
Abstract
Abstract
Abiotic stress can lead to changes in development, productivity, and severe stress and may even threaten survival of plants. Several environmental stresses cause drastic changes in the growth, physiology, and metabolism of plants leading to the increased accumulation of secondary metabolites. As medicinal plants are important sources of drugs, steps are taken to understand the effect of stress on the physiology, biochemistry, genomic, proteomic, and metabolic levels. The molecular responses of plants to abiotic stress are often considered as a complex process. They are mainly based on the modulation of transcriptional activity of stress-related genes. Many genes have been induced under stress conditions. The products of stress-inducible genes protecting against these stresses includes the enzymes responsible for the synthesis of various osmoprotectants. Genetic engineering of tolerance to abiotic stresses help in molecular understanding of pathways induced in response to one or more of the abiotic stresses. Systems biology and virtual experiments allow visualizing and understanding how plants work to overcome abiotic stress. This review discusses the omic approach to understand the plant response to abiotic stress with special emphasis on medicinal plant.
Introduction
An Abiotic stress environment can induce a wide number of responses in plants ranging from readjustment of transport and metabolic process leading to growth inhibition. The primary effect of abiotic stress is ion imbalance and hyperosmotic stress. A direct result of these primary effects is the enhanced accumulation of reactive oxygen species (ROS), which is harmful to the plant cells at higher concentrations. Oxidative stress occurs when there is serious imbalance in any cell compartment among the production of ROS and antioxidant defence, leading to significant physiological challenges. The excess ROS cause damage to proteins, lipids, carbohydrates, DNA, and ultimately results in cell death (Fig. 1).
Response of abiotic stress on plants.
The onset of a stress also triggers some (mostly unknown) initial sensors. They activate cytoplasmic Ca2+ and protein signaling pathways leading to stress responsive gene expression and physiological changes. The stress response is far more complex, with distinct, almost nonoverlapping phases of gene expression. In particular, the numbers of newly active genes (genes that were not active in earlier time points) for each time point is remarkably high.
Plant Response to Environmental Stress
Plant response to environmental stress include water stress, osmotic and salinity stress, as well as exposure to extreme temperatures, wounding, light conditions, and atmosphere is of critical importance both in vivo (greenhouse, field) and in vitro (tissue cultures) conditions. The plant response is complex because it reflects over space and time the integration of stress effects and responses at all underlying levels of organization (Blum, 1996). Under field conditions these responses can be synergistically or antagonistically modified by the superimposition of other stresses. Particularly, salinity and drought are two major abiotic factors that limit crop productivity (Misra et al., 1990, 2002).
Salinity stress
Among abiotic stresses, high salinity stress is the most severe environmental stress, which impairs crop production on at least 20% of irrigated land worldwide. The plant response to salinity consists of numerous processes that must function in coordination to alleviate both cellular hyperosmolarity and ion disequilibrium. Recent confirmation that many salt-tolerant determinants are ubiquitous in plants, has led to the use of genetic models, like Arabidopsis thaliana, to further dissect the plant salt stress response (Hussain et al., 2010). Using the Arabidopsis root tip, Dinneny et al. (2008) examined how different cells within a tissue respond to the physiological stresses due to salinity. Different layers of cells, whether at the surface of the root or more internal, responded differently to the environmental stress of too much salt. Furthermore, stressed cells could influence their neighboring cells, and gene expression patterns changed over the duration of the stress (Hines, 2008). Growth inhibition is a common response to salinity, and plant growth is one of the most important agricultural indices of salt stress tolerance as indicated by different studies (Parida and Das, 2005). Agricultural productivity is severely affected by soil salinity, and the damaging effect of salt accumulation in soils has become an important environmental concern (Jaleel et al., 2007b). The response and growth of plants to saline conditions vary at different stages of growth depending upon the genotype. All plants tolerate salinity up to a certain threshold level without any yield reduction. After that level, an increase in salinity level in the soil has a strong inhibitory effect on the growth and harvestable yield of the plant. When compared to normal conditions (Fig 2A–C), in vitro grown three medicinal plants under stress conditions viz. Chlorophytum borivilianum, Bacopa monierri, and Jatropha curcas showed reduced growth under higher stress concentrations (Fig. 2D–F). The reduction in chlorophyll a and chlorophyll b associated with reduced photosynthetic activity and transpiration rate in plants at all salinity levels is considered a defense mechanism against damaging reactive oxygen species by diminishing light absorbing capacity, which reduces the flow of electrons through the photosystems (Amira et al., 2007). The earliest plant response of salt stress is a reduction in the rate of leaf surface expansion, followed by cessation of expansion as the stress intensifies. The deleterious effect of salinity on plant growth is attributed to the decreased osmotic potential of the growing medium, specific ion toxicity, and nutrient ion deficiency. Saline environments can reduce a wide number of responses in plants, including readjustment of transport and metabolic processes, leading to growth inhibition (Sairam and Tyagi, 2004). Na+ is the predominant soluble cation in most saline soils and water, particularly in coastal areas. Most crop plants exhibit considerable hypersensitivity to saline environments because intercellular accumulation of Na+ is toxic to cellular metabolism, and for many salt-sensitive plants excess Na+ in the soil plays a major role in growth inhibition. High sodium levels disturb potassium (K+) nutrition, and when accumulated in cytoplasm it inhibits many enzymes. These effects are also due to a combination of adverse osmotic gradients and the inhibitory effects of salts and ions on cell metabolism, as well as nutrient imbalance and oxidative stress linked to the production of toxic reactive oxygen intermediates (Jaleel and Panneerselvam, 2007). The chlorophyll a:chlorophyll b ratio also varied significantly under salinity stress. Alkaloid content increases under saline soil conditions (Jaleel et al., 2007a; Wang et al., 2008). A decrease in the fresh and dry mass of Catharanthus roseus seedlings when grown in 1/2 Hoagland solution containing 0–250 mmol×L (−1) of NaCl was noted by Wang et al. (2008). Enzyme activities are also affected in plants under stress. There was a significant enhancement of superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and phenylalanine ammonia-lyase (PAL) tryptophan decarboxylase (TDC) activity reported in plants under NaCl stress (Gao et al., 2008b).

Comparative analysis of plants under stress. (
Dehydration causes a number of physiological and biochemical changes in plants, such as a decrease in photochemical activities, reduction of CO2 fixation, accumulation of osmolytes and osmoprotectants, and alteration in carbohydrate metabolism (Tabaeizadeh, 1998). Also, high salinity (e.g., increased concentrations of Na+ and Cl− in the soil solution) causes osmotic/ionic stress (Hasegawa et al., 2000). An important consequence of salinity stress in plants is the excessive generation of ROS such as superoxide anion and the hydroxyl radicals particularly in chloroplast and mitochondria (Jaleel et al., 2007c; Jaleel et al., 2008; Wang et al., 2001). Generation of ROS such as superoxide, HO, and hydroxyl molecules causes rapid cell damage by triggering off a chain reaction. SOD is one of several important antioxidant enzymes with the ability to repair oxidation damage caused by ROS. Thus, SOD is considered a key enzyme for maintaining normal physiological conditions and coping with oxidative stress in the regulation of intracellular levels of ROS (Mittler, 2002).
High salinity exerts negative impact mainly by disrupting the ionic and osmotic equilibrium of the cell. In saline soils, high levels of sodium ions leads to plant growth inhibition and even death; therefore, mechanisms of salinity tolerance involves sequestration of Na+ and Cl− in vacuoles of the cells, blocking of Na+ entry into the cell, Na+ exclusion from the transpiration stream. After understanding the mechanisms of stress tolerance along with the knowledge of the genes involved in the stress-signaling network, an improvement for high salinity stress tolerance in crops plants can be foreseen (Tuteja, 2007).
Drought stress
Drought stress is also one of the most important environmental factors causing reduction in plant growth and development as well as plant productivity and crop yields (Boyer, 1982). The effect of water stress can be manifest in many ways, as varied morphological, physiological, and biochemical changes in plants under different water stress. For instance, changes in leaf morphology (Parker, 1968), effects on shoot and root growth and development, limit photosynthetic activity by decreasing CO2 influx, decrease in carboxylation, and electron transport chain activities of the chloroplasts in the mesophyll cells (Akinci, 1997). It also affects many other metabolic pathways, mineral uptake, membrane structure (Davies, 1995), stomatal structural changes, conductance, and CO2 uptake (Akinci, 1997).
