
Abstract
Under-vehicle blast (UVB) caused by landmine detonation induces a distinct traumatic brain injury (TBI) and can be accompanied by head impact. Injured soldiers often undergo multiple flights after injury to access medical care and return to duty. Previous work has shown that low air pressure during air travel (hypobaria [HB]) exacerbates neurological injury, but the effects of one or more HB exposures on chronic brain injury are unknown. We hypothesized that multiple HB exposures after TBI would result in worse outcomes than 0–1 exposures. Sedated male ferrets underwent UVB and controlled cortical impact (CCI) under anesthesia (BCCI), followed by zero (normobaria [BCCI + NB]), one (BCCI + 1HB), or five (BCCI + 5HB) 6-h HB exposure(s) over 6 months post-injury or remained experimentally naïve. Anxiety-like behavior was assessed with the Open Field at 6 months post-injury. Ferrets also underwent T2-weighted (T2w), resting-state functional magnetic resonance imaging, and diffusion-weighted imaging scans at pre-injury baseline and 6 months post-injury under anesthesia. Relative to naïve, BCCI + 5HB animals expressed significantly more anxiety-like behavior. Region of interest (ROI)-to-ROI functional connectivity (FC) analysis was conducted to evaluate FC changes within the anxiety network. ROI-based diffusion tensor and kurtosis imaging modeling was conducted to evaluate the white matter (WM) integrity of major anxiety-associated WM tracts. Results indicated increased FC between prefrontal cortex, amygdala, and hippocampus and decreased fractional anisotropy and mean kurtosis in cerebral WM, corpus callosum, cingulum, and fornix WM tracts when comparing BCCI + 5HB with all other groups. Together, these suggest that multiple HB exposures after BCCI exacerbate changes in neurological activity in the anxiety regulation brain network, as well as structural damage in the anxiety-associated WM tracts. Our findings demonstrate that air travel after TBI, particularly multiple flights, can have a chronic negative impact on brain structure and function.
Introduction
Traumatic brain injury (TBI) remains a significant concern for military personnel, particularly those exposed to combat environments where the threat of blast exposure is prevalent. 1 Among the various mechanisms of TBI, under-vehicle blasts (UVBs) from landmines and improvised explosive devices represent a distinct and often devastating form of injury, characterized by hyperacceleration forces that can lead to severe neurological damage even without direct impact to the head. However, in addition to exposure to the blast, impact injuries frequently occur with UVB from the head striking the roof or window of the vehicle. TBI from both impact and/or blast exposure can lead to structural, neurochemical, and functional impairments, 2 including mood disturbances such as increased anxiety. 3
Aeromedical evacuation (AE) is a critical component of the medical response system for injured soldiers, facilitating their rapid transport from the battlefield to advanced medical facilities for treatment. After receiving life-saving treatment, soldiers may take additional flights home for rehabilitation and then return to duty. Worse clinical outcomes have been noted in injured personnel who fly soon after injury relative to those who delayed flight, 4 suggesting that patients are particularly sensitive to the effects of air travel in the first 24 h after injury. However, the chronic effects of low air pressure (hypobaria [HB]) experienced during air travel on neurological outcomes after TBI remain poorly understood.
To date, most studies have been performed in rodents, 5 –7 which may be limited in translation to humans by differences in cerebroarchitecture, including a low white:gray matter ratio and a lack of cortical gyrification. A few studies have been performed in swine, 8 –10 which possess brains that more closely resemble humans. However, the size of the animals often limits researchers to more acute time points, possibly due to the high costs of per diem care. In this study, we aim to address this knowledge gap by utilizing a combined model of UVB and impact TBI, followed by 0, 1, or 5 aeromedical evacuation-relevant hypobaria (AERH) exposures over 6 months post-injury. Similar to humans and swine, ferrets are gyrencephalic with an increased white:gray matter ratio. However, unlike swine, an adult male ferret is ≤2kg, making chronic studies with adequate statistical power feasible. Furthermore, ferrets are highly amenable to behavior testing, allowing for the assessment of cognitive function. Based on this, ferrets are a valuable translational model for studying the long-term neurological consequences of TBI and the effects of HB exposure on the injured brain.
Ferrets also fit in a small animal magnetic resonance (MR) scanner, providing researchers with the ability to collect highly translatable information about the structure and function of the brain. The neuroimaging modalities that have been extensively used for assessment following TBI are resting-state functional magnetic resonance imaging (rs-fMRI) and diffusion tensor and kurtosis imaging (DTI/DKI). 11,12 The rs-fMRI measures spontaneous low-frequency oscillations in the blood oxygen level–dependent (BOLD) signal at rest or no task performed, 13,14 allowing for examination of intrinsic brain activity and functional connectivity (FC) patterns, which are often disrupted following TBI. 12,15 –19 These disruptions can affect networks related to emotional regulation, potentially contributing to anxiety and other psychiatric symptoms.
DTI/DKI are tools used to assess the white matter (WM) integrity following TBI. 20 –23 DTI/DKI are particularly sensitive to the direction and magnitude of water diffusion in the brain, which reflects the underlying microstructural organization of WM. Disruptions in normal water diffusion patterns can indicate microstructural damage, such as axonal injury or changes in cellular density, which are key factors in understanding the extent and impact of brain injuries. 24,25 Key metrics derived from DTI/DKI, such as fractional anisotropy (FA), mean diffusivity (MD), and mean kurtosis (MK), are critical for assessing the microstructural integrity and complexity of brain tissues. Following TBI, FA and MK are reduced in major WM tracts including the corpus callosum, internal capsule, and long association fibers compared with healthy controls. 26 –29 Several studies on U2 pilots also showed that non-hypoxic hypobaria exposure is associated with a global decrease in FA and increased WM hyperintensity. 30 Additionally, chronic WM changes are associated with ongoing neurobehavioral symptoms in TBI patients. 31
This study investigated the impact of AERH on anxiety-like behavior and FC alterations between regions within the anxiety network, as well as microstructural changes within WM tracts associated with anxiety. We hypothesized that HB exposure after TBI would worsen chronic behavioral and neuroimaging outcomes, particularly in the case of multiple exposures.
Methods
The study protocol was reviewed and approved by the University of Maryland—Baltimore Institutional Animal Care and Use Committee (protocols 0620009 and 00000048) and the Air Force Medical Readiness Agency (protocol AFOSR-2020-0015A). Animals were handled, and studies were conducted, under a program of animal care accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International and in accordance with the National Research Council’s 2011 Guide for the Care and Use of Laboratory Animals in compliance with Department of Defense Instruction 3216.1. Procedures for the blast and neurobehavioral testing were developed in a pilot study (publication pending).
