
Abstract
Contusive and compressive spinal cord injury (SCI) induces pathological changes to spinal cord white matter (WM) including periaxonal swelling and resultant disruption of the axomyelinic interface, axonal swelling/spheroid formation, and secondary axonal transection. To further our knowledge of the role of vascular edema in these pathological changes to WM, we designed, and three-dimensional (3D) printed a dual-compartment imaging chamber separated by a semipermeable membrane to mimic and manipulate interstitial and vascular fluid compartments in real time. We hypothesized that hypertonic saline (HTS) applied to the “vascular” chamber would osmotically shift fluid out of the periaxonal space and preserve myelinated fibers after SCI. Adult male and female 6- to 8-week-old Thy1YFP+ transgenic mice underwent a C5, mild contusive SCI (30 kilodyne, IH Impactor) in vivo, and their spinal cords were harvested for ex vivo imaging. Utilizing longitudinal two-photon excitation microscopy (2PE), we imaged both myelin (Nile red) and axons (YFP+) simultaneously up to 4 h after SCI. C5 contusive SCI conditions induced significant increases in periaxonal swelling and axonal spheroid formation within the dorsal column fibers over time. In contrast, perfusion of 3% and 5% HTS in the “vascular” compartment beginning 30 min after SCI was highly protective and significantly reduced periaxonal swelling and axonal spheroid formation from 1 h 30 min to the last hour recorded (4 h post-SCI) compared to normal saline (NS) controls. At 2 post-SCI, treatment with 3% and 5% HTS significantly (Kruskal–Wallis ANOVA on Ranks, H(3) = 3, p = 0.05, n = 5–6/group) reduced periaxonal swelling compared to NS (median, 25th percentile; 11.00, 4.00 versus 9.00, 7.00 versus 48.00, 29.50, respectively; Dunn’s method, both p < 0.05). By 4 h post-SCI, treatment with 3% and 5% HTS significantly (H(3) = 15.74, p = 0.001, n = 5–6/group) decreased axonal spheroids compared to NS (5.00, 3.00 versus 4.00, 3.00 versus 95.00, 38.75, p = 0.001, p < 0.001, respectively). In contrast, 7.5% HTS had no beneficial effect. Collectively, these data provide insight into the dynamic interplay between interstitial fluid exchange within the periaxonal space and pathological changes in myelinated fibers following SCI. Delayed in vivo administration of 3% HTS significantly increased axonal survival and reduced periaxonal swellings 24 h post SCI compared to NS control, validating the translatability of our dual compartment imaging chamber (mean, standard deviation; 58.09, 3.34 versus 32.08, 5.98, p = 0.003; 595.19, 326.10 versus 1525.25, 259.82, p = 0.018, respectively). Our findings suggest that low-dose hypertonic solutions may have a protective effect in part by mitigating periaxonal swelling and thereby potentially reducing the occurrence of axonal spheroids within these denuded regions. These results enhance understanding of secondary axonal degeneration mechanisms and hold promise for targeted therapeutic interventions to improve outcomes in SCI.
Keywords
Introduction
Traumatic spinal cord injury (SCI) elicits a delayed secondary degenerative response that causes the loss of additional white matter (WM) that survives the initial impact but then degenerates over time. Importantly, this delayed secondary “bystander” loss of WM exacerbates neurological recovery after injury and is therefore a viable and important target to improve outcome after SCI.1–3 Ultrastructural studies of SCI in preclinical animal models and in humans have reported that periaxonal swelling, (i.e., when the periaxonal space enlarges and disrupts the axon myelin interface), axonal swelling and spheroid formation, myelin vesiculation, and secondary axonal transection are prominent features of WM pathology acutely after SCI.4–8 However, the underlying mechanisms remain incompletely understood. As such, clinically viable treatments to prevent WM loss after SCI remain elusive.
It is well established that traumatic SCI causes ionic dysregulation in WM and concomitant fluid–electrolyte disturbances that play a causative role in WM injury.1,2,9 Using longitudinal intravital imaging of genetically encoded Ca2+ indicators in axons, it has been shown that intra-axonal Ca2+ levels increase after SCI in axonal swellings and influence axonal fate.4,5 Furthermore, inhibition of Na channels and Na–Ca exchangers in models of WM injury are highly protective and improve neurological recovery after SCI. 9 Moreover, significant and progressive edema occurs after SCI and can spread above and below the epicenter of injury in humans6,7 and experimental models.8,10–12 Edema formation is evident within hours and peaks ∼3 days after injury but remains elevated weeks after injury.11,13,14 Fluid resuscitation using hypertonic saline (HTS) is Food and Drug Administration approved for hyponatremia and cerebral edema with increased intracranial pressure.15,16 HTS fluids have a much higher concentration of solute compared to both plasma and interstitial fluids and thereby increase the osmolarity of the blood driving fluid from interstitial spaces to the intravascular space.16,17 This in turn reduces edema and improves cerebral blood flow. 16 Despite several studies showing HTS improves aspects of neurological recovery after SCI,17–20 the effects on WM pathophysiology remain poorly understood.
Given that periaxonal swelling after SCI in vivo spatiotemporally overlaps with edema formation, and blood–spinal cord barrier breakdown, we predict that interstitial fluid accumulation negatively impacts the axon myelin interface. 21 Hypertonic solutions, by osmotically extracting water from the interstitial space, may reverse fluid accumulation and prevent periaxonal swelling. In turn, this could reduce the formation of axonal spheroids, as our previous data suggest that periaxonal swelling precedes and is negatively correlated with axonal spheroid formation 21,22
Here, we designed and manufactured a three-dimensional (3D)-printed dual imaging chamber that allows us to image the contused spinal cord ex vivo, while simultaneously modifying the extracellular fluid. The spinal cord chamber is separated by a semipermeable membrane that allows the “vascular compartment” of the chamber to be experimentally manipulated with HTS and drive water out of the interstitial compartment into the vasculature compartment of the chamber (Fig. 1).
