
Abstract
In this study, we have investigated the possibility of combining a cyclodextrin-containing polymer (CDP) with siRNA molecules to modulate gene expression in a light-directed manner through photochemical internalization (PCI) technology. We utilized S100A4 as a model gene to evaluate the efficacy of gene silencing. After optimization of carrier/cargo ratio and illumination dose, real-time reverse transcriptase–polymerase chain reaction data showed between 80% and 90% silencing in the siRNA samples treated with PCI compared with untreated control. In contrast, only a 0%–10% silencing effect was detected in the siRNA samples without PCI treatment, demonstrating the potency of light-specific delivery of siRNA molecules. Light-directed siRNA delivery was shown in 2 different cell lines with corresponding potency. Further, time-lapse results demonstrated maximum gene silencing only at 5 hours after endosomal release, implying, for example, rapid carrier decondensation when using the CDP. This work represents a first success in using a CDP delivery agent, without endosomolytic properties for siRNA gene silencing in a light-directed manner, opening the opportunity to use CDPs for light-directed siRNA gene silencing in vivo.
Introduction
Cyclodextrin (CD)-containing polymers (CDP) consist of cyclic oligomers of glucose where moieties of 6, 7, and 8 glucose units are termed alpha (α)-, beta (β)-, and gamma (γ)-CDs, respectively. Of importance, CDs are known to lack immunogenicity and were approved by the Food and Drug Administration (Szejtli and Osa, 1996). Amongst a class of novel linear, cationic, β-cyclodextrin-containing polymers (β-CDPs) with 4, 5, 6, 7, 8, and 10 methylene units, the β-CDP based on 6 methylene units (β-6CDP) has been found effective for nucleic acids delivery (Gonzalez et al., 1999). However, in contrast to polyethylenimine (PEI), CDPs do not contain any pH-buffering properties and are therefore unable to escape the endosomal pathway by themselves (Gonzalez et al., 1999); Midoux and Monsigny, 1999; Kulkarni et al., 2005). To overcome endosomal and lysosomal entrapment and subsequent degradation, β-6CDPs have been modified with imidazole groups for improved intracellular delivery (Davis et al., 2004). The imidazole-containing variant (CDPim) has been shown to buffer the pH inside endosomal vesicles, resulting in increased intracellular delivery compared with unmodified β-6CDPs (Kulkarni et al., 2005). An alternative strategy for overcoming entrapment in the endosomal pathway without modifying the CDP is by the use of photochemical internalization (PCI) technology (Berg et al., 1999). The main reason for using PCI compared with the use of CDPim is the possibility for light-directed delivery of siRNA molecules into the tissue of interest.
PCI technology depends on photosensitizers (PSs) that localizes preferentially to endosomal and lysosomal membranes, such as disulfonated tetraphenylporphine (TPPS2a) and AlPcS2 (Berg and MOAN, 1994; Moan et al., 1994). Illumination of these PSs initiates photochemical membrane damage followed by release of endosomal and lysosomal content. The potential of PCI technology for in vivo applications have been previously documented on tumor treatment with bleomycin (Berg et al., 2005), the protein toxin gelonin (Selbo et al., 2001), and a plasmid encoding a therapeutic gene (Prasmickaite et al., 2000; Ndoye et al., 2006). Further, the PCI strategy has been employed for siRNA molecules in vitro with carriers such as Lipofectamine™ (LF) 2000 (Oliveira et al., 2007), jetSI/jetSI-ENDO (Boe et al., 2007), and linear and branched PEI (B-PEI) with various molecular weights (Boe et al., 2008) and in vivo using LF 2000 (Oliveira et al., 2008). Common for these reported siRNA results is the use of a carrier with intrinsic endosomolytic capacity (ie, lipid fusion or pH buffering). There are at least 2 obvious disadvantages when using these carriers: first, their nature may be less suited for in vivo applications because of immunogenicity and cytotoxicity. Second, the intrinsic endosomolytic capacity may naturally result in a more severe gene silencing effect also in samples (areas) not treated with PCI compared with the use of nonendosomolytic carriers. The latter has been previously demonstrated by investigating various PEI formulations under different nitrogen/phosphate (N/P) ratios (Boe et al., 2007, 2008).