Ferret BCCI and multiple hypobaria exposures model
A total of n = 40 ferrets (Marshall Bioresources) were maintained on a 12-h light:dark cycle (lights on 0600) with ad libitum access to food and water (n = 10/group). Ferrets were pair-housed prior to injury and then housed individually until sutures were removed, at which time they were reunited with their cage mate. At 4 months post-injury, all animals were housed individually due to an increased incidence of fighting and dominance behaviors noted between the first two animals in the study. They were handled and habituated to the animal behavior suite at least three times prior to the onset of experimental procedures. At approximately 10 weeks of age (∼1000 g), ferrets underwent UVB + controlled cortical impact (CCI) or remained experimentally naïve. Naïve animals underwent identical housing, imaging, and behavioral assessment but did not receive any surgery, pain medication, or HB exposure. A craniotomy-alone control was not used as our previous study noted that craniotomy can still result in a mild brain injury. 2 Only males were used for this study due to ethical concerns with using female ferrets for chronic outcome studies as the unique ferret estrous cycle poses significant health risks to intact females if not bred. 32
Under-vehicle blast
Procedures were based on previous experiments using rats. 5 Ferrets were sedated 15–30 min prior to UVB with 0.05 mg/kg of dexmedetomidine (i.p.; Zoetis) and anesthetized in an induction chamber for 4–5 min with 4% isoflurane in 45% O2 in medical air. Ferrets were then secured in a prone position in a custom polycarbonate cylinder restraint (12.7 × 40.6 cm) atop two 2.5 cm thick aluminum blast plates (40.6 × 38 cm), which were separated by a 6-mm thick rubber pad. Pentaerythritol tetranitrate (PETN 2.5 g) was submerged in a polyurea-coated steel water tank (1.2 × 0.6 × 1.2 m) 5 cm below the surface with a 0.5 cm air gap between the water surface and the bottom plate. PETN was detonated under the center of the blast plates, leading to vertical hyperacceleration of the plates and animal approximately 1 m, guided by poles. Peak acceleration (g) and jerk (m/s3) were recorded for each UVB.
Controlled cortical impact
Immediately following the UVB, ferrets were reanesthetized with 4% isoflurane in 45% O2 in medical air in an induction chamber for up to 5 min and were then placed into a stereotactic frame with ear bars (Kopf) where they were maintained on 1.5–2.0% isoflurane via nose cone. Body temperature was monitored with a rectal thermometer and maintained at 37 ± 0.5°C with a homeothermic blanket (Harvard Apparatus). Blood oxygen saturation (SpO2), heart rate, and respiratory rate were monitored throughout the procedure (Biopac Systems), approximately 1 h. Following aseptic preparation, a midline dorsal incision was made 3–4 cm over the calvarium. The junction of the supraorbital crest was identified and used as a landmark on the skull. A 6–7 mm craniotomy was established with a surgical bone microdrill (Foredom) between the coronal, lambdoid, and sagittal sutures, with the center located 15 mm caudal and 6 mm lateral to the landmark. A CCI device (Leica Biosystems) with a 5 mm beveled piston was used to induce an impact injury with dura rupture at 6.0 mm depth with 6.0 m/s velocity and 50 ms duration. The craniotomy was covered with Surgifoam (Ethicon), secured with dental acrylic, and the incision sutured closed. Atipamezole (i.p., 0.05 mg/kg) was injected after CCI procedures to reverse dexmedetomidine sedation, and buprenorphine (s.c., 0.01–0.05 mg/kg) was administered to all injured animals for analgesia every 8–12 h for 3 days with the first dose administrated prior to surgical manipulation. Animals were monitored twice daily for the first 3 days and then daily until sutures were removed 10–14 days post-injury.
Hypobaria exposure
UVB + CCI (BCCI) animals were assigned to one of three groups: (1) BCCI + normobaria (BCCI + NB) animals remained at near sea level (∼760 mmHg) simulating ground transport for the duration of the experiment, (2) BCCI + 1HB animals underwent 1 AERH exposure (1 day post-injury), or (3) BCCI + 5HB animals underwent 5 AERH exposures (1, 3, 28, 112, and 168 days post-injury). Fully conscious and normally ambulating ferrets were placed inside a portable pet carrier (21.9ʺ × 13.6ʺ × 13.6ʺ) and placed inside a dedicated ferret pressure chamber in preparation for hypobaria exposure as previously described. 2 Each exposure was 6 h long at an atmospheric pressure equivalent to either the cabin of a commercial aircraft at altitude 8000 ft (576 mmHg; HB groups) or to the pressure found near sea level (NB and naïve groups). A mixture of medical air and oxygen (28% O2) was delivered to the chamber during hypobaria to maintain the ambient O2 concentration at levels similar to that at sea level (21%) to avoid hypoxia. Animals were not anesthetized. Food, water, and enrichment objects were provided to animals while in the chamber.
Behavior test
The Open Field test is a popular measure of anxiety-like behavior that has been validated in rodents and applied to other species, including humans. 33 At 6 months post-injury, animals were placed into a custom Open Field arena (122 × 122 × 77 cm) for 10 min. The apparatus was cleaned between animals with 70% ethanol. An activity tracking system (AnyMaze; Stoelting) was used to record and quantify behavior. Time spent in the center zone of the arena was used as a metric of anxiety-like behavior.
MRI data acquisition
All ferrets were subjected to MRI at baseline (no more than 1 week before injury, 8–10 weeks old) and 6-month post-injury (32–34 weeks old). The scans were conducted using a horizontal bore Bruker BioSpec 7T 70/30 MR Scanner (Bruker Biospin MRI GmbH) and a Bruker ParaVision 6.0.1 console. A Bruker 86 mm linear volume coil served as transmitter and a Bruker 1H four-element surface coil array or a rapid circular loop coil (RAPID MR International, LLC) served as receiver. Each animal was injected (i.p.) with dexmedetomidine (0.03 mg/kg) followed by isoflurane anesthesia (4% in 40% O2) in an induction chamber for up to 5 min; 0.3% isoflurane was maintained during the imaging session via nosecone. The respiration rate, SpO2, and body temperature were monitored using a MR-compatible small animal monitoring and gating system (SA Instruments, Inc.). The body temperature was maintained at 37 ± 0.5°C by an MRI-compatible circulating water blanket.
Structural T2-weighted (T2w) images covering the whole brain were obtained using a 2D rapid acquisition with relaxation enhancement pulse sequence along the coronal direction. The imaging parameters are the following: repetition time (TR) = 3200 ms, echo time (TE) = 33 ms, number of averages = 1, flip angle = 90°, field of view = 55 × 55 mm2, in-plane resolution = 0.15 × 0.15 mm2, matrix size = 366 × 366 voxels, slice thickness = 1.5 mm with no gap between slices, and total number of slices = 18.
A total of 400 rs-fMRI image volumes were obtained matching the same spatial coverage as its structural T2w image. A standard 2D single-shot echo planar imaging (EPI) pulse sequence along the coronal direction was used with the following imaging parameters: TR = 1500 ms, TE = 24 ms, in-plane resolution = 0.611 × 0.611 mm2, and matrix size = 90 × 90 voxels. The rest of the imaging parameters matched with those of the corresponding T2w imaging. Additionally, 16 rs-fMRI image volumes were obtained using the same imaging parameters but with an opposite phase encoding direction for geometric distortion correction.
DTI/DKI data were collected using a 2D EPI spin echo diffusion scheme along the coronal axis with 2 b-values (1000 and 2000 s/mm2) and 30 gradient directions for each b-value. Briefly, five b = 0 images were collected, followed by 30 gradient directions for b-value 1 and b-value 2, respectively. This procedure was repeated once, resulting in a total of 130 brain volumes being collected with a total acquisition time of 5 min 25 s. The imaging parameters were TR = 2500 ms, TE = 24.58 ms, in-plane resolution = 0.611 × 0.611 mm2, and matrix size = 90 × 90 voxels. The rest of the imaging parameters matched with those of corresponding T2w imaging. Additionally, 12 brain volumes with b = 0 using the same imaging parameters but with an opposite phase encoding direction were collected for geometric distortion correction.
