
Abstract
Vascular endothelial growth factor (VEGF) plays a critical role in angiogenesis. The aim of this study was to use chitosan/short hairpin VEGF (shVEGF) [short hairpin RNA (shRNA)-expressing pDNA targeting VEGF-A] complexes in the treatment of rat breast cancer model. Therefore, chitosan/shVEGF complexes were prepared in (2/1) ratio and injected to the breast-tumor bearing Sprague–Dawley rats. Intratumoral and intraperitoneal injections were applied and compared. Tumor volumes were measured during the 36 days. To investigate the effect of complexes on the VEGF expression, VEGF were analyzed by immunohistochemistry and western blotting. The mRNA levels of VEGF were determined by real-time polymerase chain reaction. Tumor volume decreased at the end of experiments after shRNA treatment. The highest suppression in the tumor volume was observed after intratumoral complex injection to rats (96%). Compared with intratumoral and intraperitoneal injection, higher tumor inhibition was obtained with intratumoral injection. Free shRNA injection indicated lower tumor suppression. The immunohistochemistry and western blotting results correlated with the real-time polymerase chain reaction and tumor volume measurements. The data suggest that chitosan/shVEGF complexes can be used to inhibit tumor growth in breast carcinoma model of rats.
Introduction
RNA interference is a recently developed technique to silence proteins in a sequence specific manner by inhibiting mRNA and, consequently, reducing protein expression (BEHLKE, 2006). Many in vivo and in vitro studies demonstrated the efficiency and versatility of short hairpin RNA (shRNA) (Wannenes et al., 2005; Alam et al., 2006; Jere et al., 2008; Lee et al., 2008; Rao et al., 2009). To increase transfection efficiency of shRNA, nonviral delivery systems have been designed such as cationic liposomes, polymers, and lipids (Wannenes et al., 2005; Alam et al., 2006; Niola et al., 2006; Jere et al., 2008). Nonviral polymeric delivery systems have much better safety than viral systems though their transfection efficiency is generally lower cationic polymers (Zhang et al., 2007).
Among the nonviral vectors, chitosan has been extensively used in pharmaceutical applications due to its biodegradability, low toxicity, low immunogenicity, and biocompatibility. Numerous studies on small interfering RNA (siRNA) delivery with chitosan as a carrier have shown effective suppression of reporter genes in vitro and in vivo (Howard et al., 2006; Liu et al., 2007; Jiang et al., 2009; Salva and Akbuga, 2009; Wang et al., 2009). However, no report was available about the chitosan formulation containing shRNA-expressing pDNA for VEGF delivery for inhibiting the experimental breast tumor model.
Although several nonviral delivery systems have demonstrated the therapeutic potential of siRNA in breast cancer cells, only a few in vivo studies have been performed. Chien et al. (2005) used cationic cardiolipin analog-based liposomes for c-raf siRNA delivery in SCID mice bearing human breast xenograft tumors, and 73% tumor growth suppression compared with the group treated with free c-raf siRNA was obtained. A short interfering molecule targeting HER-2 was encapsulated in cationic immunoliposomes modified with antitransferrin receptor antibody fragman and administered to human breast carcinoma tumors xenograft mice. After i.v. treatments, inhibition of HER-2 expression was observed (Hogrefe et al., 2006).
In our earlier study, we developed chitosan/short hairpin VEGF (shVEGF) (shRNA-expressing pDNA targeting VEGF-A) complexes in vitro and obtained (60%) VEGF inhibition in breast cancer cell lines (Salva and Akbuga, 2009). Further, we used the complex formulation applied in the in vivo rat breast tumor model and inhibition of VEGF expression was observed.
In the present study, we evaluated chitosan/VEGF complexes for antiangiogenic therapy in breast cancer model of rats. Therefore, after the intratumoral and intraperitoneal injection of chitosan/shVEGF complexes to the breast tumors bearing rats, tumor growth was measured, and the levels of VEGF expression in tumors were assayed.
Materials and Methods
Materials
psiRNA-mVEGF plasmid vector-based gene silencer system was purchased from InvivoGen. In this study, N-Nitroso-N-methylurea (NMU) (Sigma), chitosan (75 kDa, 75%–85% deacetylation; Fluka), monoclonal VEGF antibody (Santa Cruz), UltraVision Detection System Anti-Polyvalent, horseradish peroxidase (HRP) (Lab Vision), UPL probe, reverse and forward primer, and TagMan Master kit (Roche) were used. All substances of molecular grade were used.
Plasmid constructs for shRNAs
siRNA-expressing pDNA driven by human 7SK and human CMV-HTLV promoter were constructed from psiRNA-mVEGF vector (InvivoGen). The plasmid psiRNA-mVEGF (shRNA) transcribes a single-stranded RNA, which forms stem-loop-structured siRNA, targeted to VEGF mRNA with loop sequences. The target mouse VEGF sequence was siVEGF; 5′-GCUACUGCCGUCCGAUUGAU ucaagag AUCAAUCGGACGGCAG UAGC-3′. The plasmid was transformed into Escherichia coli GT116 bacterial strain. The transformed cells were grown in larger quantities of TB broth supplemented with 100 μg/mL Zeocin. The plasmid was purified using a Geno Pure Plasmid Maxi kit. The purity was checked by agarose gel electrophoresis followed by ethidium bromide staining, and DNA concentration was measured by ultraviolet absorption at 260 nm.
Characterization and preparation of chitosan/shRNA complexes and gel retardation assay
For in vivo studies, chitosan was dissolved in 0.1 M tris acetate/0.1 M acetic acid buffer (pH 5.4) to form a solution of 2.5, and 1 mg/mL of shRNA solution was prepared in the TE (Tris-ethylenediaminetetraacetic acid, pH 8.0) buffer. A series of complexes at various charge ratios was prepared by mixing chitosan solutions with shRNA solution. The mixture was vortexed for 15 seconds and left for 30 minutes for complete forming of the complexes. The volume of shRNA was constant, and the volume of chitosan solution was varied to adjust to theoretical charge ratio (±) (molar ratio of amine to phosphate groups) during complex preparation. The complexes were controlled by 0.7% electrophoresed agarose gel (Salva and Akbuga, 2009).