For the PCI-induced siRNA gene silencing, we chose the human S100A4 as a model gene. The S100A4 protein is a small (11.5 kDa) calcium-binding protein previously linked to the invasive and metastatic phenotype of cancer cells (Davies et al., 1993; Maelandsmo et al., 1996; Takenaga et al., 1997). In this study, we demonstrate the possibility to use an unmodified β-6CDP for light-directed siRNA gene silencing. Second, we demonstrate the opportunity for light-directed siRNA delivery using a carrier without endosomolytic properties. Third, we show the actual amount of time needed from endosomal escape to potent mRNA degradation through the RNAi mechanism by β-6CDP-mediated siRNA delivery. In addition, we have here directly compared the performance for carriers previously tested for light-directed gene silencing with β-6CDP. In conclusion, the properties of β-6CDP are favorable compared with previous light-directed siRNA protocols by offering rapid and enhanced targeting specificity and reduced long-term accumulation in vivo due to the biocompability of CDP.
Materials and Methods
Cell lines and culture conditions
The OHS (osteosarcoma) and the RMS (melanoma) cell lines were established at the Norwegian Radium Hospital (Fodstad et al., 1986). Cells were cultured and maintained in RPMI 1640 medium (Bio Whittaker). The medium was supplemented with 10% fetal bovine serum (PAA Laboratories) and 2 mM
siRNA design
siRNA molecules were selected against the S100A4 mRNA sequence (GenBank accession no. NM_002961). A specific siRNA (481–499; sequence 5′-UGAGCAAGUUCAAUAAAGA-3′, 3′-ACUCGUUCAAGUUAUUUCU-5′) was designed from the rational siRNA design program (Reynolds et al., 2004). Additionally, a scrambled control version (5′-CGCAUAAGUGAAAUAGAAU-3′, 3′-GCGUAUUCACUUUAUCUUA-5′) was designed on the basis of the specific siRNA sequence. The GC content of the duplexes was kept within the 30%–70% range, and all siRNA molecules were synthesized with dTdT overhangs at their 3′ ends for optimal stability of the siRNA duplexes. siRNA molecules were obtained from Eurogentec. siRNA sequences were also aligned to the human genome database in a BLAST search to eliminate those with significant homology to other genes before ordering. Dried siRNA oligonucleotides were resuspended to 100 μM in diethylpyrocarbonate (DEPC)-treated water and stored at −20°C. Annealing was performed by separately aliquoting and diluting each RNA oligonucleotide to a concentration of 50 μM. Then, 30 μL of each RNA oligo solution and 15 μL of 5 × annealing buffer were combined and made to a final concentration of 50 mM Tris (pH 7.5) and 100 mM NaCl in DEPC-treated water. The solution was then incubated for 3 minutes in a water bath at 95°C, followed by gradual cooling for 45 minutes on the workbench. Successful annealing was confirmed by 4% NuSieve agarose gel electrophoresis (data not shown).
siRNA transfection
The β-6CDP formulation was synthesized as described previously (Gonzalez et al., 1999). B-PEI (25 kDa), LF 2000, and jetSI™ were obtained from Sigma, Invitrogen, and Polyplus transfection, respectively. β-6CDP and B-PEI were diluted in sterile water before use. All complex formations were performed in serum-free medium. The complexion solution was added to serum-containing transfection solution after 30 minutes for β-6CDP and B-PEI, whereas the manufacturer's protocol was followed for both LF 2000 and jetSI. Before transfection, cell lines were cultured as described in the Cell lines and culture conditions section and plated in 12-well plates before transfection. Cells were transfected with complexes with or without PS (TPPS2a = 0.5 (g/mL), incubated for 18 hours, then washed 3 times with fresh medium, and reincubated for 4 hours prior to light treatment. The cells were exposed to blue light (7 mW/cm2) and reincubated before harvesting at various time points (0–48 hours), with 24 hours as the default incubation time. To measure the effect of PCI on gene silencing, the untreated cells, scrambled siRNA/β-6CDP, and S100A4 siRNA/β-6CDP were combined in different wells of the same plate, with or without PS, and given the exact same treatment. Cells were light protected by aluminum foil during the experiments. siRNA and carrier concentrations, together with illumination doses, are described under each experiment in the Results section because of many different parameters tested.