MRI data processing and analysis
As has been documented in other species, 34 –36 there are neuroanatomical differences by age detectable by MRI in the ferret. Therefore, we processed the data from these two time points separately but using the same procedures.
Study-specific T2w brain template
To register individual T2w, rs-fMRI, and DTI/DKI data into a common space, a study-specific T2w template image was generated following the same method as described by Hutchinson et al. 37 First, all the T2w images from the naïve group were collected for template generation, and the image with the best orientation and quality was chosen as the initial template. The selected T2w images were then bias-corrected using the “N4BiasFieldCorrection” tool from ANTs and brain-extracted using the “BET” tool from FMRIB software library (FSL). The resulting brain masks were visually checked and manually modified using ITK-SNAP tool. Subsequently, the “buildtemplateparallel.sh” script from ANTs was used to create the T2w template based on the T2w brain images and the initial template. The generated T2w templates for the baseline and 6-month post-injury time points are shown in Figure 1.
Constructed study-specific T2w brain templates at
T2w image processing and lesion volume estimation
As above, all the T2w images from all groups were bias-corrected, and brain masks were extracted and manually modified. The resulting T2w images were then registered to the T2w brain template using an affine transformation. By checking the lesion location and boundaries visually, a cuboid is defined to cover the lesion on aligned T2w images of all injured animals. Then, a 2D Otsu image segmentation method and a series of morphological operations (erosion, dilation, identification, and removal of small objects) were applied to extract the brain lesion slice by slice. A manual modification was conducted if necessary. The lesion volume was estimated by the total number of voxels within the extracted lesion mask multiplied by the voxel size. An overall lesion mask was also generated based on the extracted lesion masks from all injured ferrets.
Resting-state fMRI data processing and FC analysis
For the rs-fMRI data, FSL top-up correction was applied to correct the geometric distortion of the brain, and then the standard preprocessing procedures were applied to the rs-fMRI data. The standard preprocessing procedures include the removal of the first 10 brain volumes, slice timing correction, realignment, registration to the T2w template space, and smoothing. Nuisance signals include the average BOLD signal from WM and CSF, and six motion parameters and its first-order derivatives.
ROI-to-ROI FC analysis was applied. The ROIs associated with anxiety included the medial prefrontal cortex (mPFC), dorsal lateral prefrontal cortex (dlPFC), dorsal hippocampus (dHippo), ventral hippocampus (vHippo), and amygdala (see Fig. 2A). The ROIs were manually drawn on the study-specific T2w template using the population-based MRI atlas as reference. 37 Each T2w image was registered and normalized to the study-specific T2w template by applying antsRegistrationSyn.sh with the overall lesion mask. The ROIs of each ferret were obtained by applying the inverse transformation matrix and inverwarp on the ROIs drawn on the study-specific template. The averaged BOLD signal for each ROI was calculated by extracting the BOLD signals from all voxels within the ROI and averaging them. Pearson correlation was calculated between paired averaged BOLD signals from all the ROIs to generate an FC matrix, and all the elements in the FC matrix were z-transformed.
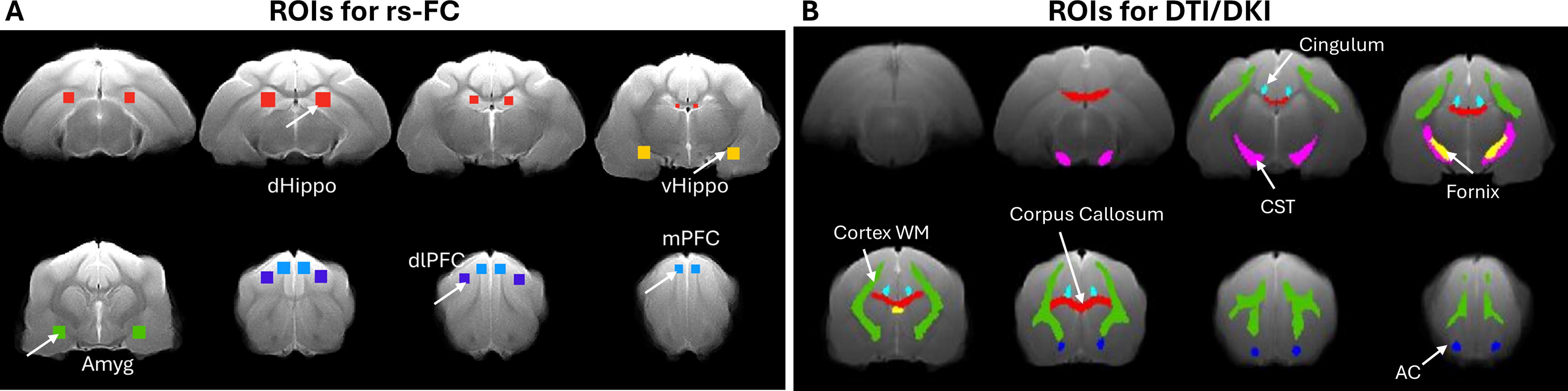
Regions of interest (ROIs) of brain on T2w brain template at 6-month post-injury.
DTI/DKI data modeling and parametric derivation
The DTI/DKI data were first preprocessed using the standard preprocessing pipeline of FMRIB’s Diffusion Toolbox from FMRIB FSL. 38 The preprocessing steps include susceptibility-induced distortion correction, motion correction, and BET for brain extraction on the distortion-corrected B0 image. The brain masks were visually checked and manually corrected using FSLeyes. 39 Then DTI and DKI model fitting was performed using customized diffusion kurtosis software, 27 and metric maps were derived for FA, MD, AD, radial diffusivity (RD), MK, axial kurtosis (AK), and radial kurtosis (RK) in individual space.
The B0 brain images with non-brain tissue removed were rigidly registered to a study-specific T2w template using Advanced Normalization Tools (ANTs) registration with the overall lesion mask. 40 The resulting transformation matrix was then applied to the corresponding metric maps. The registered FA maps were averaged, and the WM regions with higher FA values were extracted. Six WM regions associated with anxiety were identified, manually edited, and used as ROIs. These regions include the cortex WM, corpus callosum, cingulum, fornix, corticospinal tract, and anterior commissure (see Fig. 2B). All ROIs were overlaid on the individual registered FA map and T2w image. The T2w image was used as an anatomical reference, and ROIs were modified if necessary. The values of FA, MD, AD, RD, MK, AK, and RK were extracted from the metric maps within the WM ROIs for each animal. Figure 3 shows representative maps of the estimated DTI/DKI-derived matrices.