The diameters and zeta potentials of the nanoplexes were determined using a Malvern Zetasizer (Malvern NanoZess). The instrument is equipped with both a particle sizer and a zetameter unit. The samples were measured in phosphate-buffered saline (PBS) in glass cuvettes at 25°C with a constant angle of 90°. Each measurement was done in triplicate.
Mammary tumor induction
Female Sprague–Dawley rats at 40 days old were supplied from Marmara University Experimental Animals Laboratory. The rats were housed under standard conditions and had free access to water and food. Animal procedures were performed in accordance with approved protocols and followed recommendations for proper care and use of laboratory animals of ethics committee (37.2006.mar).
The NMU was always immediately dissolved before use in 0.9% NaCl adjusted to pH 5 with acetic acid. The animals of the negative control group received 0.9% NaCl solution adjusted to pH 5 with acetic acid only. For tumor formation, rats in the experiment were intraperitoneally injected (i.p.) 50 mg/kg NMU (Sigma) on the days 45–50 of age alternately through the left and right abdominal wall (Thompson and Adlakha, 1991). After the tumor formation, animals were divided into 7 groups (n = 6); chitosan/shVEGF (intratumoral and intraperitoneal), free shRNA (intratumoral and intraperitoneal), chitosan (intratumoral), untreated (positive control group), and negative control groups. The untreated control group received intratumoral and intraperitoneal injections of PBS (pH 7.0) for proper injection formulation. Tumor volumes were measured during the test.
Administration of chitosan/shRNA complexes
Treatment of the tumors was started when the tumor size reached ∼50–60 mm3 (Kim et al., 2008). Chitosan/shRNA complexes (w/w ratio of 2/1, 50 μg chitosan/25 μg shVEGF complexes) were diluted in PBS, and 400 μL was directly injected twice a week by intratumoral and intraperitoneal to rats at a dose of 25 μg shRNA/rat.
The tumors were measured every 4 days, and rats were examined for appearance and growth of necrosis as well as decreased physical activity. Tumor progression was reported as tumor growth over a period of 4 weeks at the end of the experiments. All animals were euthanized, then all palpable tumors of mammary glands and control mammary glands were removed and frozen in liquid nitrogen and stored at −80°C. Tumor size was measured using a caliper across its longest (a) and shortest diameters (b), and its volume was calculated using the formula V = 0.5ab2 (Niola et al., 2006). Following controls such as mRNA quantity, VEGF detection were made.
Immunohistochemical staining
Formalin-fixed, paraffin-embedded breast cancer tissue sections were cut. The sections were deparaffinized in xylene and rehydrated in graded alcohol. Sections were blocked for endogenous peroxidase activity with 3% hydrogen peroxide for 20 minutes. For antigen-retrieval, the slides were microwave-treated in 10 mM citrate buffer (pH 6.0) for VEGF. Nonspecific binding was blocked by incubating the sections for 5 minutes with blocking solution (Lab Vision). After washing with PBS for 5 minutes, monoclonal VEGF antibody for the detection of blood vessels was incubated for 60 minutes at room temperature (Santa Cruz). After incubation with primary antibody, sections were washed thrice in PBS for 5 minutes, followed by two 20 minute incubations with the biotinylated secondary antibody and streptavidin peroxidase complex (UltraVision Detection System Anti-Polyvalent, HRP; Lab Vision) and were subsequently washed in PBS. Staining was visualized with 3.3′-diaminobenzidine tetrahydrochloride. Finally, the slides were counterstained with hematoxylin and mounted for examination (Turhal et al., 2004).
VEGF expression was analyzed by scoring the extent and intensity of staining. Only tumors that had good quality staining of microvessels with the streptavidin biotin complex/HRP technique were evaluated in this study. After the areas of highest neovascularization had been identified, all microvessels were counted on a 1 × 400 field and according to their staining density, cases were scored as VEGF 3(+) when cytoplasmic staining in all tumor tissue was strong and diffuse, 2(+) when staining was moderate, 1(+) when cytoplasmic staining was weak or focal, and negative when no staining was present (Turhal et al., 2004; Bolat et al., 2006). Extent of staining was scored as 0 (0%), 1 (1%–25%), 2 (26%–50%), 3 (51%–75%), and 4 (76%–100%) according to the percentages of the positive staining areas in relation to the whole carcinoma area (Mobius et al., 2007).
RNA extraction and quantitative real-time polymerase chain reaction for VEGF
Total cellular RNA was isolated from tumor tissues using RNeasy Plus Mini kit according to the manufacturer's instructions (Qiagen). RNA was reverse-transcribed using Transcriptor High Fidelity cDNA synthesis kit (Roche Diagnostics) according to the manufacturer's instructions. For the quantitative real-time polymerase chain reaction (RT-PCR), 10 ng of the reverse transcriptase product was used for amplification of VEGF-A or β-actin genes using respective primers and UPL probe, together with TaqMan Universal PCR Mix. Primers used were as follows: VEGF-A, 5′-GGC CTCTGAAACCATCAACT-3′ and 5′-TGCTCCCTTCTTCT GTCGTG-3′; β-actin, 5′-CCCGCAGTACAACCTTCT-3′ and 5′-CGTCATCCATGGCGAACT-3′. The plate was run on the Light-Cycler 1.5 Real Time PCR System (Roche Diagnostics). Standard curves were generated, and the relative quantity of target gene mRNA in each sample was normalized to β-actin mRNA. All reactions were done in triplicate (Chen et al., 2005).