Light source and PS
We used the same light source and PS as described previously (Boe and Hovig, 2006). Briefly, TPPS2a was purchased from Porphyrin Products. When irradiated, cells were exposed to light with a LumiSource prototype (PCI Biotech AS) containing a bank of 4 fluorescent tubes (Osram 18W/67) with the highest fluorescence around 420 nm.
Cell viability study
OHS cells were plated in 12-well plates and allowed to adhere overnight in serum-containing medium. Cells were then transfected with 0.7 μg siRNA combined with 25 μg β-6CDP, with or without PS (0.5 (g/mL). Untreated cells and cells transfected with scrambled siRNA/β-6CDP complexes were used as controls. After transfection for 18 hours, cells were washed 3 times with fresh medium and reincubated for 4 hours prior to light treatment. Cells were then exposed to blue light (7 mW/cm2) at 3 different irradiation doses (140, 280, and 420 J/cm2). Twenty-two hours after irradiation, 200 (L of MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) solution (Promega) was added to each well, making a total volume of 1200 (L (1:6 dilution). The plates were then reincubated for another 2 hours before absorbance was measured at 24 hours after irradiation at 490 nm. Cells were light protected by aluminum foil during the experiments.
Zeta potential of β-6CDP/siRNA complexes
S100A4 siRNA and β-6CDP were combined in serum-free medium or sterile water to form complexes under different weight ratios. Zeta potential was measured using a Zetasizer Nano ZS (Malvern Instruments Ltd.).
Real-time reverse transcriptase–polymerase chain reaction of S100A4
Total cellular RNA was isolated with the GenElute Mammalian Total RNA Miniprep Kit (Sigma-Aldrich) and the iScript cDNA synthesis kit (BioRad) was used for reverse transcription. Both kits were used according to the manufacturers' manuals. All polymerase chain reactions (PCRs) were run in parallels. Real-time detection was obtained by use of SYBR Green I. For each PCR, 10 μL cDNA, 30 μL iQ SYBRGreen Supermix (BioRad), 300 nM of each primer, and nuclease-free water were added to a final volume of 60 μL. Samples of 25 μL each were then applied to the PCR plate. This method ensures that the parallels are true ones, and that there is enough PCR mix for all replicates. Primer design was performed using the software Primer Express (Applied Biosystems). The primer set used (forward primer 5′-AAGTTCAAGCTCAACAAGTCAGAAC-3′ and reverse primer 5′-CATCTGTCCTTTTCCCCAAGA-3′) amplifies a 79-bp segment covering parts of exons 2 and 3 of the S100A4 sequence. Real-time reactions were run on an iCycler (Bio-Rad) with the following amplification protocol: 3 minutes initial denaturation at 95°C, 50 cycles of 10 seconds denaturation at 95°C and 35 seconds annealing/extension at 60°C, a hold at 95°C for 20 seconds followed by a hold for 1 minute at 55°C, and finally, a melt curve analysis of 80 steps each for 10 seconds, with 0.5°C increase until a final temperature of 95°C. The quality of the RNA samples was verified by amplification of 2 housekeeping genes: the TATA-binding protein with forward primer 5′-GCCCGAAACGCCGAATAT-3′ and reverse primer 5′-CGTGGCTCTCTTATCCTCATGA-3′ and the human acidic ribosomal phosphoprotein PO with forward primer 5′-CGCTGCTGAACATGCTCAAC-3′ and reverse primer 5′-TCGAACACCTGCTGGATGAC-3′. The Gene Expression Macro, version 1.1 (Bio-Rad), was used for quantitative calculations. The program performs calculations based on the ΔΔCT method (Vandesompele et al., 2002), which allows comparison of cycle threshold values obtained using different sets of primers on the same set of samples.