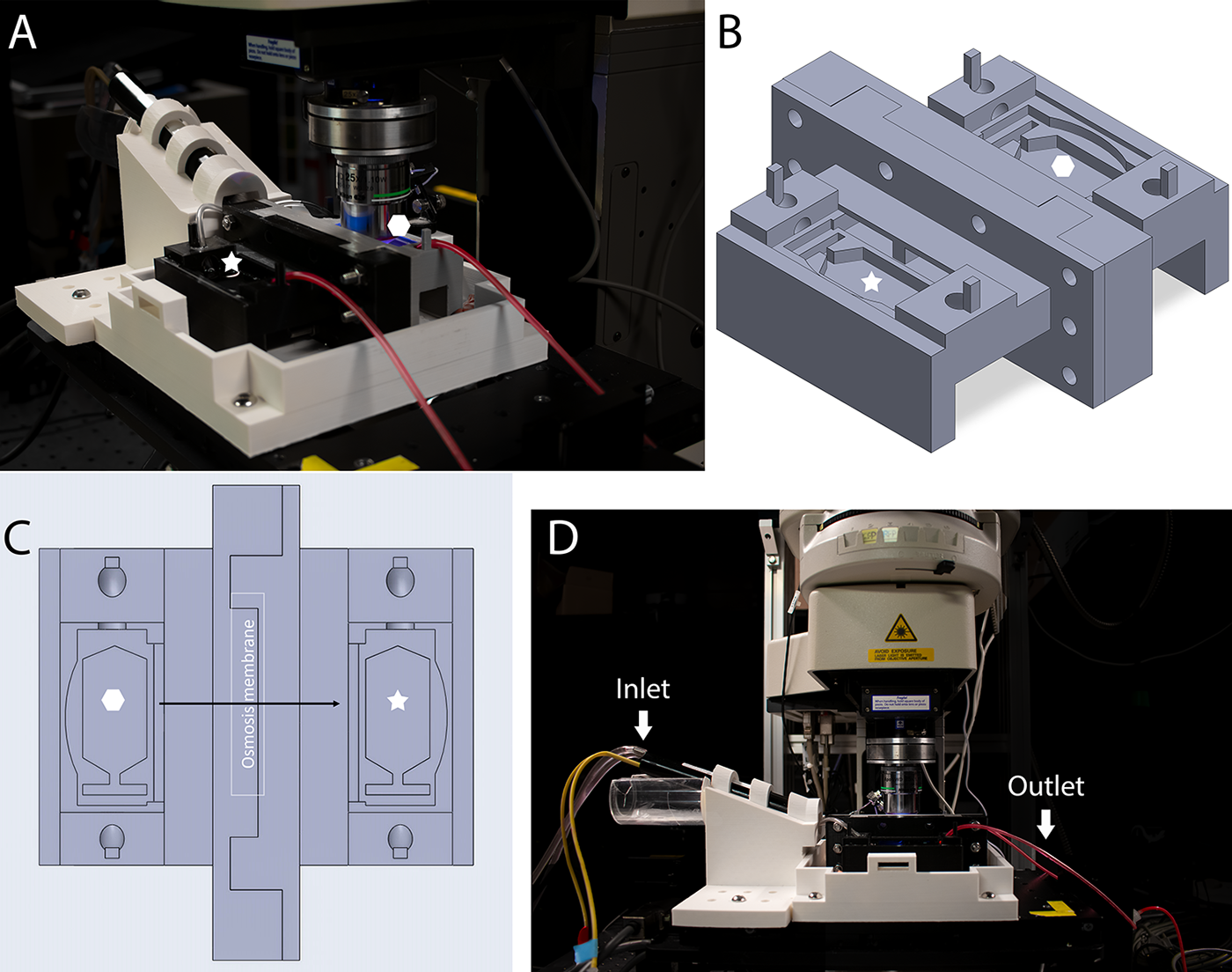
Overview of the 3D-printed dual imaging chamber setup.
To determine the effects of HTS on periaxonal swelling and axonal spheroid formation following C5 contusive SCI, we used our previously characterized ex vivo spinal cord model that allows time-lapse two-photon excitation imaging to track dynamic changes in axons and myelin in real time acutely after SCI. We assessed three different clinically relevant HTS concentrations versus normal saline to determine their effectiveness in mitigating periaxonal swelling and axonal spheroid formation following a cervical SCI in real time.