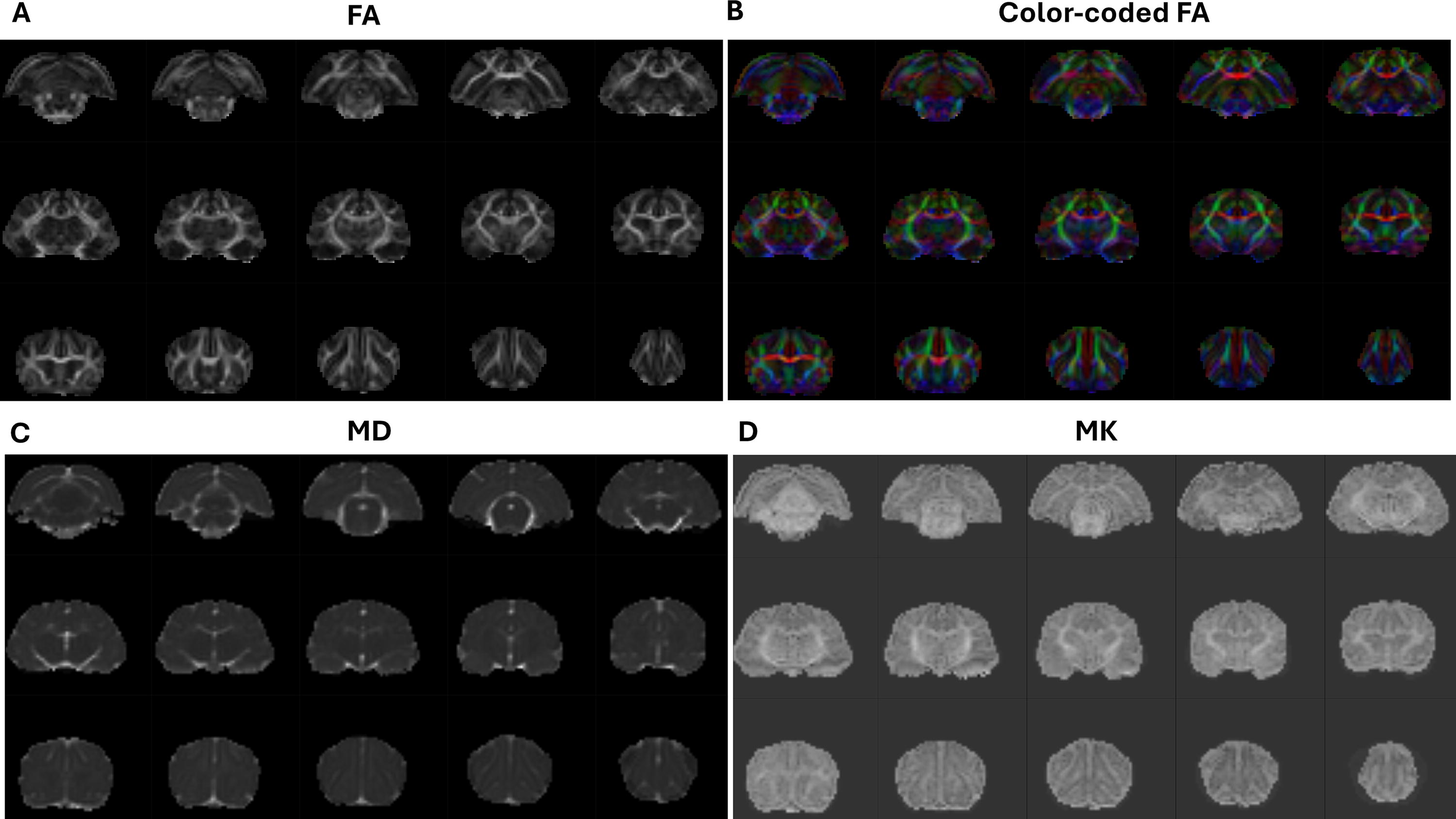
Representative maps of estimated DTI/DKI-derived metrics:
Statistical analysis
Experimenters were blinded to treatment during data collection to the fullest extent possible. Statistical analyses of behavioral data were performed using SigmaPlot 15.0 (Systat). One-way analysis of variance (ANOVA) was used to assess differences between groups. Post-hoc comparisons were made with the Fisher’s Least Significant Difference method. Assumptions of normality and equal variance were assessed with the Shapiro–Wilk and Brown–Forsythe methods prior to data analysis. The Kruskal–Wallis one-way ANOVA on ranks with Dunn’s post-hoc test was used in the event of failed normality tests. Statistical significance was defined as p < 0.05.
Statistical analysis of ROI-to-ROI FC and DTI/DKI-derived metrices (FA, MD, and MK) was performed using one-way ANOVA and Tukey’s post-hoc analysis in IBM SPSS (version 26). Statistical significance was defined as p < 0.05.
Results
Increased anxiety-like behavior in BCCI ferrets after multiple HB exposures
Figure 4 depicts time spent in the center of the arena during the 10-min Open Field test and shows representative track plots of ferret activity in the Open Field apparatus. One-way ANOVA found significant between-group differences in the time spent in the center zone, F(3, 37) = 9.16, p = 0.027. Post-hoc analyses confirmed that, relative to naïve ferrets, BCCI + 5HB ferrets spend significantly less time in the center zone (p = 0.031). No between-group differences were detected at baseline.
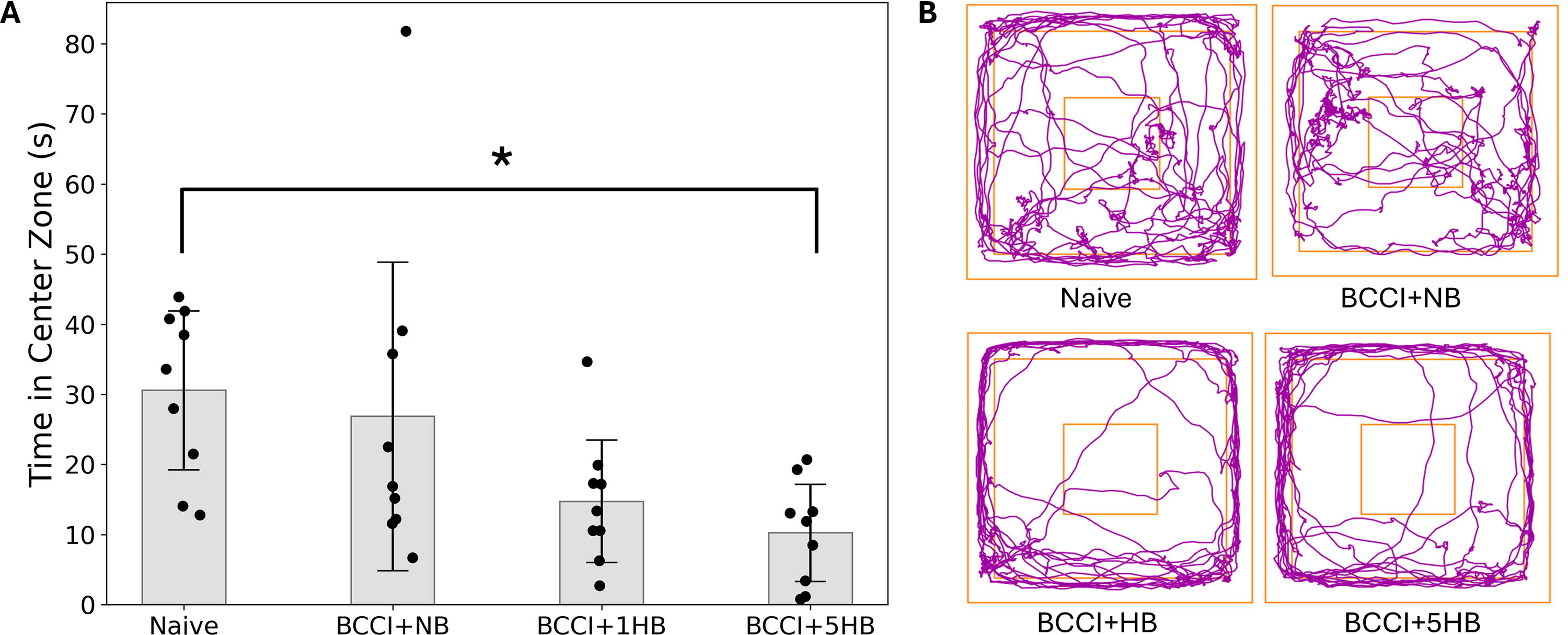
Open Field behavioral test.