Results
Optimization of β-6CDP and siRNA concentrations upon light-directed gene silencing
The S100A4 gene was chosen as a model gene to evaluate the efficacy of gene silencing with or without PCI treatment. We initiated our studies with the intention to investigate the optimal ratio between β-6CDP (Fig. 1) and siRNA for light-directed gene silencing at the mRNA level. For this reason we chose to measure gene silencing efficacy at 24 hours after illumination (default incubation time) using different concentrations of β-6CDP and siRNA, either with or without PS (Fig. 2). An illumination dose of 280 J/cm2 was set as default based upon unpublished screening results at the protein level. Our results showed around 90% gene silencing using siRNA concentrations of 0.7 and 1.4 μg in combination with 25 or 50 μg β-6CDP in the samples containing PS, compared with the untreated control. Minor gene silencing effects of around 50% and 70% were measured when using 0.35 μg siRNA in combination with either 12.5 or 25 μg β-6CDP in the samples containing PS, respectively. The results imply that there is no optimal ratio that is independent of siRNA concentration. Importantly, in the samples without PS, siRNA in combination with β-6CDP induced only between 0% and 10% gene silencing at all weight ratios tested. Of note, a 10%–20% decrease in S100A4 mRNA levels was detected in the untreated and scrambled siRNA control samples containing PS compared with the same samples without PS.
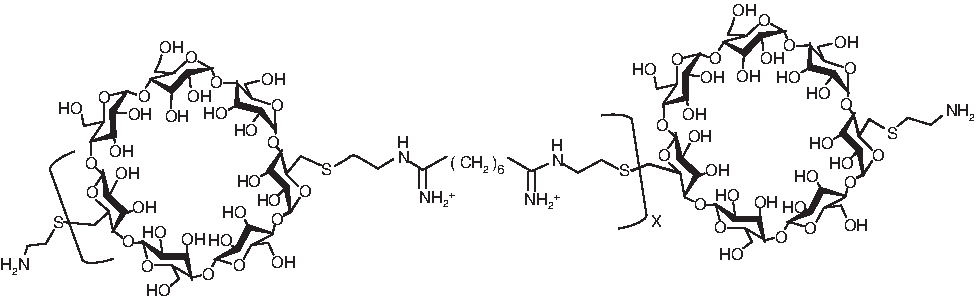
The structure of β-cyclodextrin-containing polymers based on 6 methylene units (β-6CDP).
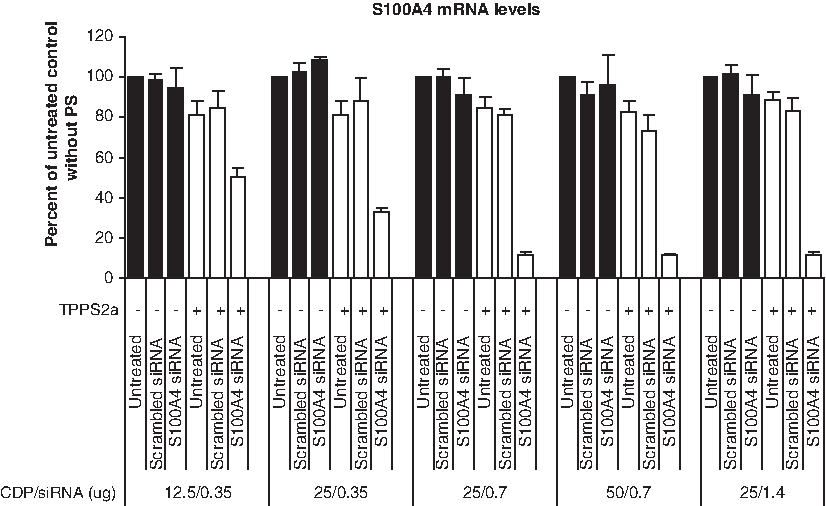
Optimization of CDP and siRNA concentrations upon light-directed gene silencing in the OHS cell line. The gene silencing effect, with or without photosensitizer (PS), was measured as reduction of S100A4 mRNA levels at 24 hours after illumination at 280 J/cm2 using various concentrations of β-6CDP and S100A4 siRNA. Black and white bars display the gene silencing effect without (−) or with (+) PS, respectively. Bars represent the untreated control samples, scrambled siRNA control, and S100A4 siRNA. All samples are represented as percent of the untreated control without PS, and bars represent the mean of 3 individual experiments. Error bars show standard error of the mean ± standard error of the mean (SEM).