Water deficit in plants causes the closure of stomata that decreases both transpiration and photosynthesis in many plant species (Fatemy et al., 1985). Stomatal closures occur via the distress signal “abscisic acid” (ABA) and leads to a decreased rate of transpiration from the mesophyll chloroplasts to the guard cells of the stomata during water stress conditions (Wright and Hiron, 1969).
Water stress results in stomatal closure, reduction in transpiration rates, decrease in the water potential of plant tissues, decrease in photosynthesis and growth inhibition, accumulation of ABA, proline, mannitol, sorbitol, formation of radical scavenging compounds (ascorbate, glutathione, α-tocopherol, etc.), and synthesis of new proteins and mRNAs. Besides these physiological responses, plants also undergo morphological changes. The adaptation of plants and chloroplasts to high light (sun) and low light (shade exposure) is also induced by many other stress factors including drought (Lichtenthaler et al., 1981). Along with proteins, lipids are the most abundant component of membranes, and they play a role in the resistance of plant cells to environmental stresses (Suss and Yordanov, 1986). Strong water deficit leads to a disturbance of the association between membrane lipids and proteins as well as to a decrease in the enzyme activity and transport capacity of the bilayer (Caldwell and Whitman, 1987). Poulson et al. (2002) established that for Arabidopsis, polyunsaturated trienoic fatty acids may be an important determinant of responses of photosynthesis and stomatal conductance to environmental stresses such as vapor pressure deficit. When Vigna unguiculata plants are subjected to drought conditions, the enzymatic degradation of galacto- and phospholipids increases. The stimulation of lipolytic activities is greater in the drought-sensitive than in drought-tolerant plants (Sahsah et al., 1998). Water-deficit stress also affects reactive oxygen metabolism in Catharanthus roseus. Jaleel et al. (2008) analyzed the changes in the reactive oxygen metabolism of Catharanthus roseus (L.) G. Don. plants in terms of H2O2 content, lipid peroxidation, and the free radical quenching systems (nonenzymatic and enzymatic antioxidants) under water (drought) stress and concluded that the water-deficit areas may be used for the cultivation of medicinal plants like C. roseus, and the economically important alkaloid production can be enhanced in the plant.
Heavy metal stress
Heavy metal toxicity is one of the major abiotic stresses leading to hazardous health effects in animals and plants. The anthropogenic release of heavy metals into the environment continuously increases soil contamination. Research in the past years, however, has confirmed the immense damage by metal pollution to plants, soil, and ultimately to humans. Although many heavy metals in trace amounts are essential for various metabolic processes in organisms, they create physiological stress when present in high concentration leading to generation of free radicals (Hall, 2002). The generation of ROS is a common event in heavy metal treatment. Regardless, ROS are involved in the oxidation of unsaturated fatty acids, which initiate the formation of oxylipins, a highly variable class of lipid-derived compounds in plants. Oxylipins represent new endogenous signals involved in abiotic-induced stress responses (Mithofer et al., 2004). Higher peroxidase activities contribute to the heavy metal tolerance by alleviating the ROS damage (Chiang et al., 2006).
Because of their high reactivity they can directly influence growth, senescence, and energy synthesis processes (Maksymiec, 2007). The effects of heavy metals like cadmium and mercury showed that there was a reduction in germination percentage, germination index, and seedling length (Peng et al., 2010). Root growth and leaf growth inhibition induced by excess Cu2+ is connected in Z. mays (Maksymiec and Krupa, 2007). Based on their chemical and physical properties, three different molecular mechanisms of heavy metal toxicity can be distinguished: (1) production of ROS by autoxidation and Fenton reaction, (2) blocking of essential functional groups in biomolecules, and (3) displacement of essential metal ions from biomolecules. Transition metals cause oxidative injury in plant tissue, but a literature survey did not provide evidence that increased levels of antioxidative systems could alleviate this stress. The reason may be that transition metals initiate hydroxyl radical production, which cannot be controlled by antioxidants. Exposure of plants to nonredox reactive metals also resulted in oxidative stress as indicated by lipid peroxidation, H2O2 accumulation, and an oxidative burst. Cadmium and some other metals caused a transient depletion of GSH and an inhibition of antioxidative enzymes, especially of glutathione reductase. Assessment of antioxidative capacities by metabolic modeling suggested that the diminution of antioxidants was sufficient to cause H2O2 accumulation. The depletion of GSH is apparently a critical step in cadmium sensitivity because plants with improved capacities for GSH synthesis display higher cadmium tolerance. Available data suggest that cadmium may trigger, via the disturbance of the redox control of the cell, a sequence of reactions leading to growth inhibition, stimulation of secondary metabolism, lignifications, and finally cell death (Schutzendubel and Polle, 2002).