Materials and Methods
Cervical SCI and ex vivo spinal cord isolation
All experiments were conducted in accordance with the University of Louisville Institutional Animal Care and Use Committee, adhering to the Guide for the Care and Use of Laboratory Animals. Adult 6–10-week-old male and female Thy1YFP+ transgenic mice (IMSR_JAX: 003782; The Jackson Laboratory, Bar Harbor, ME) were bred in house. The ex vivo spinal cord model used here has been previously described in detail.22–25 Mice were anesthetized with an intraperitoneal injection of 100 mg/kg ketamine/10 mg/kg xylazine. The surgery site was shaved, disinfected, and incisions were made to expose and dissect the overlying musculature exposing C5/6 vertebrae. A laminectomy was performed at C5/6, then mice received a C5, 30 kdyn contusive SCI in vivo (IH impactor, PSI, Lexington, KY, USA). Mice were then euthanized with an intraperitoneal injection of 500 mg/kg of ketamine, perfused transcardially with ice cold low Ca2+ artificial cerebrospinal fluid bubbled in 95% O2/5% CO2 to keep the spinal cord viable, and the cervical region of spinal cords were removed and placed in a customized imaging chamber with two compartments. The perfusion bath was changed to oxygenated 2 mM Ca2+ artificial cerebrospinal fluid (aCSF) in the compartment containing the spinal cord. The temperature of the aCSF was held constant at 35–37°C by an inline heater (Warner Instruments). The adjacent “vascular” compartment was perfused with either normal saline (control group), 3%, 5%, or 7.5% NaCl solution (n = 5–6/group) from the moment the spinal cord was transferred into the chamber. An osmosis membrane (FTSH2O Flat Sheet Membrane, CTA, FO, CF016; Sterlitech, Kent, WA, USA) was inserted between both compartments to facilitate osmosis, where the HTS solutions draw water out from the aCSF perfusing the spinal cord. To visualize myelin, the lipophilic dye Nile Red (Cat# N1142; ThermoFisher Scientific, Waltham, MA, USA) was first dissolved in dimethylsulfoxide (5 mM; VWR, Radnor, PA, USA) and used at a final concentration of 25 μM, then directly and briefly added to the perfusion chamber containing the spinal cord (custom built) in 5 μL increments until clear labeling of myelin surrounding YFP+ axon profiles was obtained. Importantly, using this experimental perfusion system, the spinal cord remained viable up to 12 h after isolation based on morphological observation of axons, oligodendroglia, and myelin, as previously reported. 23
Imaging chamber with two compartments
The imaging chamber was designed, and 3D printed with two compartments to investigate the effects of fluid resuscitation on WM pathology after SCI. The overall experimental setup aligned with the ex vivo spinal cord chamber model previously described, 23 with a notable modification, substituting the original imaging chambers with the one depicted in Figure 1.
Figure 1 provides an overview of the 3D model of the imaging chamber and its setup during experiments. The imaging chamber design, created using SOLIDWORKS 2022, was fused filament fabrication 3D printed through the Additive Manufacturing Institute of Science and Technology at the University of Louisville.
In vivo imaging
In vivo images were acquired following a previously described protocol. 21 In brief, six adult (8–12 week old) Thy1YFP+ mice were anesthetized with isoflurane, received buprenorphine (1 mg/kg; ZooPharm, Laramie, WY), and given 2 mL sterile saline SC. Next, mice received a T13 laminectomy after which dilute Nile Red (500 μM; Cat# N1142; ThermoFisher Scientific, Waltham, MA) was applied to the intact dura for several minutes. Next, mice were placed below the A1RMP+ multiphoton microscope, and baseline (laminectomy only) images were collected. Immediately following imaging, mice were given a 30 kdyn SCI with a dwell time of zero seconds. Mice were then taken of anesthesia and received postoperative care as previously described. At 3 h post-SCI, the tails of the mice were warmed with warm water to induce vasodilation and either NS or 3% HTS (5 μL/g) was slowly administered intravenously via the lateral tail vein. Mice were then allowed to recover until 24 h post-SCI when mice were anesthetized again and reimaged. Following 24 h imaging, mice were euthanized as described above.
Microscopy
To visualize the dynamic response of dorsal column axons following contusive SCI, time-lapse two-photon excitation images of fluorescently labeled dorsal column axons and myelin were obtained using a commercial A1RMP+ multiphoton microscope (Nikon Instruments Inc., Melville, NY) and Elements software (Nikon Instruments Inc.). Briefly, the contusion site of the spinal cord was aligned under a 25×, 1.1 NA objective, and excited with a wavelength of 950 nm (∼17 mW measured at the exit of the objective) for simultaneous Nile Red and YFP imaging. The fluorescence emitted was filtered through appropriate dichroic and bandpass filters to isolate the fluorescent emission of the fluorophores, as previously described.23,24
Image analysis
Time-lapse image recordings of the contused cervical region were collected in z-stacks (2.0 µm/step) at 40 min, 1 h, 1 h 30 min, 2 h, 3 h, and 4 h post-SCI. Data were then imported into ImageJ for visualization and analysis. Periaxonal swelling and axonal spheroids counts were quantified and expressed as periaxonal swellings or axonal spheroids normalized to imaged area (20893 µm2). Here, periaxonal swelling is defined when the myelin detaches from its axon (creating a periaxonal space between the myelin and its axon), and axonal spheroids are swellings with a size twice the width of the axon. For in vivo images, periaxonal swellings were quantified as described above, and axons spared 24 h post-SCI were tabulated and compared to their baseline conditions and reported as percent axonal sparing (mean and standard deviation). All quantification of images was done blind to treatment.
Statistical analysis
Statistical analysis for all imaging experiments was performed using IBM SPSS Statistics (Version 29) and graphs created using MATLAB R2024a. The data were first assessed for normality using the Lilliefors test and equal variance using the Levene test, both in MATLAB. Due to the non-normal distribution of the data, nonparametric tests were used. Differences in periaxonal swelling and axonal spheroid formation across the different treatment groups (normal saline, 3%, 5%, and 7.5% NaCl) and over time (from 40 min to 4 h post-SCI) were assessed using the Kruskal–Wallis ANOVA on Ranks, followed with Dunn’s method of multiple pairwise comparisons where significant differences were detected. Data were expressed as median and 25th and 75th percentiles. In vivo statistical outcomes were assessed with t-tests, and data were expressed as mean ± standard deviation (SD). Statistical significance was set at p < 0.05.