Zeta potential of β-6CDP/siRNA complexes
To characterize the β-6CDP/siRNA complexes with respect to light-directed gene silencing, we measured zeta potentials under the weight ratios used in this study. Results showed that the zeta potentials were around +4 mV for all β-6CDP/siRNA complexes complexed in serum-free 1640 medium. Noteworthy, serum-free 1640 medium alone was measured to around −15 mV. In sterile water, zeta potentials were around +50 mV for all β-6CDP/siRNA weight ratios tested.
Optimization of light doses upon light-directed gene silencing
To maximize endosomal escape it is important to optimize the illumination dose for each cell line, well format, carrier, and PS type. We chose in this experiment the complexes consisting of 25 μg β-6CDP and 0.7 μg siRNA, as this was the minimum amount of siRNA and β-6CDP leading to an effective gene silencing in 12-well plates at 24 hours after irradiation. To define the optimal illumination dose resulting in effective gene silencing, we evaluated 3 different light doses, that is, 140, 280, and 420 J/cm2 in the OHS cell line (Fig. 3). Light exposures of 280 and 420 J/cm2 both resulted in similar maximal gene silencing effects of ∼90% reduction, compared with the untreated control. An illumination dose of 140 J/cm2 was less suitable, as it resulted in a moderate gene silencing of ∼65% when compared with the untreated control. The effect of gene silencing in the samples treated without PS was between 0% and 10% for all illumination doses (140, 280, and 420 J/cm2). A decrease in S100A4 mRNA levels dependent upon illumination doses (420 ≥ 280 >> 140 J/cm2) were detected in the untreated and scrambled siRNA control samples containing PS compared with the corresponding samples without PS.
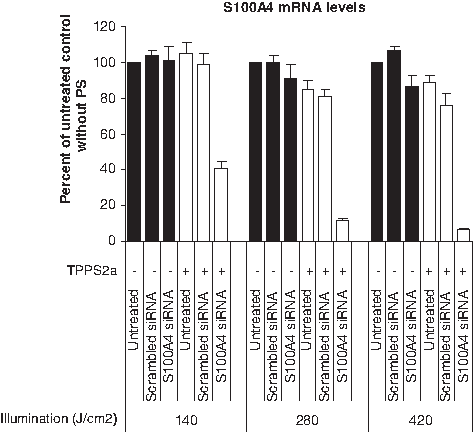
Optimization of illumination doses upon light-directed gene silencing in the OHS cell line. The level of S100A4 mRNA gene silencing was measured at 24 hours after light treatment with 3 different illumination doses (140, 280, and 420 J/cm2) using 25 μg β-6CDP in combination with 0.7 μg siRNA. Black and white bars display the gene silencing effect without (−) or with (+) PS, respectively. Bars represent the untreated control samples, scrambled siRNA control, and S100A4 siRNA. All samples are represented as percent of the untreated control without PS, and bars represent the mean of 3 individual experiments. Error bars show standard error of the mean ± SEM.
The influence of treatment conditions upon cell viability
To minimize possible toxic effects of PCI treatment, it is important to optimize the illumination dose for each cell line, well format, carrier, and PS type. In general, there is a balance between delivery efficacy and cell viability, and it is therefore mandatory to establish a treatment window for optimal results. In this experiment, we employed the same weight ratio of β-6CDP and siRNA as in the Optimization of light doses upon light-directed gene silencing section with 3 different illumination doses of 140, 280, and 420 J/cm2, respectively. Results showed that cell viability measured at 24 hours after irradiation in the S100A4 siRNA samples with PS was reduced by <5% compared with the untreated control without PS, when using an illumination dose of 140 and 280 J/cm2, respectively (Fig. 4). In contrast, an ∼20% reduction in cell viability was measured in the S100A4 siRNA samples with an illumination dose of 420 J/cm2, compared with the untreated control without PS. The untreated control, scrambled siRNA control, and S100A4 siRNA control samples (either with or without PS) showed comparable level of cell viability within each illumination dose.