No differences in brain lesion volumes among injury groups
An example of a registered injured brain and the delineated lesion is shown in Figure 5A and B. Figure 5C displays the lesion volumes for each ferret group (BCCI + NB, BCCI + 1HB, and BCCI + 5HB) at 6 months post-injury. A one-way ANOVA revealed no significant difference in lesion volume among the three groups, F(3, 27) = 0.86, p = 0.437.
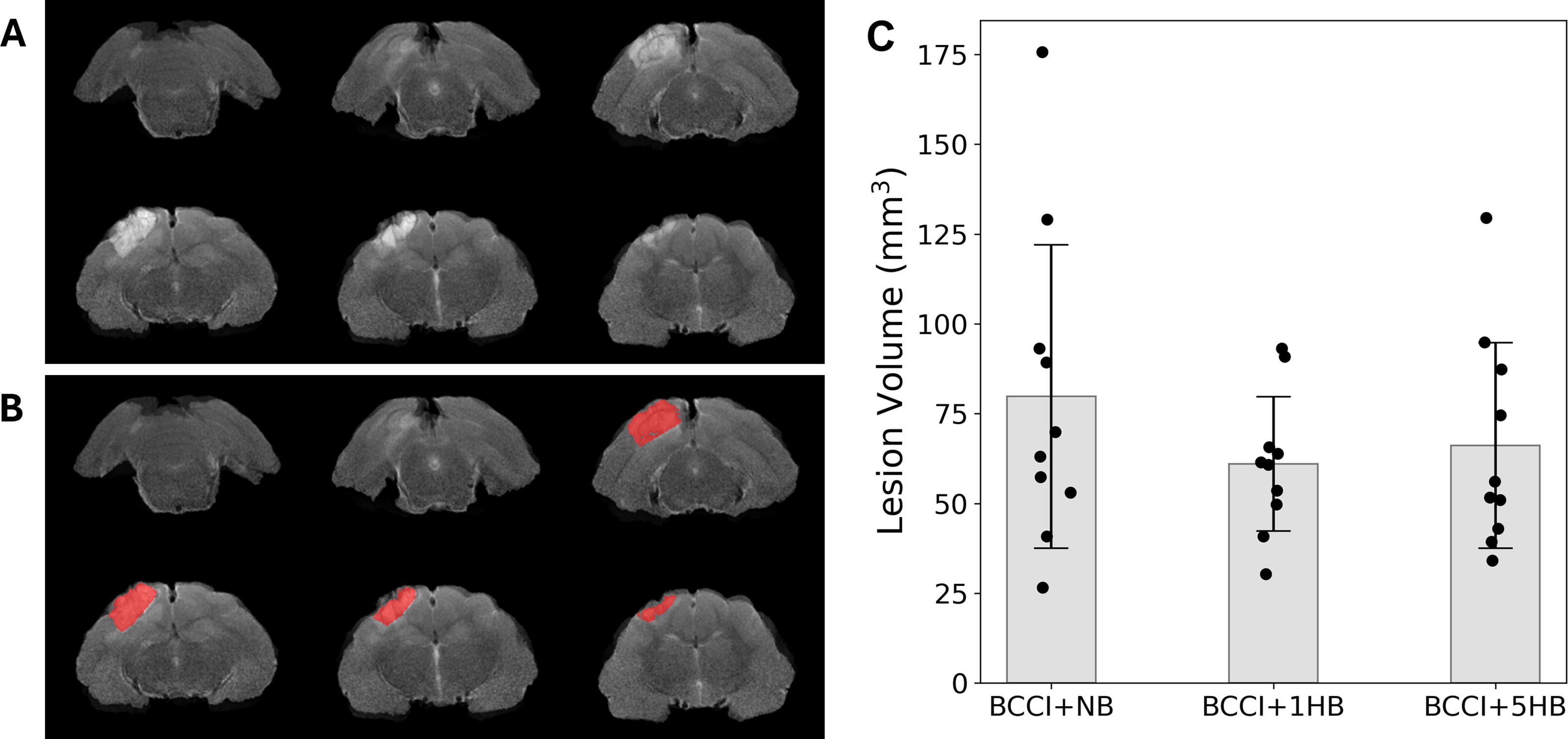
Depiction of lesion mask extraction and lesion volumes at 6-months post-injury (n = 10 per group):
Increased FC between regions within the anxiety network
Figure 6 shows the mean FC matrix for each group (naïve, BCCI + NB, BCCI + 1HB, and BCCI + 5HB) and difference FC matrix between groups. One-way ANOVA found significant differences among groups in FC between mPFC and vHippo (ventral hippocampus), F(3, 37) = 3.52, p = 0.025, and between mPFC and amygdala, F(3, 37) = 3.60, p = 0.023. Post-hoc analysis showed that when compared with naïve group, BCCI + 5HB showed a significant increased FC between mPFC and vHippo (p = 0.013) and between mPFC and amygdala (p = 0.018).