Plants have a range of potential mechanism at different levels that might be involved in the detoxification and thus tolerance to heavy metal stress (Hall, 2002). The main detoxifying strategy of plants contaminated by heavy metals is the production of phytochelatins (PCs) (Cobbett, 2000). PCs, a family of metal-induced peptides, are produced in plants upon exposure to excess heavy metals, such as copper (Cu), cadmium (Cd), or zinc (Zn) (Hall, 2002), and can be detected in plant tissues and cell cultures (Keltjens and van Beusichem, 1998). Several studies have reported that PCs can form complexes with lead (Pb), silver (Ag), and mercury (Hg) in vitro (Rauser, 1999).
Genomic Approach
To improve crop productivity, it is necessary to understand the mechanism of plant responses to environmental stress (Fig. 3). All these abiotic stress never works alone and a complex of various interacting environmental factors contributes at varying degrees to the overall stress. The transduction pathways for osmotic and other environmental stress responses are likely to be very complicated and will involve signal molecules such as ABA, cyclic nucleotides, and inositol polyphosphates. Thus, the precise mechanism(s) by which plants respond to drought or high salinity remains unresolved. Consequently, engineering genes that protect and maintain the function and structure of cellular components can enhance tolerance to stress. However, at the molecular level, most of the changes are likely the result of alterations in the expression of genes. Therefore, it is important to identify the relevant genes and characterize their regulation in response to water and/or salinity stress (Xianan and Bair, 2003).
The molecular basis of abiotic stress.
The stress-responsive genes can be classified into two classes: early and delayed response genes. The former are induced quickly and transiently, whereas the latter are activated more slowly and their expression is sustained. The early response genes encode transcription factors that activate downstream delayed response. This induction does not require new protein synthesis, because all signaling components are already in place. In contrast, delayed response genes, which constitute the vast majority of stress-responsive genes, are activated by stress more slowly and their expression is often sustained. The early response genes typically encode transcription factors that activate downstream delayed response genes. Evidences indicate that salt- or water-deficit response genes are under complex regulation. Although specific branches and components exist, the signaling pathways for salt, drought, cold, and ABA interact, and even converge at multiple steps. This was suggested by mutational analysis in Arabidopsis that single-gene mutation was found to affect responses to all or a combination of these signals (Ishitani et al., 1997).
Gene expression in response to drought stress
Hundreds of genes that are induced under drought have been identified. A range of tools, from gene expression patterns to the use of transgenic plants, is being used to study the specific function of these genes and their role in plant acclimation or adaptation to water deficit. However, because plant responses to stress are complex, the functions of many of the genes are still unknown. Many of the traits that explain plant adaptation to drought—such as phenology, root size and depth, hydraulic conductivity, and the storage of reserves, are associated with plant development and structure are constitutive rather than stress induced. But a large part of plant resistance to drought is the ability to get rid of excess radiation, a concomitant stress under natural conditions. The nature of the mechanisms responsible for leaf photoprotection, especially those related to thermal dissipation, and oxidative stress is being actively researched. Stress-inducible genes have been used to improve the stress tolerance of plants by gene transfer.
It is notable that many drought-regulated genes are also responsive to light (KNAT3, KNAT4, SEN1, DIN9, DIN10, and ACP4) and/or circadian rhythm (e.g., CCA1, WNK1, and FSD1), suggesting that drought might affect a plant's light and circadian cycles and/or vice versa (Huang et al., 2008). Some drought-regulated genes were responsive to multiple stresses and stimuli. For example, RD29B, COR47, and ERD14 were responsive to both water deprivation and cold stress; RD20 and RD22 were responsive to both desiccation and salinity; VSP2 was responsive to heat, oxidative stress, desiccation, insect, salinity, and wounding; and RHL41 (At5g59820) was responsive to salinity, wounding, cold, heat, oxidative stress, chitin, and light stimulus. Swindell (2006) identified 67 genes that exhibited responses to all nine stress treatments (cold, osmotic stress, salt, drought, genotoxic stress, ultraviolet light, oxidative stress, wounding, and high temperature).