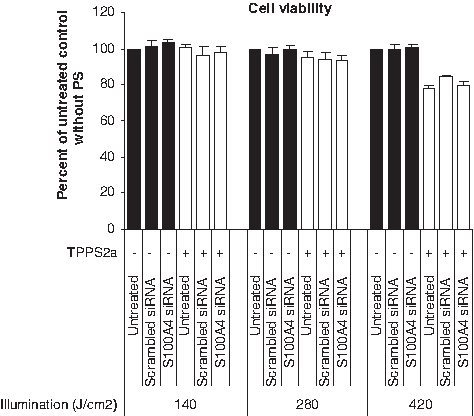
The influence of treatment conditions on cell viability in the OHS cell line. The level of cell viability was measured at 24 hours after light treatment at different illumination doses (140, 280, and 420 J/cm2) using 25 μg β-6CDP combined with 0.7 μg siRNA. Black and white bars display the gene silencing effect without (−) or with (+) PS, respectively. Bars represent the untreated control samples, scrambled siRNA control, and S100A4 siRNA. All samples are represented as percent of the untreated control without PS, and bars represent the mean of 3 individual experiments. Error bars show standard error of the mean ± SEM.
Time-lapse studies of gene silencing efficiency after endosomal and lysosomal escape
To indirectly explore the rate of β-6CDP decondensation from siRNA, we measured the time from endosomal escape to obtain a maximum gene silencing effect (Fig. 5). In this experiment, we employed the same weight ratio of β-6CDP and siRNA as in the Optimization of light doses upon light-directed gene silencing section, with a default illumination dose of 280 J/cm2. Results showed increased silencing in the S100A4 siRNA samples up to 5 hours when the highest silencing effect was reached. All later time points showed the equivalent gene silencing effect in the S100A4 siRNA samples as at the 5 hours time point. As observed for the prior experiments using an illumination dose of 280 J/cm2, a typical 10%–20% decrease in S100A4 mRNA levels was detected in the untreated and scrambled siRNA control samples containing PS, compared with the same samples without PS. Of note, this result was only observed up to the 25 hours time point, as no reduction in S100A4 mRNA levels was detected in comparable samples at 48 hours.
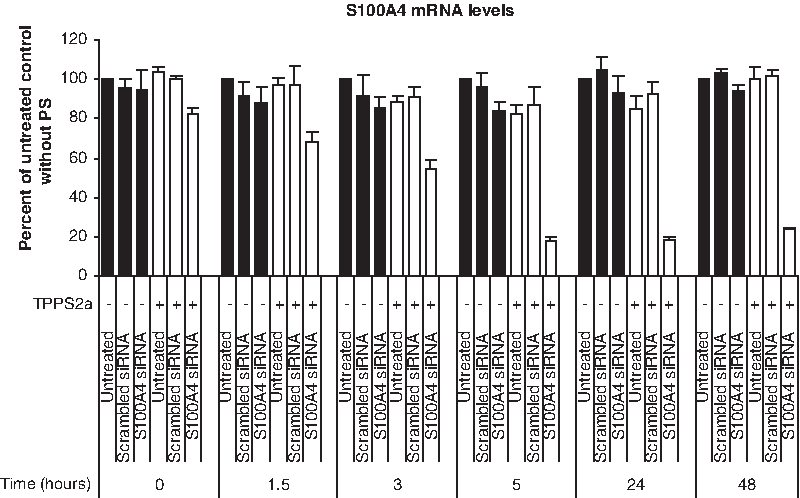
Time-lapse studies of gene silencing efficiency after endosomal and lysosomal escape in the OHS cell line. The rate of gene silencing after endosomal and lysosomal escape was measured at different time intervals after light treatment using 25 μg β-6CDP combined with 0.7 μg siRNA at an illumination dose of 280 J/cm2. Black and white bars display the gene silencing effect without (−) or with (+) PS, respectively. Bars represent the untreated control samples, scrambled siRNA control, and S100A4 siRNA. All samples are represented as percent of the untreated control without PS, and bars represent the mean of 3 individual experiments. Error bars show standard error of the mean ± SEM.
Light-directed siRNA delivery with β-6CDP in the RMS cell line
To show the potential for light-directed gene silencing also in other human cancer cell lines, we chose to test the melanoma cell line RMS. In this experiment, we employed the same weight ratio of β-6CDP and siRNA as in the Optimization of light doses upon light-directed gene silencing section, but with a lower illumination dose (210 J/cm2) selected after optimization. Similar to the gene silencing efficacy in the OHS cell line, 80%–90% silencing was detected in the S100A4 siRNA samples for RMS compared with the untreated control samples at 24 hours after irradiation (Fig. 6). In contrast, only between 0% and 10% gene silencing was obtained in the S100A4 siRNA samples without PS.
