparent following SCI when compared with healthy animals (all parameters one–28 days SCI vs. sham: p < 0.005, hindlimb mean intensity at 14 days: 0.005, 28 days: p = 0.0034, Fig. 7B, see all data: Supplementary Table S3).
Discussion
The principal novel finding of this study is the definition of a therapeutic window of opportunity for neurovascular unit repair within the first days up to two weeks after experimental SCI. Within this work, the spatial and temporal distribution of the post-traumatic damage to the BSCB and the cells of the NVU as well as its endogenous repair were characterized over the acute into the chronic injury phase after SCI. This insight into the temporal and spatial regeneration of the spinal cord is complemented by the display of the unfolding secondary injury via longitudinal in vivo 2PM through an implanted spinal window chamber up to two weeks after injury. With this, axonal injury and neurorecovery were defined as experimental readouts for future studies testing therapies targeting neurovascular unit repair.
BSCB permeability
Post-traumatic edema formation as assessed by MRI volumetry in this study peaked in the first three days after injury and was restituted thereafter, while EVB permeability quantification by fluorescence assay revealed a thoroughly impaired BSCB into the chronic phase two weeks after injury, with significant restitution only after 28 days. In vivo 2PM in this study, however, supports the assumption that the clinically most relevant permeability changes of the BSCB are to be met in the first days after injury, as pathological permeability could not be visualized later than day three.
According to this study, the ideal timeframe identified for therapy applications set to overcome the physiological barrier of the BSCB after SCI would be in the first days up to two weeks after injury. This could be especially relevant, as the pathological opening of the BSCB aggravates and upholds the secondary injury cascade after SCI and thus poses a desirable target for specific therapies aiming to seal this pathological permeability. 5,8,24,44
Moreover, most therapies in clinical studies are applied during the acute injury phase, following the surgical decompression of the injury site. 7,45,46 Our findings support the assumption that the application of local therapies over a longer time period after acute SCI might be desirable. Therefore, research into devices that permit constant local therapy delivery in situ seems promising. 6,8,46 –52
MRI edema and 2PM permeability assessment in this setting seemed to have a lower sensitivity and were definitely more observer-dependent than automated EVB fluorescence assay. 43,53 However, EVB with 64 kDa, while still being the gold standard for permeability analyses according to the literature, 43,53,54 is a rather large molecule, as is RD, which was used for 2PM (70 kDa) in this study. Experimental studies using contrast-enhanced MRI with gadolinium complemented by immunohistochemistry with albumin (66 kDa) were able to state restitution of the BSCB variably from seven up to 56 days after SCI. 22,55 It is to be assumed that the application of smaller molecules like small dextrans (4–10 kDa) or small-molecule MRI contrast agents in this setting would possibly reveal a longer opening of the BSCB due to a higher sensitivity to much smaller leakages. However, the clinical relevance of such minimal BSCB permeability changes remains to be examined. 17,18,21,56,57
Post-traumatic spinal cord vessel perfusion and injury to the NVU
In this study, spinal cord blood vessel count decreased significantly over the first three days after SCI, returned almost to baseline after one to two weeks, and even surmounted baseline after four weeks, with less vascular injury assessable in the region surrounding the injury epicenter. Vessel morphology differed starting from one week after SCI displaying a wider and more flexuose morphology, quantifiable through a larger vessel-covered area, with this effect being more pronounced in the trauma epicenter. Functionally perfused vessel density, however, remained significantly diminished compared with healthy animals at all timepoints in both the epicenter and peri-trauma regions. After four weeks, only 50–54% of spinal cord vessels were functionally perfused.
These results elucidate that revascularization takes place as an endogenous repair process in the injured spinal cord but is insufficient in supplying functionally perfused vasculature and thus to effectively diminish tissue hypoxia after SCI. 44,58 –60 Therapeutic enhancement of these insufficient endogenous processes would be promising in promoting tissue perfusion and consequent structural regeneration. 24,25
The therapeutic potential of this approach is furthermore highlighted by the results of in vivo 2PM in this study, visualizing the progressive degeneration of initially preserved spinal cord axons in the peri-traumatic zone after SCI while displaying the rarefication of functional vessels over the acute to subacute secondary injury phase. These findings support the hypothesis that axons necessitate functional vessels for perseverance and regrowth. 25,28,31,32
In search of potential targets to promote functional vessel regeneration after SCI, this study characterizes the post-traumatic responses of the known players of the BSCB and NVU: pericytes, adherens junction, and tight junction proteins. 10,61 –64 Pericyte coverage of spinal cord vessels in this study dropped significantly at all examined timepoints after SCI with only a trend toward recovery over the observed period, displaying differences between trauma epicenter and surrounding regions. Tight junction proteins were depleted in the acute injury phase and restituted quickly afterwards, while no significant changes in the quantification of adherens junction proteins could be stated. These findings demonstrate the thorough injury to the integral parts of the NVU in the first days after SCI, significantly impairing cell–cell contacts between pericytes and endothelial cells, with partial restitution of this system already taking place within the first few days after injury.