The new tools that operate at molecular, plant, and ecosystem levels are revolutionizing our understanding of plant response to drought and our ability to monitor it. In recent years, microarray technology has been applied to identify stress-responsive genes in Arabidopsis (Swindell, 2006). Microarrays are employed to analyze soybean transcriptions under water stress conditions in both root and leaf tissues at vegetative stage. Novel drought responsive genes may be thus identified and used for enhancing drought tolerance in soybean or other plants through genetic/metabolic engineering.
Recently many efforts have been focused on the molecular response of plants to water-deficit stress using the model plant Arabidopsis thaliana (Ingram and Bartels, 1996; Shinozaki and Yamaguchi-Shinozaki, 1997). Many genes respond to drought at the transcriptional level, and their products are thought to function in drought tolerance and response (Bohnert et al., 1995; Bray, 1997; Ingram and Bartels, 1996; Shinozaki et al., 1999; Shinozaki and Yamaguchi-Shinozaki, 2000). Some stress-inducible genes have been used to improve the stress tolerance of plants by gene transfer (Umezawa et al., 2006). Although hundreds of genes have been found to be involved in abiotic stress responses and a number of them have been well characterized (Shinozaki et al., 1999; Shinozaki and Yamaguchi-Shinozaki, 2000, 2007; Zhu, 2002), the functions of the majority of the genes remain unknown and there are probably more genes yet to be discovered.
Gene expression in response to salinity stress
There is a wealth of evidence indicating that changes in gene expression occur in plants following an exposure to salt. During the last decade, a number of salt-responsive genes have been isolated and characterized. Within crop species studied, a wide range of tolerance to salinity exists, from very high (Beta vulgaris) to extremely low (Citrus spp.). Further, within a given crop species, salt-tolerant and salt-sensitive varieties exist. Salt-tolerant species have been frequently employed to isolate genes involved in conferring salt tolerance, and thereby gain an understanding of the mechanisms that distinguish them from their salt-sensitive counterparts.
A number of approaches have been undertaken to isolate genes whose expression is influenced by salinity in plants. A majority of these experimental approaches involves screening cDNA libraries constructed from mRNA populations isolated from plants or cells that have been exposed to a salt treatment. One of the most common and successful methods used to isolate salt-responsive cDNA clones from these libraries has been differential screening with probes derived from mRNA isolated from salt-stressed and nonstressed plant tissue. In this case, mRNA that corresponds to genes that are preferentially expressed in the salt-affected plants is isolated and characterized. More recently, cDNAs that encode proteins that enhance salt tolerance have been selected using salt-sensitive mutants of yeast that are rescued following transformation with plant cDNA libraries.
Pulla and coworkers (2009) reported that cDNA clone containing an S-adenosyl-L-methionine synthetase (SAMS) gene, called PgSAM, may help to protect the plants against various abiotic stresses. They isolated the gene PgSAM from a commercial medicinal plant Panax ginseng. PgSAM is predicted to encode a precursor protein of 307 amino acid residues, and its sequence shares high homology with a number of other plant SAMS. PgSAM is expressed at different levels in various organs of ginseng under various abiotic stresses like salt, salicylic acid, abscisic acid, and chilling stresses.
A gene csa encoding for a citrus salt-stress-associated protein (Cit-SAP) was cloned from Citrus sinensis salt-treated cell suspension by Holland and coworkers (1992). The partial amino acid sequence of the protein, as well as that deduced from the nucleotide sequence of csa, revealed a considerable homology to mammalian glutathione peroxidase (GP), and to clone 6P229 from tobacco protoplasts. The increased expression of Cit-SAP in NaCl-treated cultured citrus cells and in citrus plants irrigated with saline water, and its similarity to GP, raise the possibility that one of the effects of salt stress in plants may be the increase of the level of free radicals.