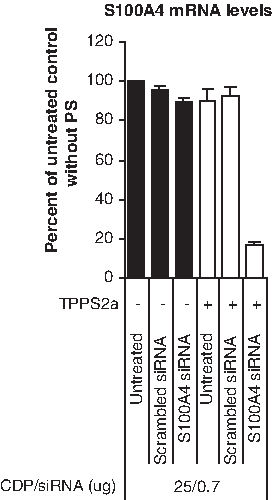
Light-directed siRNA delivery with CDP in RMS. The level of mRNA gene silencing was measured at 24 hours after light treatment in the RMS cell line using 25 μg β-6CDP combined with 0.7 μg siRNA at an illumination dose of 210 J/cm2. Black and white bars display the gene silencing effect without (−) or with (+) PS, respectively. Bars represent the untreated control samples, scrambled siRNA control, and S100A4 siRNA. All samples are represented as percent of the untreated control without PS, and bars represent the mean of 3 individual experiments. Error bars show standard error of the mean ± SEM.
Comparison of different carriers for light-directed siRNA delivery
In this experiment, we compared targeting and silencing efficacy of β-6CDP with carriers previously investigated for light-directed gene silencing. All parameters were kept similar, except for the amount of carrier used (25 μg β-6CDP, 0.3 μg B-PEI, 4.2 μL jetSI, and 2.0 μL LF 2000) and the protocol for complex formation. Experiments were performed using 0.7 μg siRNA with an illumination dose of 280 J/cm2. Results imply an increased targeting specificity at 24 hours after irradiation using β-6CDP compared with the other carriers (β-6CDP > B-PEI > jetSI > LF 2000), as gene silencing without PS increased in the respective order (Fig. 7). The other carriers tested showed higher silencing efficacy with PS under the given conditions (B-PEI > jetSI = LF 2000 > β-6CDP).
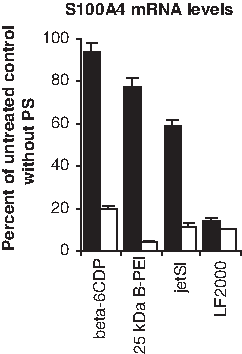
Comparison of different carriers for light-directed siRNA delivery. The level of mRNA gene silencing was measured with or without PS in the OHS cell line using 0.7 μg siRNA combined with 25 μg β-6CDP, 0.3 ng branched polyethylenimine (B-PEI; 25 kDa), 4.2 μL jetSI, or 2.0 μL Lipofectamine (LF) 2000. Illumination dose was set to 210 J/cm2 and cells were harvested at 24 hours after illumination. Black and white bars display the gene silencing effect without (−) or with (+) PS, respectively. All samples are represented as percent of the untreated control without PS, and bars represent the mean of 3 individual experiments. Error bars show standard error of the mean ± SEM.