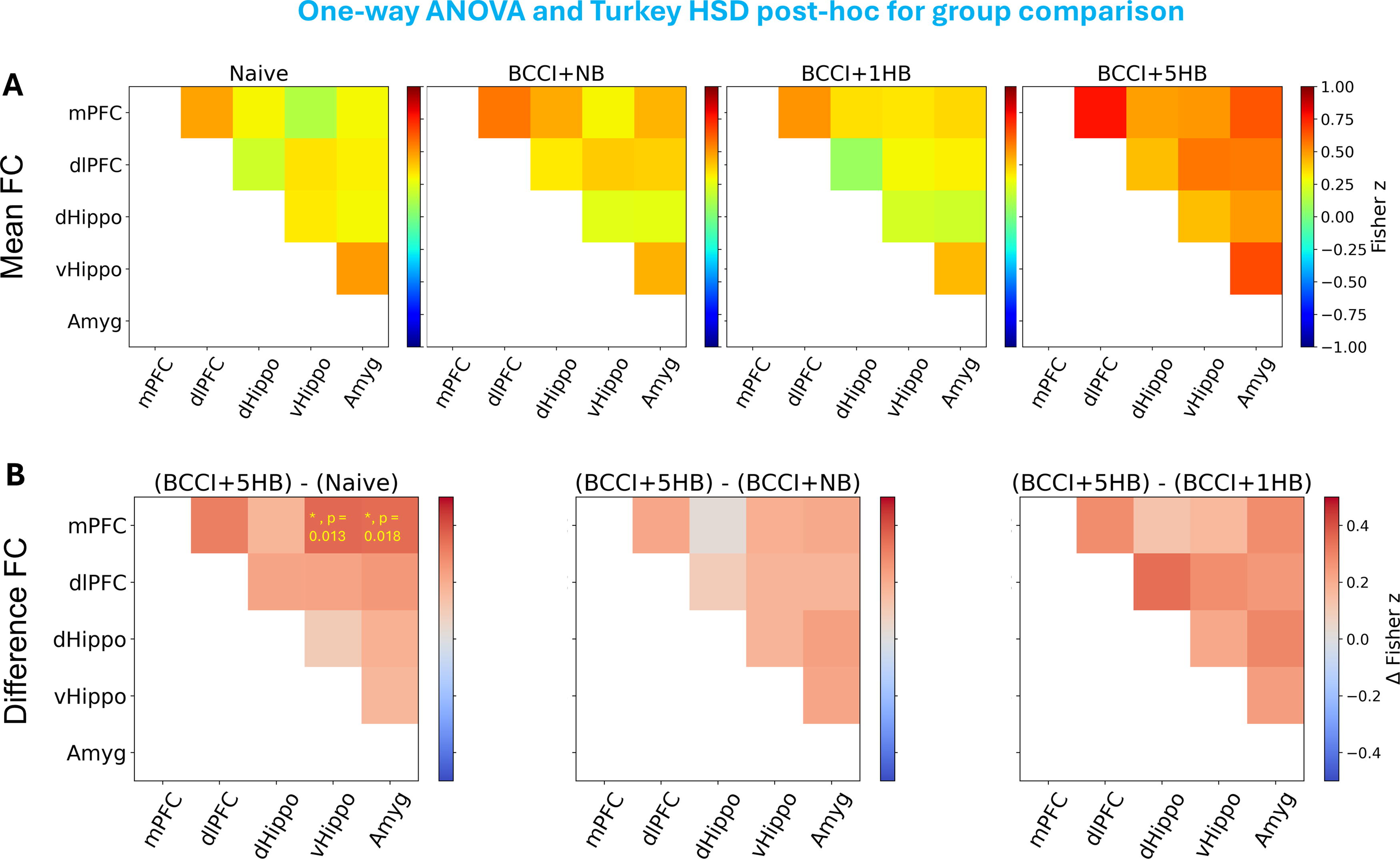
ROI-to-ROI FC analysis results.
Reduced WM integrity in WM tracts associated with anxiety
Fractional anisotropy
A one-way ANOVA was conducted to evaluate the relationship between treatment groups (naïve, BCCI + NB, BCCI + 1HB, and BCCI + 5HB) and FA in major WM tracts associated with anxiety (Fig. 7). The ANOVA results indicated significant differences in FA for the following tracts: corpus callosum, F(3, 37) = 9.00, p < 0.001; cingulum, F(3, 37) = 16.63, p < 0.001; and cortical WM, F(3, 37) = 4.84, p = 0.006. Tukey’s post-hoc analysis indicated the following significant differences:
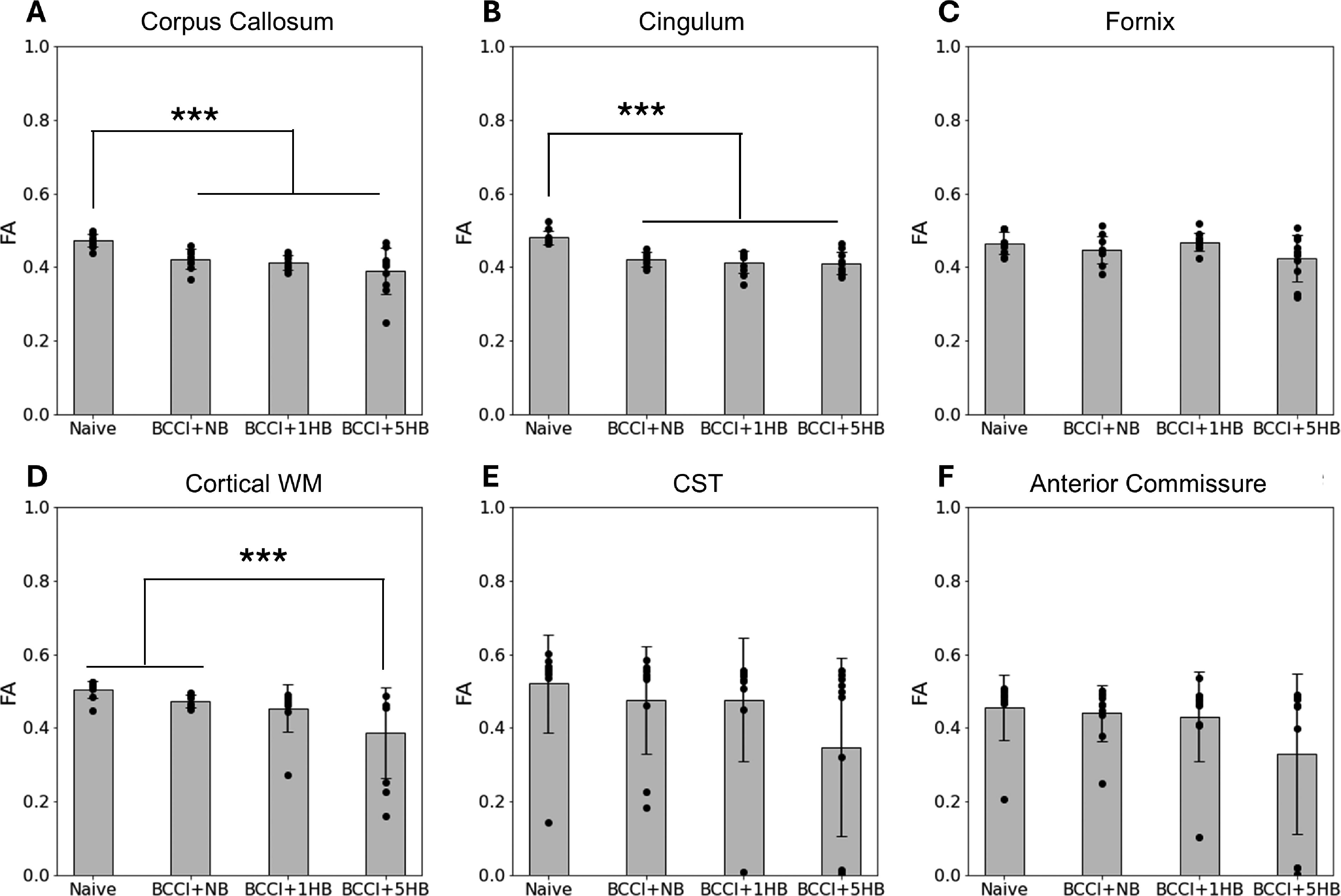
One-way analysis of variance and Tukey’s post-hoc results of FA for six white matter tracts associated with anxiety:
Corpus callosum: Compared with naïve group, significantly lower FA was found in the BCCI + NB group (p = 0.021), the BCCI + 1HB group (p = 0.004), and the BCCI + 5HB group (p = 0.0001).