Results
Longitudinal imaging revealed that in sham-only (no SCI) conditions, myelinated spinal cord axons appear linearly arranged, with myelin closely opposed to their axons, without detectable gaps or signs of myelinated fiber pathology (Fig. 2A–C). In distinction to sham conditions, at 1 h post-SCI, all groups showed initial signs of periaxonal swelling but minimal amounts of axonal spheroids (Fig. 2D–G). In normal saline (NS, 0.9% NaCl) conditions, there was a gradual increase in axonal spheroids and by 4 h post-SCI many of the regions of myelinated fibers with periaxonal swelling became filled with axonal spheroids (Fig. 2H, L, P, T). Despite an initial increase in periaxonal swelling at 1 h post-SCI, the 3% HTS-treated spinal cords appeared to develop fewer regions of periaxonal swelling and spheroids from 2 h to 4 h after SCI (Fig. 2E, I, M, Q, U). Similarly, there appeared to be less axonal spheroids in 5% HTS-treated spinal cords (Fig. 2F, J, N, R, V). Surprisingly, the 7.5% HTS appeared less protective than both low-dose HTS treatment groups, especially the 3% HTS group, and more closely resembled the control NS group from 1 h 30 min to 4 h after SCI (Fig. 2G, K, O, S, W).
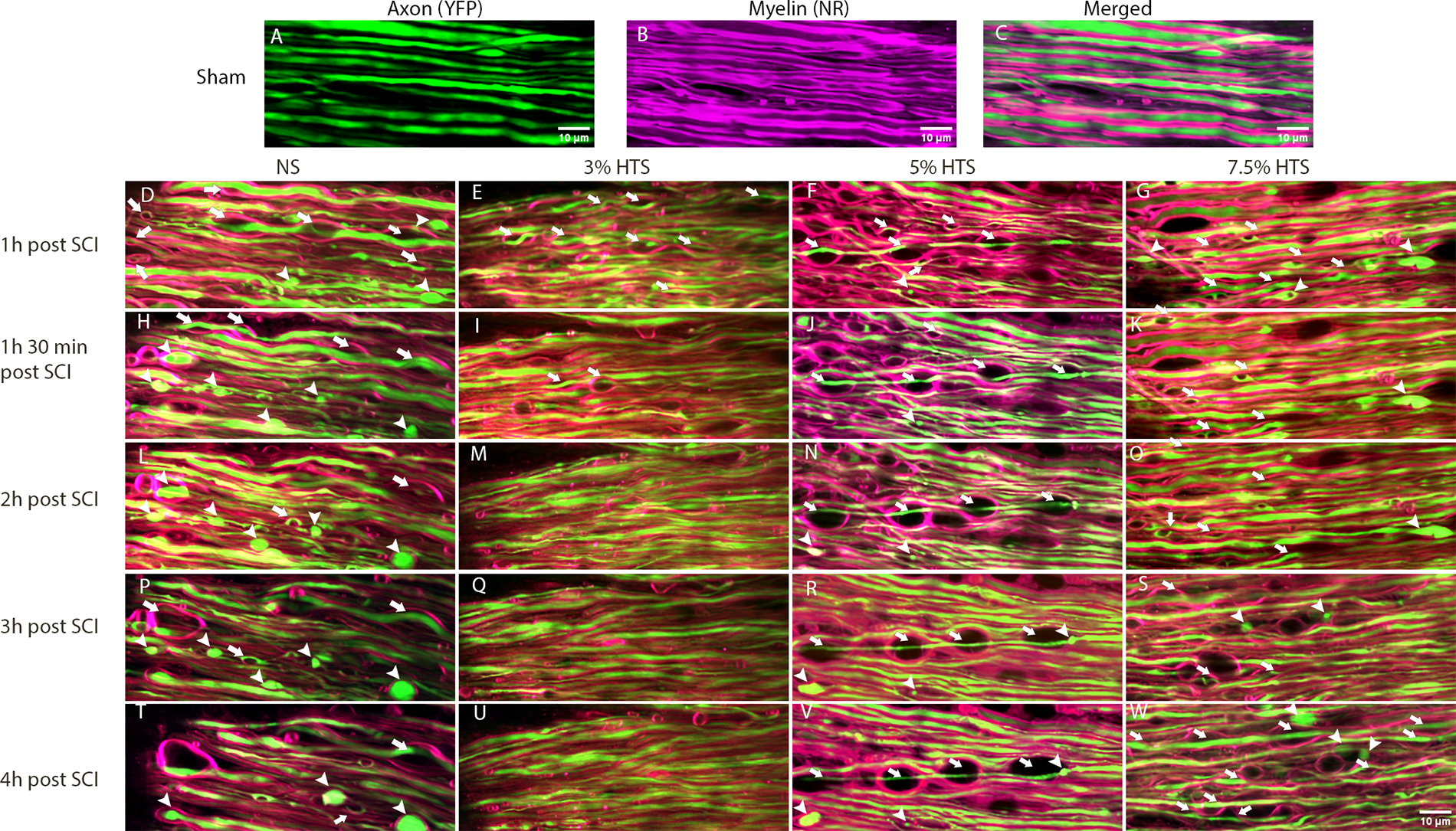
Longitudinal intravital imaging of myelinated cervical spinal cord axons post-SCI. Representative MIPs of axons (YFP, green,
To better characterize the progressive changes in axonal morphology and the effectiveness of low dose HTS treatment, instances of periaxonal swelling and axonal spheroids were quantified from the first time-lapse images taken at 40 min post-SCI to 4 h post-SCI. The data were then compared between groups at each time point. At 40 min post-SCI, there were no significant differences (ANOVA on Ranks) among the HTS treatment groups and NS controls both for periaxonal swelling (H(3) = 1.92, p = 0.59) and axonal spheroids (H(3) = 15.74; p = 0.23, Figure 3A, B). Quantification of periaxonal swelling revealed that treatment with 3% and 5% HTS significantly (ANOVA on Ranks, H(3) = 3, p = 0.05, n = 5–6/group) decreased periaxonal swelling following SCI compared to NS at 2 h post-SCI (median, 25th percentile; 11.00, 4.00 vs 9.00, 7.00 vs 48.00, 29.50, respectively; Dunn’s method, both p < 0.05, Fig. 3A). Together, these results show that 3% and 5% HTS but not 7.5% HTS reduce instances of periaxonal swelling following contusive SCI.