Western blot for VEGF
VEGF protein was detected in tumor tissues by western blot analysis. Tumor tissues were homogenized with RIPA lysis buffer [1× TBS, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 0.004% sodium azide] supplemented with protease inhibitors (Santa Cruz), and protein content was assayed with Bradford assay. Cellular debris was removed by centrifugation (12,000 g, 15 minutes, 4°C), and the proteins (20 μg/lane) were separated on 10% SDS-polyacrylamide gel electrophoresis (Zhang et al., 2008). Samples were heated at 90°C for 8–10 minutes in the presence of 6× sample buffer (7 mL 4× Tris-HCl, 3.6 mL glycerol, 1 g SDS, 1.2 mg bromophenol blue, final volume 10 mL, 4 μL mercaptoethanol for 100 μL buffer) and electrophoresed for 3 hours at 100 mA. A 10% separating gel and a 4% stacking gel were used. The gel was electroblotted onto nylon membranes at 50 V for 2 hours. The membranes were blocked with 3% nonfat dry milk in Triton X-100 containing TBS. Western blot analysis was done with goat anti-mouse VEGF-A as the primary antibody (1:200; Santa Cruz). After extensive washing, the membranes were incubated with mouse anti-goat secondary antibody (1:5000; Santa Cruz) conjugated with HRP. The membranes were developed with chemiluminescence kit (Santa Cruz) on an XPosure film (Kodak). Membranes were tested with rabbit anti-β-actin (1:1000; Santa Cruz) to confirm equal loading (Chen et al., 2005).
Analysis of unspecific immunomodulatory effects
To determine unspecific immunomodulatory effects, serum of animals treated with shRNA and chitosan were analyzed for alterations in interferon-gamma (IFN-γ) expression (Raskopf et al., 2008). One month after treatment initiation, animals were sacrificed and serum was collected. IFN-γ protein that accumulated in the serum and rat IFN-γ protein standard were analyzed using solid phase sandwich ELISA according to manufacturer's instructions (Endogen). Briefly, samples and standards diluted in the assay diluent provided in the kit were incubated at room temperature in wells supplied precoated with specific capturing antibodies. After washing, the detection antibody conjugate was added and incubated at room temperature before washing again. Addition of substrate solution then resulted in color development and, after addition of a stop solution, color intensity was measured in an ELISA reader at 450 nm (Ma et al., 2005).