Cingulum: Relative to naïve, FA was significantly reduced in the BCCI + NB group (p < 0.0001), the BCCI + 1HB group (p < 0.0001), and the BCCI + 5HB (p < 0.001).
Cortical WM: The BCCI + 5HB group had significantly lower FA than the naïve group (p = 0.004) and the BCCI + NB group (p = 0.049).
Mean diffusivity
MD data are presented in Supplementary Figure S1. The ANOVA results showed no significant differences at the p < 0.05 level for all six WM tracts.
Mean kurtosis
MK data are presented in Figure 8. The ANOVA results indicated significant differences for cingulum, F(3, 37) = 9.65, p < 0.001, and cortical WM tract, F(3, 37) = 4.07, p = 0.013.
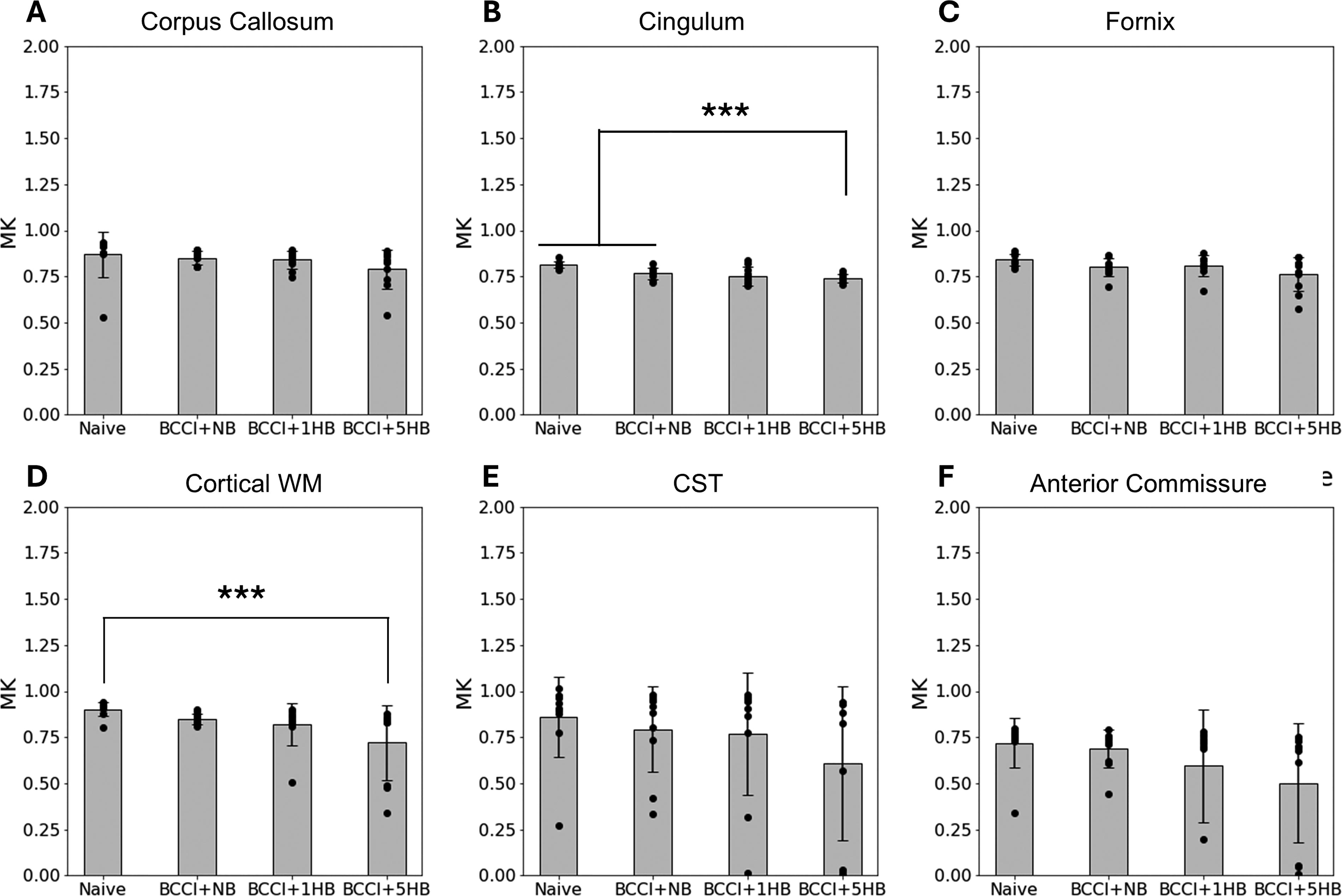
One-way analysis of variance and Tukey’s post-hoc results of MK for six white matter tracts associated with anxiety:
Tukey’s post-hoc analysis indicated the following significant differences: Cingulum: Relative to naïve group, MK was significantly reduced in the BCCI + NB group (p = 0.008), the BCCI + 1HB group (p = 0.001), and the BCCI + 5HB group (p = 0.0001). Cortex WM: Compared with naïve group, only the BCCI + 5HB group had significantly lower MK than naïve group (p = 0.0088).
Discussion
This is the first MRI study to correlate FC and WM integrity changes with long-term neurobehavioral outcomes in a ferret TBI model. Our behavioral results confirmed that multiple exposures to AERH after BCCI induced anxiety-like behavior in ferrets. Additionally, MRI studies revealed that multiple HB exposures also increased FC between regions within the anxiety network and reduced integrity of WM tracts associated with anxiety.
Previous research has demonstrated that brain injury contributes to neurobehavioral disturbances in ferrets, though research is limited. 2,41,42 We found that ferrets exposed to blast exhibit longitudinal biochemical and behavioral alterations, specifically with heightened impulsivity in decision making associated with prefrontal cortex (PFC)/amygdala axis dysfunction. 42 Impact injuries are associated with disruptions in memory. 2 TBI-induced anxiety-like behavior has not previously been described in ferrets; this may be due to the timing of testing or methodology (e.g., repeated vs. single exposure to the testing apparatus). However, our results indicate that the combination of injury and multiple HB exposures was necessary to result in statistically significant anxiety-like behavior. Studies in rats had similar findings. 7
Our rs-fMRI analysis revealed significant increases in FC within the anxiety brain network in the BCCI ferret model subjected to multiple HB exposures. The increased FC in regions associated with anxiety, including PFC, hippocampus, and amygdala, suggests heightened synchrony and possibly hyperactivity within these regions. For BCCI + 5HB group, we observed increased amygdala–PFC FC. The amygdala is crucial for processing emotional responses, particularly fear and anxiety. The PFC, especially the medial PFC, plays a vital role in regulating these responses and exerting top-down control over the amygdala. Increased amygdala–PFC connectivity has been reported in individuals with anxiety disorders, including generalized anxiety disorder, social anxiety disorder, and post-traumatic stress disorder (PTSD). 43 –45 This hyperconnectivity is often associated with impaired regulation of emotional responses. Our findings are consistent with these studies, suggesting that similar neural mechanisms may underlie anxiety across different species.