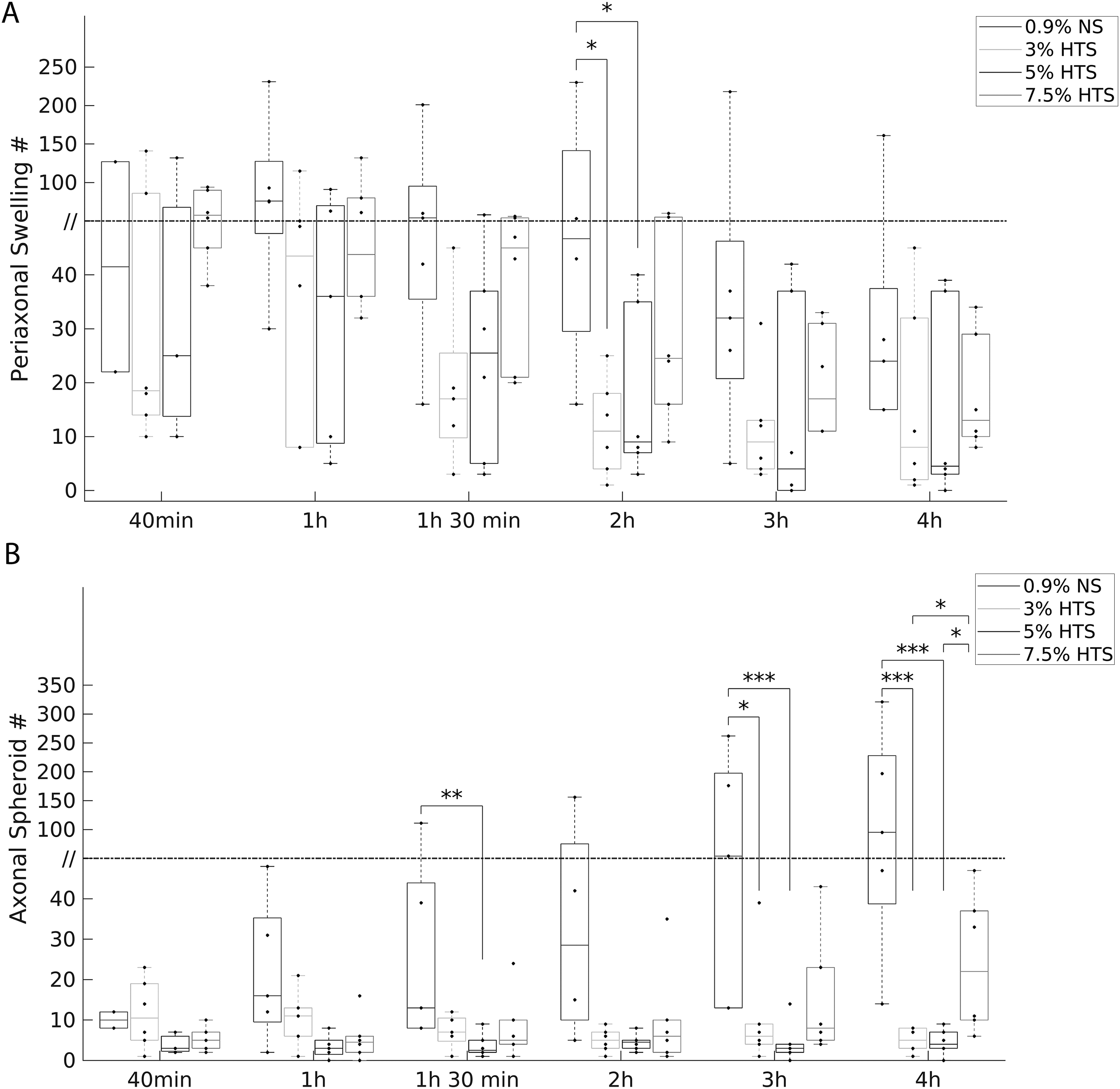
Temporal distribution of periaxonal swelling and axonal spheroid formation after SCI.
Similar to periaxonal swelling, quantification of axonal spheroids showed pronounced differences among the treatment groups post-injury (Fig. 3B). For the NS group, axonal spheroids increased markedly from the initial time point at 40 min (10.00, 8.00; median, 25th percentile) to 4 h (95.00, 38.75) post-SCI. In contrast, the 3% HTS treatment group had significantly less (6.00, 4.00; 10.50, 5.00; both p = 0.001) axonal spheroids at 3 h and 4 h post-SCI compared to the NS group (54.00, 13.00; 95.00, 38.75, respectively). Furthermore, the 5% HTS treatment had significantly less axonal spheroids at 1 h 30 min (ANOVA on Ranks, H(3) = 7.96, p < 0.05, n = 5–6/group), 3 h (H(3) = 10.97, p = 0.012), and 4 h post-SCI (H(3) = 15.74, p = 0.001; Dunn’s multiple comparisons, p < 0.01, p = 0.001, and p < 0.001, respectively) compared to NS. In addition, the Kruskal–Wallis analysis at 1 h approached significance (H(3) = 6.70, p = 0.082, n = 5–6/group), and the post hoc comparison showed that 5% HTS had significantly less (p = 0.02) axonal spheroids than the NS group. Interestingly, at 3 h and 4 h post-SCI, the NS and 7.5% HTS groups did not significantly differ, and 3 h post-SCI (H(3) = 10.97, p = 0.012, n = 5–6/group) the 7.5% HTS group approached having significantly more (p = 0.07) axonal spheroids than the 5% HTS group. Next, 4 h post-SCI (H(3) = 15.74, p = 0.001, n = 5–6/group), the 7.5% HTS had significantly more (both p < 0.05) axonal spheroids than both 3% and 5% HTS. Collectively, these results suggest that 3% and 5% HTS treatment reduce axonal spheroid formation, but 7.5% appears to not retain the same protective effects (Fig. 3B). Modeling of axonal spheroid formation after SCI and the effects of HTS treatment are revealed in Figure 4.