Statistical analysis
For all graphs presented in this study, each data point represents the mean of at least 3 replicates. Either the standard deviation of the mean or the standard error of the mean was calculated and used to make the error bars. Student's t-tests were applied for comparisons between 2 means. For >2 means, a 1-way or 2-way analysis of variance with a 95% level of confidence (analysis of variance, α = 0.05) was used. Unless otherwise stated, a P value <0.05 was considered to indicate statistical significance.
Results
Preparation and control of complexes
Binding of a cationic polymer to plasmid DNA results in neutralization of negative charges in the phosphate backbone of DNA. This interaction was evaluated by existence of the bands of uncomplexed plasmid DNA (BORCHARD, 2001). As shown in Fig. 1, the release of shRNA from chitosan complexes (partial complexation) was observed at chitosan/shRNA complexes in 0.1/1 ratio (lane 3). Full complexation was seen in 2/1 ratio in electrophoresis (lane 4) (Fig. 1).
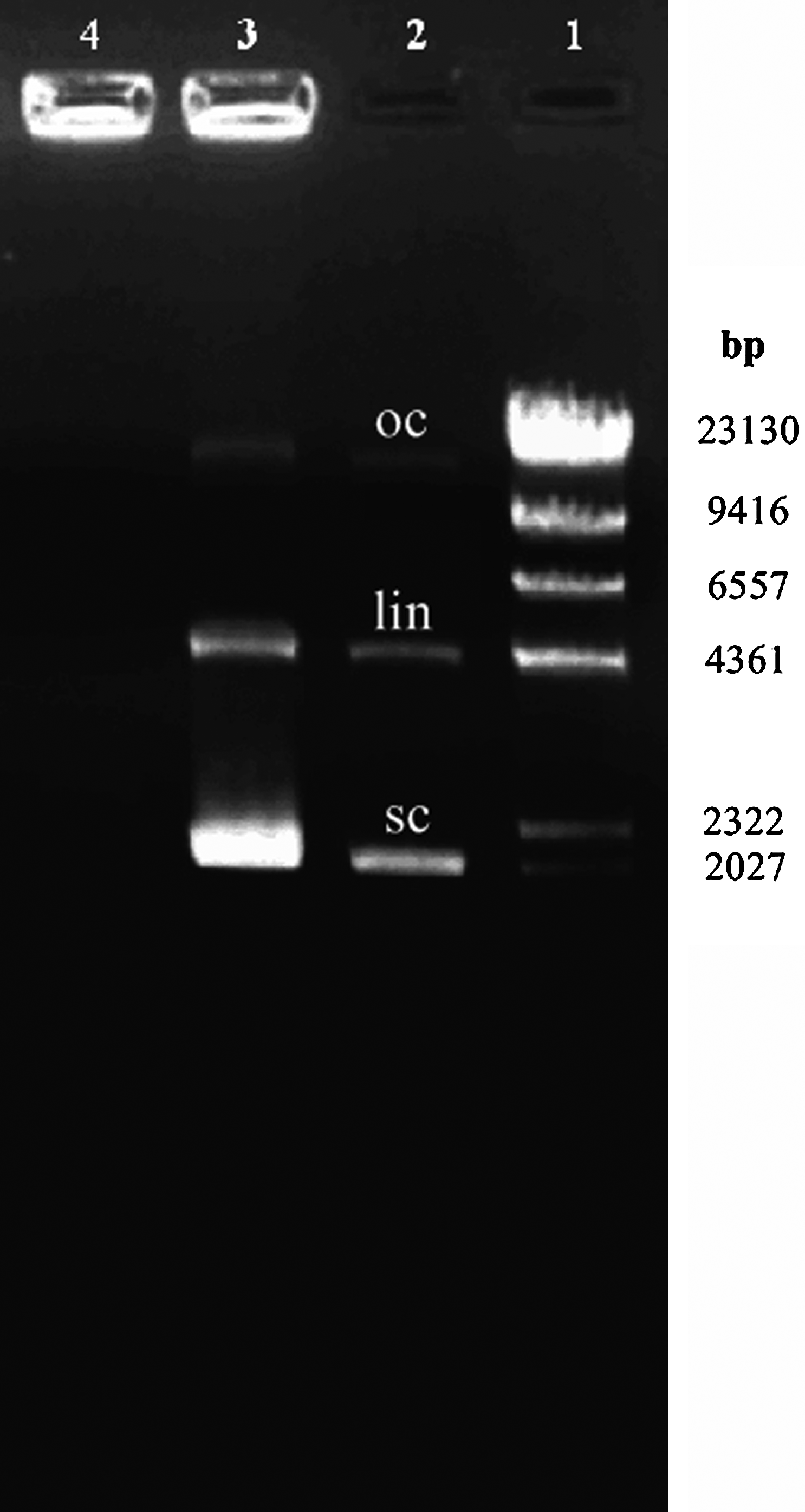
Complexation of chitosan with shRNA-VEGF, visualized by agarose gel electrophoresis. Lane 1, standard DNA (λDNA/HindIII); lane 2, shRNA-VEGF (oc, open circular; lin, linear; sc, supercoiled); lane 3, chitosan/shRNA-VEGF (±) complex (0.1/1); partial complexation; lane 4, chitosan/shRNA-VEGF (±) complex (2/1); full complexation. shRNA, short hairpin RNA; VEGF, vascular endothelial growth factor.
According to our earlier data, chitosan/shRNA complexes in 2/1 ratio were used in in vivo studies. The particle size of chitosan/shRNA complexes (2/1) was 312 ± 15.2 nm, whereas the zeta potential of these complexes was 20.2 ± 4.6 mV.
Inhibition of tumor growth after chitosan/shRNA complex application
To determine the effect of chitosan/shRNA complexes on breast tumor growth, complexes were intratumorally (i.t.) and intraperitoneally (i.p.) injected. Primary tumor growth curves are shown in Fig. 2. After injection, within the first 10 days, tumor proliferation was low except in the control group (untreated tumors). At the 8th day, tumor volume increased in free shRNA applied rats (Fig. 2). Tumor volume reached 500 mm3 after 10 days. This value was ∼5-fold higher than in complex applied groups. About 41.2% of tumor suppression was observed in free shRNA treated animals at the end of the experiment (Fig. 2). During the test, decrease in tumor volumes was determined in animals treated with chitosan/shRNA complexes (P < 0.05). Intratumoral and intraperitoneal injection of chitosan/shRNA complexes reduced tumor volumes by 96% and 89% after 36 days, of application respectively, when compared with controls. Rats having untreated tumors were used as control. In general, the tumor growth was inhibited in the groups treated with chitosan/shRNA complexes. When compared with i.t. and i.p. injection, intratumoral application of chitosan/shRNA complexes inhibited tumor growth more efficiently than did i.p. injection of chitosan/shRNA (Fig. 2). When compared with the untreated group, maximum tumor volume reduction was obtained 96% after i.t. treatment of rats with chitosan/shRNA complexes (P < 0.05). Reduction in tumor volume for all samples was statistically significant when compared with the untreated tumor group (P < 0.05). Free chitosan also indicated suppression effect on tumor growth as seen in Fig. 2. The data concerning tumor volume decrease were supported by RT-PCR and western blot results (Figs. 3, 4 and 5).
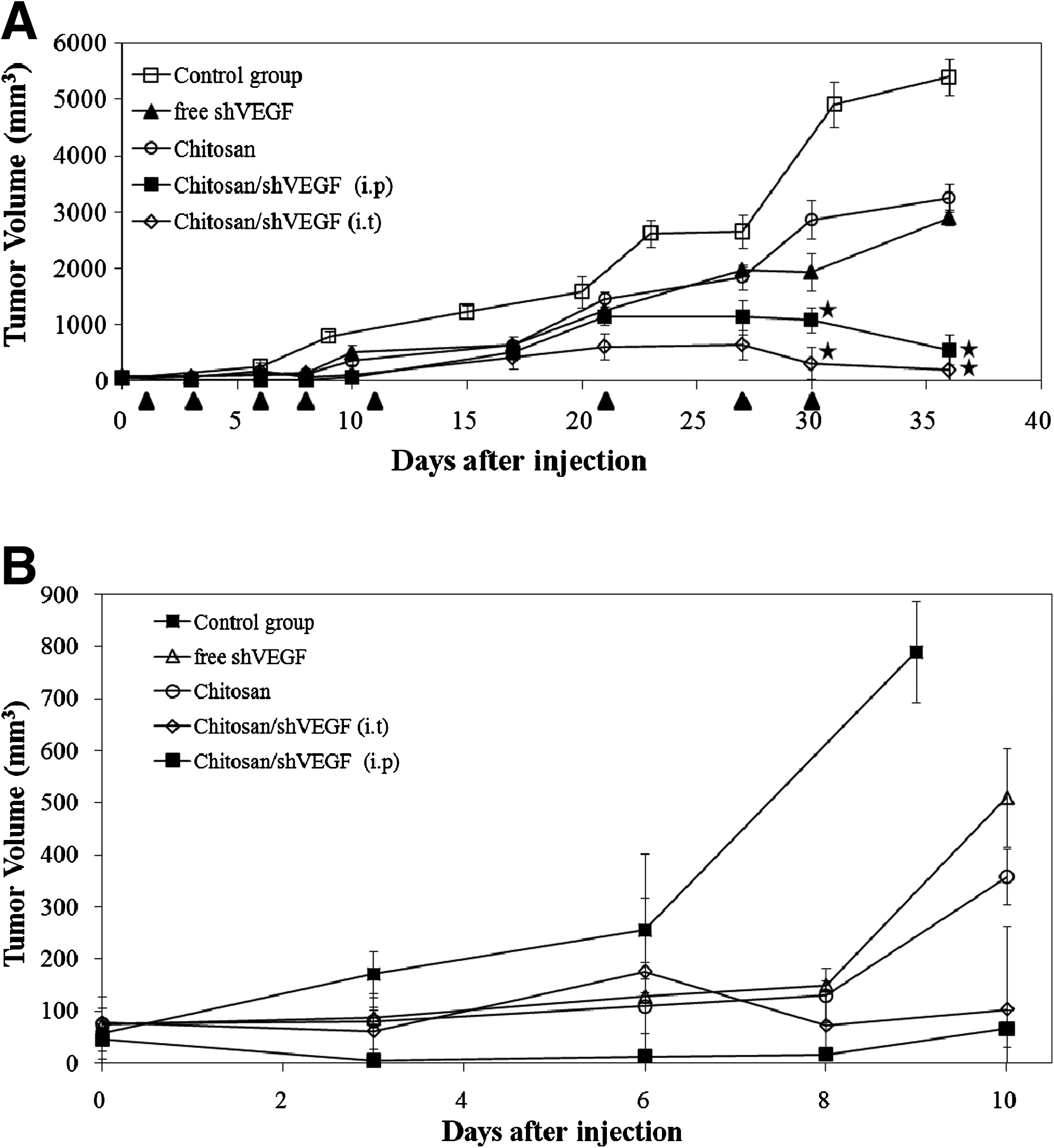
Effects of intratumoral and intraperitoneal injection of a shVEGF on tumor growth in rats. Chitosan/shVEGF complexes were injected into rats i.t. and i.p. as indicated by arrows. On day 1, the tumor volume was ∼50–60 mm3. Tumor diameters were measured at a regular interval for up to 36 days with calipers. Tumor volume measurement. (
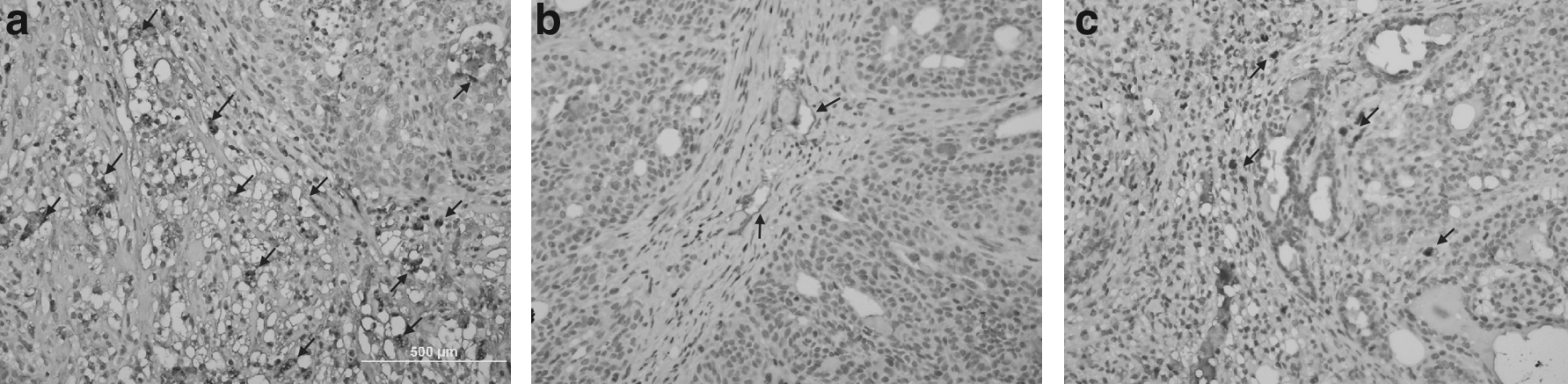
Immunohistochemical analysis of VEGF immunoexpression. Tumor VEGF expression was analyzed by staining with VEGF mAb (magnification 400×). (
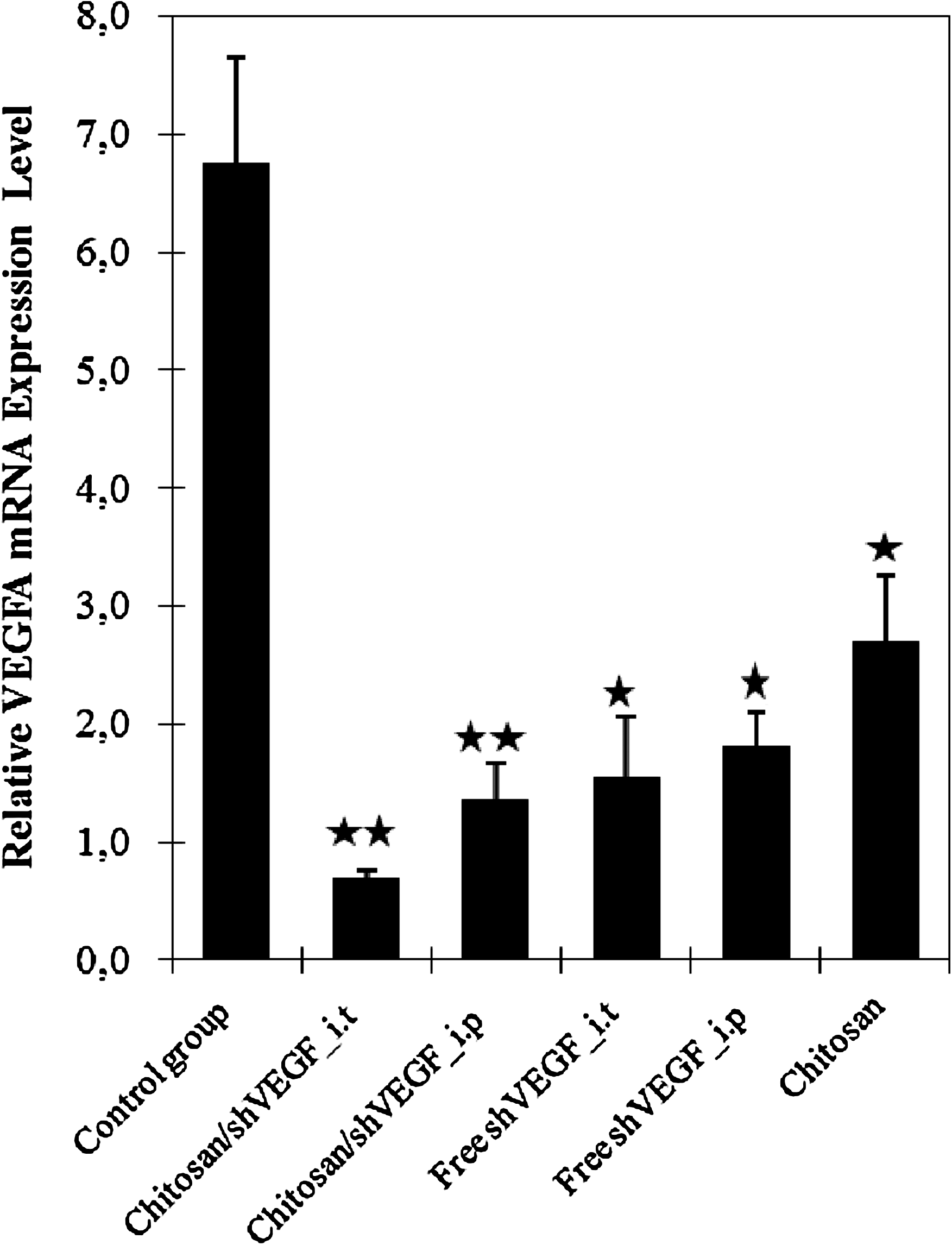
Effect of vector-based siRNA on VEGF expression in rats with breast tumor. VEGF mRNA levels were quantified by real-time polymerase chain reaction. Data were normalized with the housekeeping gene β-actin. Data are represented as mean ± SD of triplicate wells. Similar results were obtained in three independent experiments. *p < 0.05; **p < 0.001.

Western blotting for VEGF. Inhibition rate of shVEGF. Lane 1, control group; lane 2, chitosan/shVEGF complex i.t.; lane 3, chitosan/shVEGF complex i.p.; lane 4, control group; lane 5, chitosan; lane 6, free shVEGF.
Immunohistochemical analysis