Our DTI/DKI analysis revealed significant decreases in FA and MK following BCCI and multiple AERH exposures within key WM tracts associated with anxiety, including the cortical WM, corpus callosum, and cingulum. These findings provide valuable insights into the microstructural alterations in WM that accompany anxiety in this model. FA is a measure of the directional coherence of water diffusion in WM tracts, reflecting the integrity of WM fibers. Decreased FA in the cortical WM, corpus callosum, and cingulum suggests a reduction in fiber density, myelination, or coherence, indicating WM damage or degeneration associated with anxiety. MK measures the complexity of water diffusion, providing additional information about tissue microstructure beyond what FA offers. Reduced MK indicates a loss of microstructural complexity and organization in these WM tracts, which may be due to demyelination, axonal loss, or other structural disruptions. Decreased FA and MK in anxiety-associated WM tracts have been reported in individuals with anxiety disorders, PTSD, and other stress-related conditions, 46 –48 as well as in TBI patients 49,50 These findings often point to WM integrity disruption as a key factor in anxiety pathophysiology. Our findings are consistent with these studies, supporting the idea that similar microstructural changes occur across species and different models of anxiety.
An additional interesting finding is that we observed WM integrity (FA and MK) changes not only in BCCI + 5HB group but also in BCCI + NB and BCCI + 1HB groups. It may suggest that the WM changes likely reflect a direct consequence of the initial BCCI injury and are then progressively exacerbated by increasing exposures to hypobaria, as evidenced by the more pronounced FA and MK changes in the BCCI + 5HB group. However, these structural changes did not uniformly translate into FC changes or anxiety-like behavioral changes. Only the BCCI + 5HB group exhibited significant FC changes between the mPFC and the ventral hippocampus, as well as between the mPFC and the amygdala. These FC alterations corresponded to significant anxiety-like behavior observed in the open test in this group, implicating FC changes within key emotion-regulation circuits as a driver of these behavioral changes. In contrast, the BCCI + NB and BCCI + 1HB groups, despite exhibiting WM integrity change, did not show significant FC changes or anxiety-like behaviors, potentially reflecting compensatory adaptations preserving functional integrity. The mechanisms that underlie these HB-induced structural and functional changes are unknown. While we controlled for ambient hypoxia in this experiment, it is possible that the changes in air pressure induced WM hypoxia, perhaps due to decreased blood flow. 9 Hypobaria has also been linked to increased markers of inflammation and oxidative stress, both within the brain and in the periphery, 6 –8,10,51,52 suggesting that immunomodulation or antioxidants may mitigate some of the effects of HB on the injured brain. In particular, as oligodendrocytes are especially sensitive to oxidative stress due to their high metabolic demand and limited antioxidant capacity, 53 agents that protect oligodendrocytes or promote proliferation/differentiation of oligodendrocyte progenitor cells may be particularly beneficial in TBI patients exposed to hypobaria.
Limitations
While the ferret model provides valuable insights, conclusions are limited by several factors. Of primary concern is the use of a single sex (males). Sexually dimorphic outcomes have been noted in humans and other animal models after TBI, 54 so results may not be translatable to females. In addition to sex, age is an important factor in TBI pathophysiology. Although the ferrets in this study are sexually mature at the time outcomes were measured, they were approximately 10 weeks of age at the time of injury, corresponding developmentally to a human adolescent. It is unknown if injuries incurred at other ages would be similarly sensitive to HB. The model is also highly specific; future studies should investigate other injury models (e.g., closed head, blast overpressure, repeated low-level blast) to determine if similar effects of HB are seen. Previous work suggests that HB exacerbates brain injury in other models 5 –7,55 though the effects on anxiety circuits are unknown. Furthermore, the lack of a naïve + HB control group does somewhat limit our ability to make conclusions about the effects of HB on the healthy brain. It is possible that the effects of HB in the BCCI animals may not be a synergistic effect of injury + HB and could simply be due to the HB alone. However, this seems unlikely with flight-relevant HB 9,56 ; indeed, rates of anxiety disorders are similar between commercial pilots and the general population. 57 Finally, animals were not monitored for hypotension or hypercarbia while under anesthesia for surgery and imaging. It is possible that BCCI animals were more sensitive to perturbations in brain perfusion or acidosis. 2 Despite this, analyses noted greater neurological injury in BCCI + 5HB animals; these animals underwent identical anesthesia procedures as BCCI + NB and BCCI + 1HB groups yet were not as profoundly affected. This suggests that HB rather than sedation/anesthesia is likely responsible for these effects.
Conclusion
In conclusion, our study demonstrates that exposure to AERH, particularly multiple exposures, can lead to anxiety-like behavior, increased FC in anxiety brain networks, and decreased FA and MK in key anxiety-associated WM tracts after combined UVB and CCI in male ferrets. These findings highlight the importance of FC and WM integrity in anxiety regulation and underscore the potential of FA and MK as TBI biomarkers and therapeutic targets. Results indicate that air travel soon after TBI should be avoided or minimized whenever possible. This study will inform guidelines for the safe transport of injured military personnel as well as civilians who may travel after TBI, including college athletes.
Transparency, Reproducibility, and Rigor
The study design was not preregistered. A sample size of n = 10 subjects per group was planned to yield 80% power to detect a statistically significant difference in lesion volume between injured and naïve animals with a p value <0.05. Ferret shipments were assigned to treatment group prior to arrival. Behavior testing occurred between 12:00 and 15:00; imaging was performed between 09:00 and 13:00 daily. All animals survived until the end of the experiment. Imaging acquisition and analyses were performed by team members blinded to treatment group, though the presence of a CCI lesion was clearly visible on scans. Open Field results were quantified automatically by AnyMaze software. Imaging equipment and software are available from commercial sources. Statistical analyses were performed as described in the section “Methods.” Data from this study are available upon reasonable request and with approval from the U.S. Air Force (USAF).
Footnotes
Acknowledgments
The authors thank Vayda Ballard for expert animal husbandry. They also appreciate the administrative support provided by Michele Robinson, Laura Kaddis, and Todd Noe. They also thank the University of Maryland School of Medicine Center for Innovative Biomedical Resources, Center for Translational Research in Imaging, for performing neuroimaging procedures. Portions of this article came from a preprint source and abstracts from the 2024 Annual Meeting of the National Neurotrauma Society.
Authors’ Contributions
L.J.: Methodology, formal analysis, writing–original draft, writing–review and editing, and visualization. M.J.G.: Methodology, formal analysis, investigation, writing–original draft, writing–review and editing, and supervision. S.X.: Methodology, writing–review and editing, and supervision. B.P.: Investigation. J.L.P.: Conceptualization, writing–review and editing, project administration, and supervision. X. Liang, X. Li, and S.R.: Methodology. R.P.G.: Conceptualization. A.H., U.H.L., and W.F.: Methodology and investigation. J.C.C.: Project administration. C.H.T.M.: Funding acquisition. J.Z.: Writing–review and editing and supervision. G.F.: Project administration and funding acquisition.
Disclaimer
The views expressed are those of the authors and do not reflect the official guidance or position of the U.S. Government, the Department of Defense, or of the USAF. Imagery in this document is property of the USAF, AFRL-2025-0061, cleared January 16, 2025.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study is supported by USAF FA8650-21-2-6L01.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.