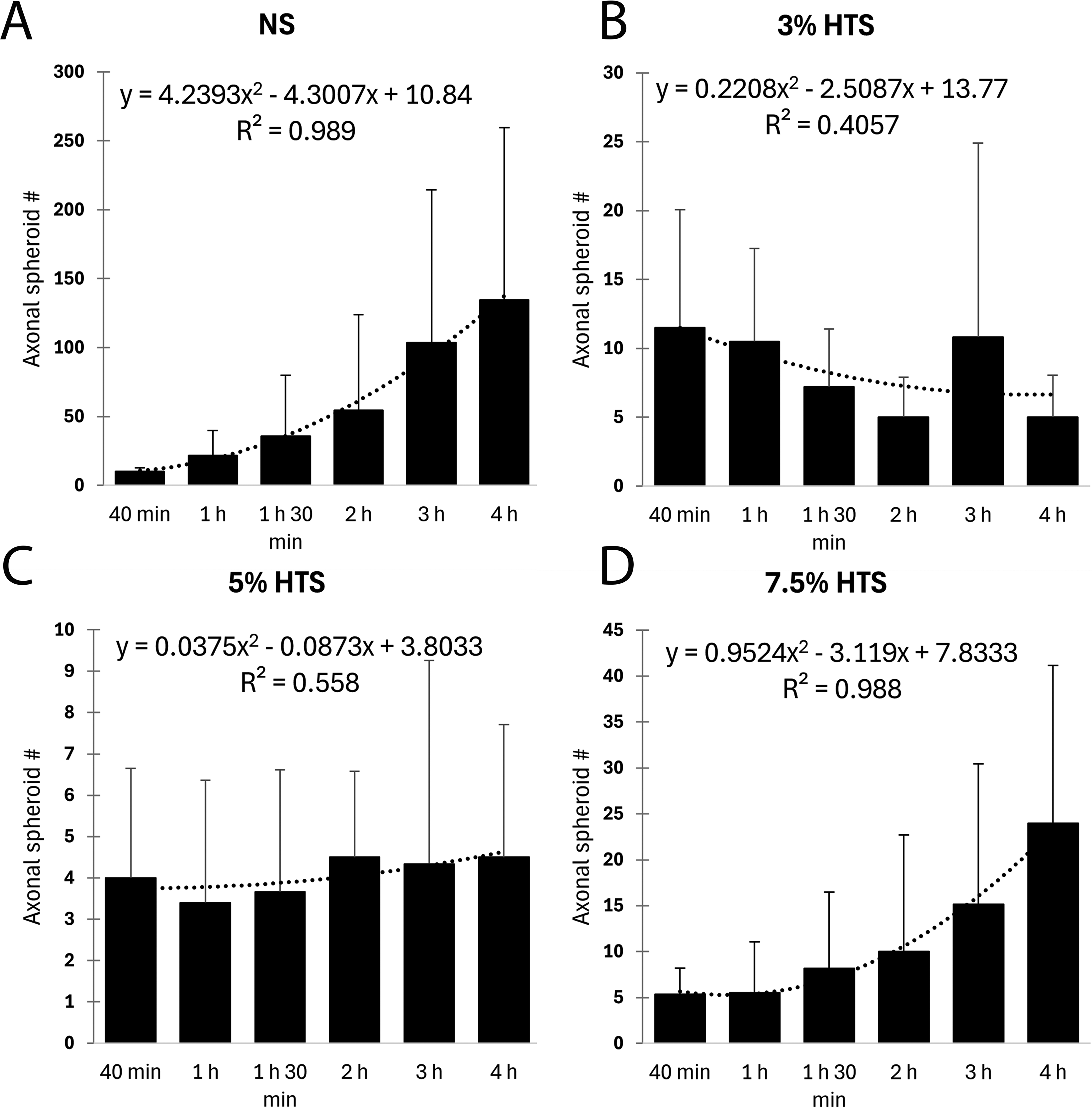
Modeling of axonal spheroid formation after cervical SCI and effects of HTS treatment. Mean axonal spheroid count as a function of time after NS
To determine the efficacy of our 3D-printed dual imaging chamber, we utilized our previously described in vivo imaging method to quantify the effect of delayed treatment (3 h post-SCI) 3% HTS on dorsal column axons post-SCI. 21 Immediately, post-laminectomy Thy1YFP+ mice were imaged via two-photon excitation microscopy to confirm that the laminectomy operation induced no damage to the dorsal column axons and myelin (Fig. 5A, C). Mice then received a 30 kdyn contusion and 3 h post-SCI were intravenously administered either NS or 3% HTS via their lateral tail vein. 24 h post-SCI mice were reimaged at the epicenter of injury, in vivo, with visible WM degeneration (axonal spheroids and periaxonal swellings) present (Fig. 5B, D). Axon survival quantification suggests that single bolus, delayed treatment of low-dose 3% HTS (mean ± SD; 58.09 ± 3.34%) significantly (t-test, t(4) = −6.575, p = 0.003, n = 3/group) protects against secondary WM degeneration 24 h post-SCI compared to NS control (32.080 ± 5.983%; Fig. 5E). Additionally, periaxonal swellings per mm2 are significantly (t(4) = 3.864, p = 0.018, n = 3/group) depressed in the 3% HTS group (595.186 ± 326.100) 24 h post-SCI compared to the NS group (1525.247 ± 259.815). Together these data suggest that our ex vivo 3D-printed dual imaging chamber is a robust model for investigating the influences of osmolarity on axo-myelinic integrity post-SCI ex vivo.