Using light microscopy on tumor tissues, gene inhibition effect of chitosan/shRNA complexes was investigated with streptavidin biotin complex/HRP method. Area of invasive tumor containing great numbers of capillaries and small venules (areas of most intense neovascularization), which most frequently occurred at the margins of the carcinoma, were examined by a microscope and were found by scanning the tumor sections at low power.
Few positively stained endothelial cells were detectable in shVEGF injected tumors, whereas positively stained endothelial cells were readily detectable in positive control tumors (Fig. 3a–c). As observed in photographs, when microvessel density and VEGF immunoexpression of shVEGF injected tumor groups were compared with untreated group, there is statistically significant difference between them (P < 0.05). VEGF immunoexpression was reduced ∼67.4% by i.t. applied chitosan/shVEGF complexes (Fig. 3b) and 58.1% by i.p. applied complexes (Fig. 3c). The highest inhibition was found in chitosan/shVEGF complexes injected tumor (Fig. 3b).
Effects of chitosan/shVEGF complexes on the VEGF expression
Further investigation was performed by detecting VEGF using RT-PCR technique from extracted total RNA of the tumor tissues, treated and untreated groups. The experiment was normalized with detection of the housekeeping gene β-actin. Expression of VEGF was analyzed by quantitative RT-PCR. As shown in Fig. 4, the relative VEGF mRNA level in tumor tissue after injection of chitosan/shVEGF complexes and free shVEGF was markedly reduced in comparison with the level of the control group. This result indicates that the tumor-suppressive effect was particularly due to the shRNA for VEGF. VEGF-A expression was reduced ∼84.60% by i.t applied chitosan/shVEGF complexes and 79.60% by application of complexes i.p. Maximum VEGF mRNA inhibition was observed with intratumoral chitosan/shRNA complexes injection.
In addition, western blot analyses showed a significant decrease in VEGF-A protein levels in the chitosan/shVEGF complexes group, compared with the control group (Fig. 5).
Analysis of immunostimulatory effect of IFN
A previous study of Ma et al. (2005) showed that i.v. administration of siRNA was associated with minimal cytokine response in mice. When we compared the presence of serum IFN among all the groups, IFN-γ level in i.p. chitosan/shVEGF complex injected group is higher than in the other group (100 ± 16.3 pg/mL) (Fig. 6).