To identify cold, drought, high salinity, and/or ABA-inducible genes in rice (Oryza sativa), a rice cDNA microarray including about 1,700 independent cDNAs derived from cDNA libraries have been reported from drought, cold, and high-salinity-treated rice plants. The stress-inducible expression of the candidate genes selected by microarray analysis using RNA gel-blot analysis and finally identified a total of 73 genes as stress inducible genes in rice. Among them, 36, 62, 57, and 43 genes were induced by cold, drought, high salinity, and ABA, respectively. Comparative analysis of Arabidopsis and rice showed that among the 73 stress-inducible rice genes, 51 already have been reported in Arabidopsis with similar function or gene name. Transcriptome analysis also reveals novel stress-inducible genes, suggesting some differences between Arabidopsis and rice in their response to stress (Rabbani et al., 2003).
Gene expression in response to nutrient stress
Nutrient deficiency may seriously interfere with normal plant growth and development. Nutrient efficiency is therefore thought to be an important goal in plant breeding. This might refer to macronutrients, such as nitrogen and P, and/or microelements, such as iron or zinc. Related to P depletion, for example, Wu et al. (2003) performed microarray studies in shoots and roots of Arabidopsis that covered transcription analysis of 6,172 genes. About 30% of these genes, that is, 1,835 genes, have been transiently up- or downregulated by twofold or more within the first 72 h under P deprivation. These genes cover a wide range of functions, including more than 100 genes each for transcription factors as well as cell signaling proteins. In the roots, 296 genes were specifically repressed and 141 specifically upregulated. In the shoot, the number of differentially regulated genes was significantly higher and accounts for 617 downregulated and 488 upregulated genes. A further 293 genes were found to be up- or downregulated in the shoot as well as in the roots. It is noteworthy that adaptive reprogramming obviously requires a higher number of downregulated genes than upregulated genes. Expression and signal transduction pathways include transcription factors, protein kinases, and enzymes involved in phosphoinositide metabolism (Kaur and Gupta, 2005). For example, a 104-kDa protein, SAP 104, accumulates in rice seedlings in response to several abiotic stress conditions, and immunological homologues of rice SAP 104 have been detected in several monocot and dicot species. It would be predicted that plants under stress would have a powerful protein turnover machinery to degrade stress-damaged and environmentally regulated proteins. A detailed description is given in Table 2.
Gene expression in response to heavy metal stress
Although analysis of gene expression at the mRNA level has enhanced our understanding of the response of plants to heavy metals, many questions regarding the functional translated portions of plant genomes under metal stress remain unanswered. The carcinogenic effect of metals may be induced by targeting a number of cellular regulatory proteins or signaling proteins participating in cell growth, apoptosis, cell cycle regulation, DNA repair, and differentiation. The carcinogenic effects of certain metals have been related to the activation of transcription factors via the recycling of electrons through the antioxidant network sending the signals to redox-sensitive transcription factors NF-kB, AP-1, and p53. These factors control the expression of protective genes that repair damaged DNA, power the immune system, arrest the proliferation of damaged cells, and induce apoptosis (Valko et al., 2005). As has been observed for most toxic stresses, heavy metal exposure can affect gene expression, through modulation of NF-kB, AP-1, and p53 activities. Plants respond to heavy metal stress by induction of several distinct mitogen-activated protein kinase (MAPK) pathways and that excess amounts of copper and cadmium ions induce different cellular signaling mechanisms in roots. Exposure of alfalfa (Medicago sativa) seedlings to excess copper or cadmium ions activated four distinct MAPKs: SIMK, MMK2, MMK3, and SAMK were reported by Jonak et al. (2004).
Xiang and Oliver (1998) reported that Arabidopsis plants treated with cadmium or copper responded by increasing transcription of the genes for glutathione synthesis, γ-glutamylcysteine synthetase, and glutathione synthetase, as well as glutathione reductase. The response was specific for those metals whose toxicity is to be mitigated through phytochelatins, and other toxic and nontoxic metals did not alter mRNA levels. In response to heavy metal stress, plants synthesize small metal-binding peptides known as phytochelatins (Oritz et al., 1992), PCS genes encode phytochelatin synthases and mediate metal detoxification in eukaryotes (Clemens et al., 1999). Phytochelatins mediate tolerance to heavy metals in plants and some fungi by sequestering phytochelatin–metal complexes into vacuoles. Tonoplast-localized Abc2 transporter mediates phytochelatin accumulation in vacuoles and confers cadmium tolerance (Mendoza-Cozatl et al., 2010).