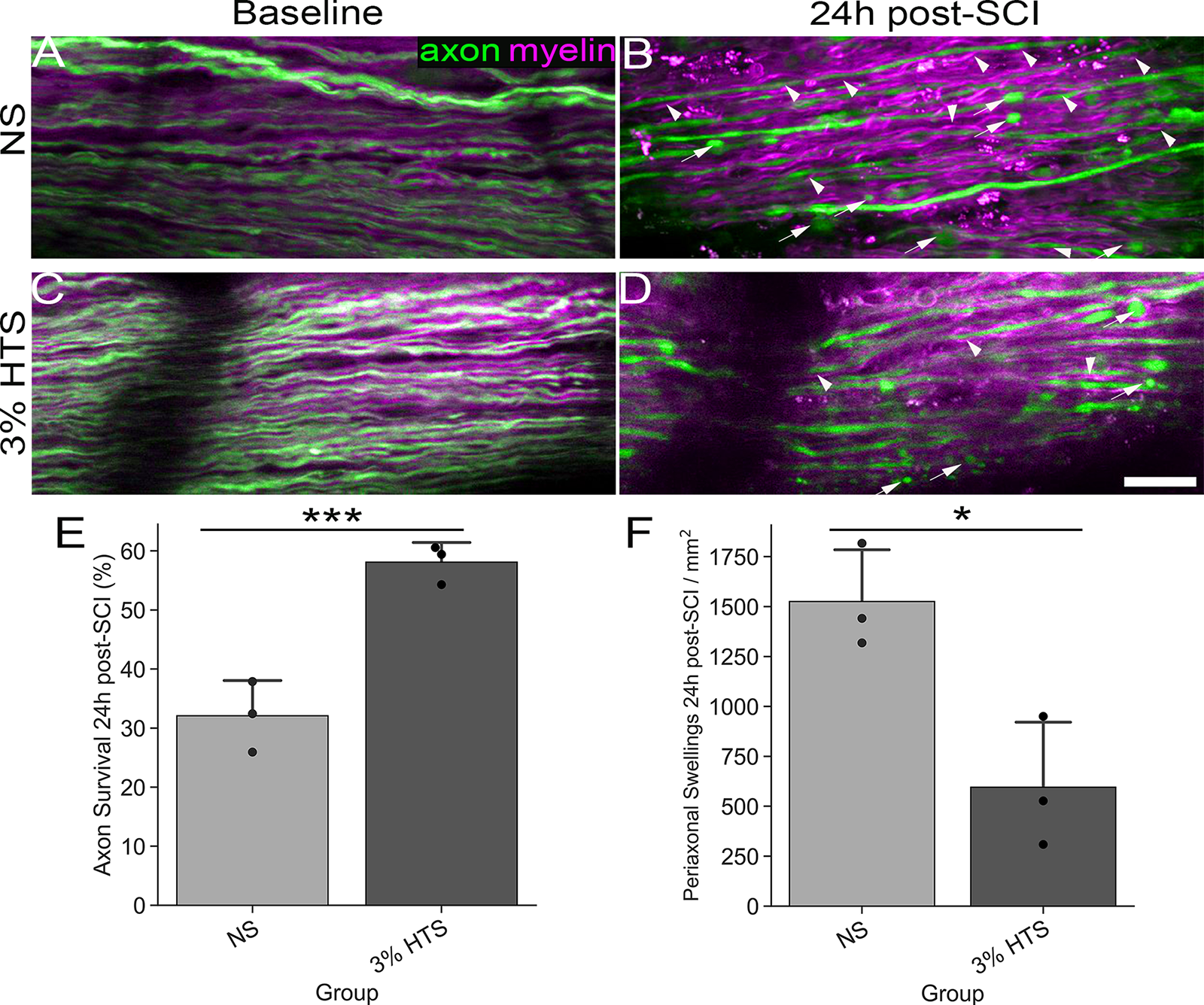
In vivo imaging of HTS-treated mice corroborates the efficacy of the ex vivo dual imaging chamber model. Baseline intravital imaging
Discussion
Our study aimed to investigate the effects of clinically relevant dosages of HTS on periaxonal swelling and axonal spheroid formation following a cervical contusive SCI. Utilizing our 3D-printed dual imaging chamber separated by a semi-permeable membrane, we assessed the effects of continuous perfusion of 3%, 5%, 7.5% HTS and NS applied to the “vascular” chamber to osmotically manipulate the interstitial fluid bathing the cord. Our findings demonstrate that 3% and 5% HTS concentrations significantly mitigate periaxonal swelling and subsequent axonal spheroid formation, suggesting that edema formation after SCI plays a causative role in periaxonal swelling. Furthermore, we have supported the efficacy of our novel ex vivo model by corroborating the protective results of 3% HTS found ex vivo into a clinically relevant in vivo model. Importantly, preventing periaxonal swelling through low-dose HTS treatment also protects axons suggestive of an interrelationship between periaxonal swelling and axonal spheroid formation.
Periaxonal swelling and enlargement of the periaxonal space is one of the most prominent features of WM pathology post-SCI.21–23,26,27 Periaxonal swelling and disruption of the axo-myelinic interface would likely negatively impact neurological recovery by obstructing vital metabolic support between oligodendrocytes and axons and impeding or blocking saltatory conduction along myelinated fibers.28–30 Consistent with previous in vivo reports, our longitudinal imaging data revealed that periaxonal swelling is the most notable morphological change that occurs within myelinated fibers acutely after contusive SCI followed by axonal spheroids that often form within the now enlarged periaxonal space.21,22 Although the precise molecular mechanisms that cause periaxonal swelling remain poorly understood, our current study strongly suggests a role for fluid accumulation in disruption of the axo-myelinic interface. In support, the number of periaxonal swellings and the width of the periaxonal space gradually increase acutely after SCI spatiotemporally coincident with edema formation.7,21 Furthermore, the number of periaxonal swellings was reduced when low dosage HTS was applied to the “vascular” chamber, whereas NS had no effect. Together, these observations support a role of SCI-induced fluid accumulation within myelinated fiber tracts that result in periaxonal swelling and disruption of the axo-myelinic interface.