Discussion
In the field of nonviral gene delivery, there is increasing interest in the development of novel polymer systems that offers enhanced biosafety and biocompatibility. Accumulation of nonmetabolized carriers in animals results in toxicity and severe immune response effects, which are hurdles that should be resolved before nonviral siRNA therapy can become a therapeutic treatment modality in humans. Nonmetabolized carriers based on intrinsic endosomolytic properties (either fusogenic or proton sponge activities) have been investigated for light-directed delivery of siRNA molecules previously (Boe et al., 2007, 2008; Oliveira et al., 2007, 2008), but even these carriers are to a large extent limited by endosomal and lysosomal barriers, especially if they are used under nonoptimal transfection conditions (Boe et al., 2007, 2008). Of note, we have previously shown that the N/P ratio is crucial for endosomal escape of siRNA/PEI complexes either with or without PCI technology (Boe et al., 2008). In contrast to the carriers previously evaluated for light-directed siRNA gene silencing, unmodified biocompatible CDPs do not possess any endosomolytic capacity and are completely protonated at physiological pH (the pKa of the amidine group is ∼12) (Davis et al., 2004). Despite promising results, earlier siRNA/PCI reports using carriers such as jetSI reagent and various PEI formulations containing endosomolytic capacity are less suitable for in vivo applications because of the toxicity of the carrier and the nonspecific targeting effect. In our effort to develop light-directed siRNA gene silencing for in vivo applications, we investigated the potential of the biocompatible and unmodified β-6CDP as a carrier for siRNA delivery. Interestingly, our results showed that maximum gene silencing was reached at 5 hours after PCI treatment. The 5-hour time interval is the total amount of time for the following processes to occur: (1) endosomal release of siRNA/β-6CDP complexes, (2) β-6CDP decondensation from siRNA molecules, (3) cleavage of S100A4 mRNA transcripts by RISC, and (4) S100A4 mRNA degradation in cytosol. These results show that β-6CDP combined with S100A4 siRNA leads to rapid and potent S100A4 gene silencing after endosomal escape. Of note, S100A4 transcripts were also silenced in the untreated and scrambled siRNA control samples containing PS by around 20% when using an illumination dose of 280 J/cm2 (Figs. 2, 3, and 5). This minor silencing effect was time dependent and only observed when using incubation times between 5 and 25 hours (Fig. 5). In contrast, no silencing was detected in S100A4 levels in the untreated and scrambled siRNA control samples when using a longer incubation time (48 hours) or a lower illumination dose of 140 J/cm2 (Fig. 3). These latter results are supported by previous PCI studies, showing both temporal inhibition and activation of transcription when evaluating gene expression shortly after PCI treatment (Prasmickaite et al., 2006; Cekaite et al., 2007). The unmodified β-6CDP used in our study was not effective for siRNA delivery without PS, which implies endosomal entrapment. In contrast, no endosomal entrapment was detected for LF 2000, whereas a high but not complete endosomal entrapment was detected for 25-kDa B-PEI and jetSI reagent. These observations are further supported by earlier studies comparing β-6CDP and β-6CDPim polyplexes condensed with DNA oligonucleotides (Davis et al., 2004; Mishra et al., 2006). Further, our results support the general agreement that LF 2000, 25-kDa B-PEI, and jetSI reagent contains fusogenic or proton sponge activities, which result in endosomal escape. A higher gene silencing efficacy was detected with 25-kDa B-PEI compared with β-6CDP. More favorable particle sizes when using 25-kDa B-PEI compared with β-6CDP is a possible explanation. This is implied by the low amount of 25-kDa B-PEI needed, compared with β-6CDP, which again reflects the ability of the 2 reagents to bind effectively to negatively charged siRNA molecules through cationically charged amine groups. Noteworthy, every third atom of PEI is a protonable amino nitrogen atom. Interestingly, nanoparticles consisting of β-6CDPim combined with polyethylene glycol and transferrin have been successful in systemic delivery of siRNA molecules to tumors in mice without abnormalities in interleukin-12 and interferon-α levels (Hu-Lieskovan et al., 2005). Further, systemic relevant doses of the same nanoparticle were used for the first proof that RNAi works in humans (Davis et al., 2010). In conclusion, recent development shows that nanoparticles consisting of β-6CDP are promising delivery vehicles for in vivo siRNA delivery with the benefit for systemic therapies of cancers. Similar to β-6CDP-based nanoparticles, the PCI technology have shown promise for in vivo applications, that is, PCI-mediated delivery of siRNA molecules has recently been demonstrated by EGFP gene silencing in mice (Oliveira et al., 2008). In this context, an attractive strategy would be to combine β-6CDP-based nanoparticles with PCI technology for systemic tumor targeting.
In conclusion, we have demonstrated a first success in using a biocompatible and unmodified β-6CDP without endosomolytic capacity for light-directed gene silencing with minimal reduction in cell viability. Also, we found that β-6CDP compares favorably with other carriers used with PCI, with respect to higher specificity but not higher silencing efficacy. We foresee that the protocol described here will inspire the development of future in vivo gene therapy applications, using nucleic acids combined with CDs for light-directed delivery.
Footnotes
Acknowledgment
The authors thank Prof. Mark Davis at the California Institute of Technology for providing samples of the CDP materials used here.
Author Disclosure Statement
There is no conflict of interest.