Especially, pericytes seem to play a multifaceted role in SCI and subsequent spinal cord regeneration, as pericytes are known to be essential for blood vessel growth and maturation, are closely related to the form and function of endothelial cells, and communicate with endothelial cells via cell–cell contacts as a central part of the NVU. 65 –70 However, there exist different pericyte subtypes and some subtypes are involved in vascular contractment benefitting local hypoxia, others promote revascularization, while others can differentiate into neurocytes, and others play a role in fibrotic scar formation. 66,67,69 –71 Given these differing functions, further characterization of their temporal and spatial involvement of pericytes in secondary injury development after SCI and spinal cord regeneration is worth studying in more detail. 70
Neurobehavioral recovery
Neurobehavioral analysis in this study revealed substantial improvement of hindlimb function starting from one week after injury. However, significant impairment persisted over the whole observation period of four weeks. This shows that although a functionally complete SCI is initially achieved within the used model, spinal cord regeneration that leads to functional regeneration can be systematically assessed over the given observation period, starting in the subacute phase. 38,72 –74 Moreover, neurobehavioral recovery as displayed in this study is paralleled by the structural regenerative changes taking place, like the partial recovery of functional vessel density and the partial recovery of the BSCB and NVU.
The utilized SCI model, however, remains a moderate to severe SCI model, with the limitation that earlier and slighter influences of structural regeneration on functional recovery might be missed. In the planning of future experiments studying the influence of single molecular therapeutic targets on spinal cord regeneration, the use of more moderate to mild SCI models might be considered. 33,34,75 –77
Conclusions
This study reveals a therapeutic window for applications targeting a pathologically hyperpermeable BSCB as part of an impaired neurovascular unit from the first days up to two weeks after traumatic SCI. Moreover, partial regeneration of functional vasculature is detected in the subacute to chronic phase two to four weeks after injury, however, with only 50–54% of existing vessels regaining functional perfusion.
The therapeutic potential of ameliorating functional vessel regeneration in this study is highlighted by the results of immunohistochemistry and longitudinal in vivo 2PM, visualizing the progressive degeneration of initially preserved spinal cord axons in the peri-traumatic zone after SCI, although displaying a rarefication of functional vessels in the acute to subacute injury phase for up to two weeks after injury. These findings support the hypothesis that axons and regenerating tissue necessitate functional vasculature for perseverance and regrowth. This incomplete structural regeneration is complemented by an incomplete neurobehavioral recovery over the whole observation period of four weeks.
In the search for restorative therapies driving functional recovery after traumatic SCI, the continuous application of therapies targeting functional vessel regeneration appears promising at a time when both impaired permeability of the neurovascular unit and ongoing endogenous revascularization exist. Continuous multifactorial therapy applications targeting the integrity of the neurovascular unit over the identified therapeutic window could aid ameliorating functional vessel perseverance and the spinal cord’s regenerative capacity.
Transparency, Rigor, and Reproducibility Statement
The datasets supporting the conclusions of this article are included within the article and its supplemental files. Additional material is available from the corresponding author upon reasonable request. The animal experimental study is registered on the platform animaltestinfo.org (NTP-ID 00019011–1-7).
Footnotes
Acknowledgments
The authors thank the Fehlings-Laboratory, especially Christopher Ahuja, MD, PhD, Anna Badner, PhD, Liu Yang, and Anoushka Singh of Toronto Western Hospital and Krembil Neuroscience Institute, Toronto, Canada for equipping the authors with the clip-compression SCI model and for their support and mentorship throughout the experiments. Moreover, the authors thank the experimental neurology department at Charité Berlin under the supervision of Prof. Andre Rex, Prof. Harald Prüß, and Larissa Mosch for their support in conducting the neurobehavioral analyses. The authors furthermore thank Dr. vet. Claudia Abramjuc, Dr. vet. Anja Schulz, Dr. vet. Hannah Theresa Nickles, and Dr. vet. Stephan Nagel, as well as Johanna Hössler of the LaGeSo Berlin for their support in planning and conducting the animal experiments according to the principles of good laboratory practice and according to the national animal welfare guidelines. The authors also thank Dr. Andrea Stroux from the Department of Biometry and Clinical Epidemiology at Charité Berlin for her help with statistical planning and sample size calculation. Figures were created with GraphPad Prism (GraphPad Software Inc., version 9.1) and BioRender.com.
Authors’ Contributions
P.V., M.G.F., and V.H. designed the study concept. V.H. and M.N. performed the animal surgeries. V.H., L.W., L.M., N.T., L.R., K.K., and E.v.B. performed neurological assessments, CatWalk® analysis and animal care, and helped with tissue processing for histological analysis. A.R. and H.P. provided the CatWalkTM analysis infrastructure and supervised the neurobehavioral experiments. V.H. and I.K. supervised the histological and immunohistochemical tissue processing and staining and Evans blue fluorescence assay. L.W., L.M., N.T., and I.K. performed the post-mortem tissue processing, Evans blue assays, and histological analysis. J.E.O. helped with constructing and providing the spinal window chamber. C.U. and K.S. helped with the establishment of in vivo microscopy after spinal window chamber implantation. A.R., A.E.H., and R.N. helped with the establishment of 2-photon spinal cord microscopy, provided the 2-photon microscopy infrastructure, and supervised the 2-photon experiments. V.H. performed the data analysis and wrote the article. L.W., L.R., L.R., K.K., E.v.B., and N.T. performed data acquisition and parts of data analysis and revised parts of the article. P.V., F.L.H., and M.G.F. supervised the experimental work and revised the article. P.V. and M.G.F. share the senior authorships of this work.
Ethical Approval
All procedures performed in this study involving animal subjects were in accordance with the ethical standards of the institutional research committee and with the ISO certification guidelines. Animal experimental approval (G031417) was granted by the institutional ethics board of the LaGeSo (Landesamt für Gesundheit und Soziales), Berlin, Germany.
Author Disclosure Statement
No conflicts of interest exist in the submission of the article and the article is approved by all authors for publication.
Funding Information
This work was funded by the EANS (European Association of Neurosurgical Societies) via the
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.