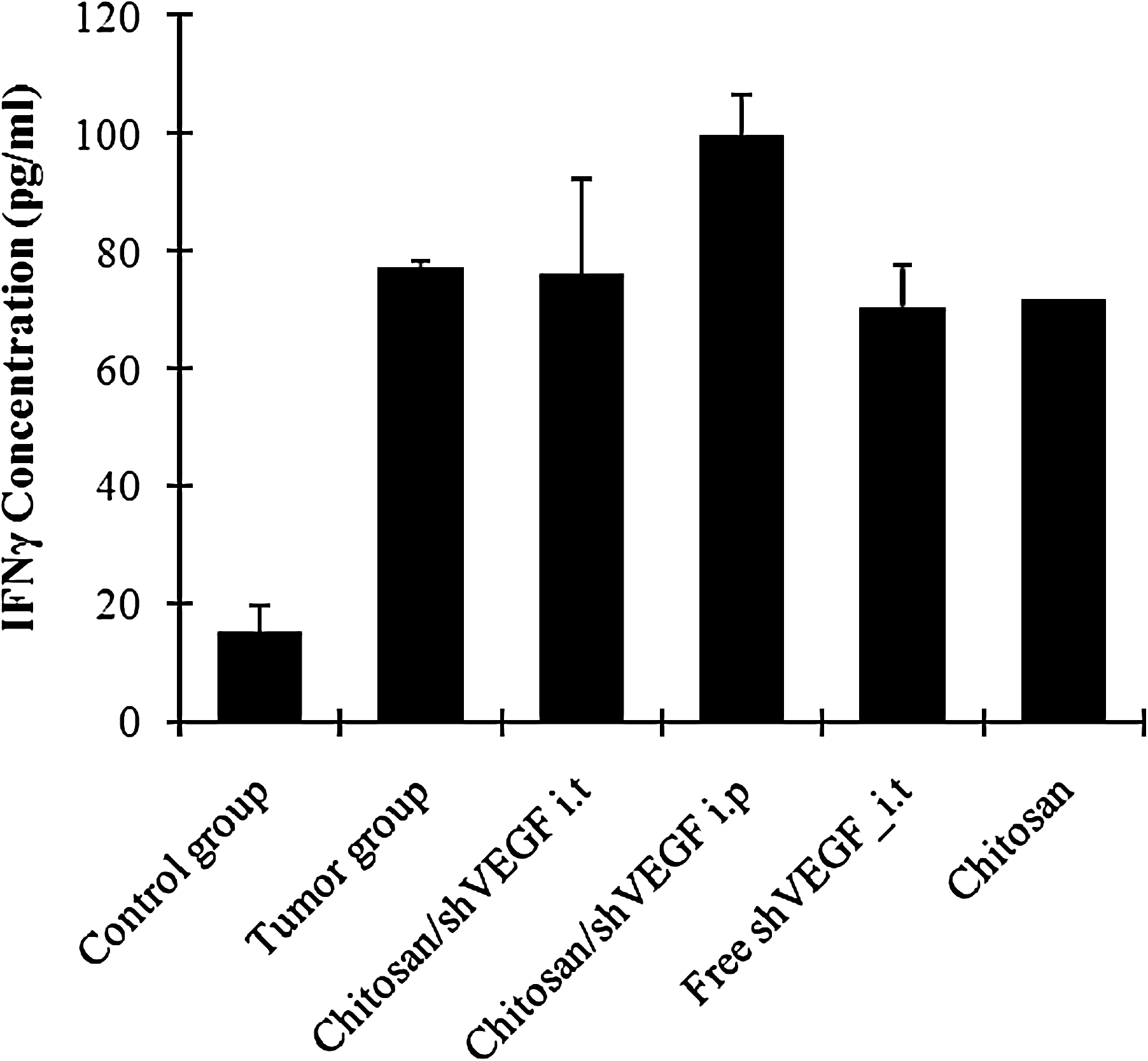
Induction of interferon-γ by chitosan/shVEGF complexes i.t. and i.p., free shVEGF in rats. Data present a mean ± SD (n = 3 per group).
Discussion
Angiogenesis is an essential factor for breast cancer progression and regulated by VEGF. The formation of solid tumors is angiogenesis dependent. Several studies have been developed for targeting the VEGF pathway as a part of anticancer therapy. Recently, attempts were made to use gene specific vector-based siRNA approach for gene silencing (Matsumoto et al., 2006; Niola et al., 2006). Niola et al. (2006) developed and tested vector encoded VEGF shRNA to impair VEGF secretion in U87 human glioblastoma cells and injected PEI/shRNA complexes i.t. to tumors of SCID mice. However, injection of complexes, even if acting against vascularization, was not able to reduce tumor growth, so they used mIL4 as a combined drug in therapy (Niola et al., 2006). Matsumoto et al. (2006) suggested that cationized gelatin microspheres containing shVEGF can be used to normalize tumor vasculature and inhibit tumor growth in a NRS-1 squamous cell carcinoma xenograft model.
The present study was designed to assess the efficiency of i.t. and i.p. administered VEGF shRNA with chitosan complexes in the treatment of breast tumor of rats and to determine the duration of its effect. In our earlier studies, we performed stabilized complexes with chitosan and shRNA (Salva and Akbuga, 2009).
These stable complexes inhibited the proliferation of the tumor, VEGF mRNA and protein expression that affects angiogenesis especially in intratumoral application to rats. There has been considerable interest in the use of RNA interference therapeutics to treat VEGF-induced cancer growth. Recent reports that siRNA targeting VEGF inhibits the secretion of VEGF for tumor antiangiogenic therapy in vitro and in vivo support this notion. Various encouraging results were obtained in several tumor models when VEGF was targeted by employing either siRNA or shRNA (Filleur et al., 2003; Matsumoto et al., 2006; Raskopf et al., 2008).
However, the efficacy of siRNA delivery systems in mammalian cells is still undesirable. Therefore, to increase the activity of siRNA, efficient delivery systems need to be developed for effectively condensing siRNA to small particles, thereby enhancing cellular uptake and protection from enzymatic degradation (Kim et al., 2007).