The mechanisms underlying the observed protective effect of lower concentrations of HTS are likely multifaceted. HTS treatment reduces edema by osmotically drawing water from the interstitial space into the vasculature, thereby reducing fluid accumulation within the former. As tracer studies have shown that dextrans applied to the interstitial fluid penetrate the periaxonal space, albeit at a slower rate,28,31 accumulated fluid within the periaxonal space would likely follow this HTS mediated osmotic gradient back to the vasculature. However, it remains to be determined why fluid would specifically accumulate within the periaxonal space and not between myelinated fibers per se as the gradients between the interstitial space and periaxonal space would be presumably similar.
Although the precise molecular mechanisms remain to be determined, discrete localized ionic dysregulation within the periaxonal region may drive periaxonal swelling. In support, it is well known that ionic dysregulation contributes to WM injury as inhibition of axonal Na channels and Na-Ca exchangers are highly protective and improve conduction.32–35 Indeed, ischemic injury to WM causes depolarization, and pronounced increases in axonal Na that in turn drive reversal of Na-Ca exchanger and resultant conduction failure. Although speculative, reversal of Na-Ca exchange may raise Na within the periaxonal space concomitantly with pathological intra-axonal Ca accumulation that together with periaxonal K accumulation may drive fluid accumulation specifically in the periaxonal space. In support, targeting intra-axonal sources of Ca and store-operated Ca entry is highly protective,25,36 and targeting the latter in part significantly reduces periaxonal swelling and subsequent axonal spheroid formation. 22
In further support of ionic dysregulation occurring within the periaxonal space, we have previously shown that blocking the Na-K-Cl cotransporter NKCC1, a major regulator of Cl levels and cell volume, reduces periaxonal swelling and spheroid formation after thoracic contusive SCI and improves fine aspects of locomotor recovery. 21 Collectively, ionic dysregulation within the periaxonal space could result in periaxonal swelling and drive axonal Ca2+ overload mechanistically linking periaxonal swelling and subsequent axonal spheroid formation.
Interestingly, while the 7.5% HTS group did not show a significant reduction in periaxonal swelling, it exhibited a pattern of axonal spheroid formation like the control group. This suggests that higher concentrations of HTS may not provide additional benefits and might even aggravate certain pathological features. In support, a quadratic fit model for the mean spheroid counts of the control and 7.5% HTS groups indicated a high degree of similarity in their temporal patterns of axonal spheroid formation, with both groups showing a marked increase over time (Fig. 4). We hypothesize that the deleterious effects observed from 7.5% HTS treatment are a result of the excessive magnitude and/or change in osmotic pressure driving water from the axons within the interstitial space into the vasculature compartment disrupting axonal homeostasis and inducing degeneration. In support, hypernatremia has previously been implicated in inducing brain lesions and increased mortality post-TBI.37,38 Additionally, it has been shown that a rapid hypernatremic shift is sufficient to induce brain lesions both in animal models and clinically.39,40 Our data suggest a “U” shaped relation between sodium concentration and markers (axonal spheroids and periaxonal swellings) of secondary degeneration post-SCI wherein we have shown that 3 and 5% HTS are protective compared to NS and 7.5% HTS.
Our findings underscore the therapeutic potential of 3% and 5% HTS in mitigating SCI-induced WM damage. The significant reduction in both periaxonal swelling and axonal spheroid formation in these groups suggests that optimizing HTS concentration is crucial for achieving maximal therapeutic outcomes. Future studies should explore the long-term effects of HTS treatment on functional recovery and neurological outcomes post-SCI in-vivo, as well as further elucidate the molecular mechanisms underlying its protective effects.
Conclusions
Our study provides robust evidence that 3% and 5% HTS effectively mitigate periaxonal swelling and axonal spheroid formation following C5 contusive SCI. Additionally, we corroborate our ex vivo model by confirming the protective results of 3% HTS in vivo. These results enhance our understanding of SCI pathophysiology and offer promising therapeutic strategies for improving outcomes in patients with SCI. Further research is needed to translate these findings into clinical applications and to determine the long-term benefits of HTS treatment in SCI.
Transparency, rigor, and reproducibility statement
All experiments and procedures adhere to a protocol approved by the Institutional Animal Care and Use Committee (IACUC – 22116) at the University of Louisville. Twenty-four adult of 6- to 10-week-old male and female Thy1YFP+ transgenic mice were randomly assigned to treatment groups and controls (n = 6/group). Six number of animals/group was guided by our previous research from our laboratory using an ex vivo SCI model sufficient to detect significant WM pathology after contusive SCI. All image analysis and quantification were done by blinded experimenters. Statistical comparisons between control and treated animals were conducted using nonparametric procedures as normality using the Lilliefors test failed. Statistical significance was set at p < 0.05, unless otherwise specified.
Authors’ Contributions
F.C.-T., S.A., and S.O.A. performed the research. E.J., S.A., and F.C.-T. analyzed data. D.P.S. conceptualized and both D.P.S. and F.C.-T. designed the research. F.C.-T. and D.P.S. wrote the article.
Footnotes
Acknowledgments
The authors thank Christine Armstrong for performing surgery and Darlene Burke for statistical analysis. They also thank Jesse Brooks for technical assistance.
Author Disclosure Statement
The authors declare no competing financial interests to disclose.
Funding Information
This work was supported by Wings for Life-US-14/22 Project# 276.