Even though several nonviral delivery systems have demonstrated the therapeutic potential of siRNA in breast cancer cells, only a few in vivo studies have been performed. Chemically modified siRNA of HER-2 containing nanoimmunoliposome were administered to mice bearing breast xenograft tumors, and inhibition of HER-2 expression was observed (Hogrefe et al., 2006). In another study, 73% tumor growth suppression was obtained after c-raf siRNA:liposome treatment of SCID mice (Chien et al., 2005).
However, no study is available on use of chitosan complexes carrying shRNA in breast cancer models. To investigate the effect of chitosan/shRNA complexes on breast tumor, rats were given 25 μg of shRNA of each animal every 3 days for 10 days, and then injections were made at 17, 22, 27, and 30 days. We observed that tumor proliferation was low due to given tumor tissue of chitosan/shVEGF complexes sequentially during first 10 days. However, tumor volumes increased during the 7 days after treatment, and then it decreased again after a certain time was given to shRNA. Therefore, chitosan/shRNA complexes should be multiple injected for obtaining sustained gene silencing. We suggested that multiple i.t. injection of complexes are a better method for providing sustained antitumor effect. In general, tumor suppression was observed in all the applications except untreated control group (Fig. 2).
Another interesting observation of this study was the antitumor ability of chitosan. This tumor growth inhibition effect of chitosan may be due to glycosis reducing effect of chitosan, thereby decreasing glucose uptake and ATP level in the tumor cells (Guminska et al., 1996). In the growth of cancer, cells require glycosis for energy production; and in this situation, excessive levels of lactate are excreted from cells. This causes the decrease in the extracellular pH of tumors. Tumor acidity produces a more aggressive phenotype of cancer cells with a number of altered behaviors (Dass and Choong, 2008). In addition, the reduction of extracellular pH contributes to the proliferation, invasiveness, metastasis, and angiogenesis of tumor cells. VEGF production is also transcriptionally and translationally regulated by acidosis. VEGF promoter activity rises under low pH conditions (Su et al., 2009).
Before chitosan/shRNA complexes were given to animals, biopsy was taken and tumor identification was pathologically made. According to our immunohistochemical data as seen in Fig. 3, when the VEGF immunoexpression was compared in all the groups, VEGF expression and microvessel density markedly increased in untreated breast tumor. However, VEGF expression significantly decreased in all the tumor treated animals (P < 0.05). The decrease in the microvessel density caused lowering in vascular surface area and permeability. As a result, a higher decline in VEGF expression was obtained in i.t. applied chitosan/shVEGF (67.4%).
RT-PCR and western blotting were applied for the determination of VEGF expression. According to our results, we were found that the silencing effect of intratumoral injection of complexes was higher than the intraperitoneal injection (Figs. 4 and 5). These data are not shown in accordance with earlier reports (Filleur et al., 2003; Raskopf et al., 2008). Filleur et al. (2003), noted that intraperitoneal treatment with siRNA-VEGF was more effective (by 66%) than intravenous or intratumoral treatment of fibrosarcoma model. Differences between our and earlier results may be due to differences in carcinoma types and application (such as ex vivo and in vivo gene application). Data on intraperitoneal treatment with free siRNA-VEGF are rare. In this study, free shRNA-VEGF was i.p. applied to the breast tumors of rats, and 41.2% of tumor growth suppression was obtained (Fig. 2).
In this study, very high gene silencing effect was determined with chitosan/shVEGF complexes (i.t.) in breast carcinoma model of rat (96% in tumor volume, 84.60% by RT-PCR, 67.4% by immunohistochemistry) (Figs. 2–5). Our results were found higher than the earlier reported gene silencing values (Hogrefe et al., 2006). Filleur et al. (2003) reported 66% reduction in tumor growth in the fibrosarcoma model. Tumor growth was inhibited by ex vivo transfer or i.p. application of siRNA-VEGF by 83% or 63% in hepatocellular carcinoma model (Raskopf et al., 2008).
One of the challenges facing siRNA therapy is immune stimulation. The immune system is probably activated via the dsRNA sensor, protein kinase R (Oh and Park, 2009). Innate immunity to siRNA is either Toll-like receptor (TLR)-mediated or non-TLR-mediated. The mediators of nucleic acid stimulated immunoactivation are TLRs 7 and 8 (typically activated by ssRNA), TLR 9 (via unmetthylated CpG activation), and TLR 3 (via dsRNA) (Rao et al., 2009; Robbins et al., 2009).
According to some previous study, the minor antitumor effects observed for siRNA might be caused by immune response originating from siRNA therapeutics (Kim et al., 2008; Raskopf et al., 2008; Oh and Park, 2009). It was previously reported that IFN-α exhibited antitumor activity by an antiangiogenesis mechanism (Kim et al., 2008). In particular, naked siRNA has been shown to activate TLR-3 on the surface of vascular endothelial cells and trigger the release of IFN-γ that mediate nonspecific antiangiogenic effects in vivo (Kim et al., 2008; Kleinman et al., 2008; Rao et al., 2009; Robbins, et al., 2009).
Immune stimulation of the chitosan/shVEGF complexes was found high after i.p. injection, but IFN-γ level of the other groups was similar with the tumor group (untreated positive control group) (Fig. 6).
Conclusion
In conclusion, chitosan is very suitable and an effective gene delivery system for VEGF shRNA in breast cancer models, and very high VEGF suppression and tumor growth decrease are obtained by using chitosan/shRNA complexes. Chitosan complexes can be intraperitoneally and intratumorally applied, and decreased tumor growth and VEGF expression can be obtained.
Footnotes
Acknowledgments
This study was supported by Marmara University Scientific Research Projects Association (SAG-DKR-290906-0185), Turkish Association for Cancer Research and Control (Terry Fox Cancer Foundation).
Author Disclosure Statement
No competing financial interests exist.