Expression of the serine acetyltransferase (SAT) gene family from Arabidopsis thaliana was investigated in response to treatment with the heavy metal cadmium (Cd). Expression of all SAT genes was increased to some extent by Cd treatment in the root and stem cortex and the leaf lamella and trichomes (Howarth et al., 2003).
Proteomic Approach
Proteomics also offers a new platform for studying complex biological functions involving large numbers and networks of proteins, and can serve as a key tool for revealing the molecular mechanisms that are involved in interactions between toxic metals and plant species (Ahsan et al., 2009). The products of stress-inducible genes are directly related to protect against these stresses. They include the enzymes responsible for the synthesis of various osmoprotectants like late embryogenesis abundant (LEA) proteins, antifreeze proteins, chaperones, and detoxification enzymes. The various functions of these proteins are enumerated below.
Cellular protection
LEA proteins are mainly low molecular weight (10–30-kDa) proteins, which are involved in protecting higher plants from damage caused by environmental stresses, especially drought (dehydration). These findings and the breeding of drought tolerant varieties would be of great value in agriculture. They form the basis of search for antidrought-inducible genes and their characterization (Shao et al., 2005). They are an ABA-inducible group of proteins and originally suggested to be associated with desiccation tolerance during seed maturation, and are also induced by salinity and water deficit. LEA proteins have been placed in different groups on the basis of amino acid sequence homology. LEA proteins are generally classified into six groups (families) according to their amino acid sequence and corresponding mRNA homology, which are basically localized in cytoplasm and nuclear region. The lea genes encode proteins that are hydrophilic and soluble upon boiling. Most of these proteins lack cysteine and tryptophan, are not compartmentalized or transported within the cell, and are likely to be located in the cytosol. These proteins protect the cellular structure and components, for example, membranes and proteins/enzymes from the effects of water loss during a salt stress.
Ion homeostasis
Survival in a saline environment involves membrane transport leading to exclusion and/or compartmentation of Na+ ions. The transport mechanisms regulate ion transport in order to maintain ion homeostasis and mediate osmotic adjustment via the accumulation and compartmentation of ions within the cell (Dietz and Arbinger, 1996). Sodium, therefore, is actively excluded from the cytosol and sequestered in the vacuole, thus allowing water to move into the cell. Na+/H+ antiporters mediate the uptake/exclusion of Na+. The activities of these carriers are coupled to the downhill flux of H+ leading to a generation of H+ electrochemical gradient across the plasmalemma or tonoplast, which is catalysed by H+-ATPase in the plasmalemma (P-ATPase) and tonoplast (V-ATPase), and H-pyrophosphatase in the tonoplast.
General defense
A number of cDNAs isolated as a result of screening libraries expressed under salinity stress were found to be related to previously characterized genes associated with plant defense against pathogens or wounding damage, for example, PRP from rice, endochitinase from tomato (Chen et al., 1994) and b-glucanase from rice. Even osmotin, a polypeptide initially associated with salinity adaptation, has been related to a family of PRP. Pathogen attack as well as signaling molecules such as salicylic acid and ethylene, which are produced in plants as a result of pathogen attack or wounding, induces its expression in salt affected plants. Protease inhibitor and glyoxylase also plays a role in general defense.
Osmotic adjustment
Genes encoding enzymes like P5CS involved in the biosynthesis of proline, glycine–betaine, and the sugar alcohol pinitol/ononitol have been reported from various plants in response to salt stress. Genes and cDNAs encoding choline monooxygenase and betaine aldehyde dehydrogenase, which are involved in conversion of choline to glycine–betaine, have been observed in response to salinity in sugar beet, spinach, and barley (McCue and Hanson, 2004). Ishitani et al. (1996) was noted sixfold increase in the transcript of myo-inositol phosphate synthase, which is the precursor for pinitol under salt stress. The gene encoding mannitol dehydrogenase, which facilitates its entry into the central carbon metabolism, is downregulated by salt stress. This serves to maintain high concentration of mannitol in salt-stressed cells, to allow it to function as an osmo-protectant.
Metabolism
Some plant like M. crystallinum is a facultative CAM plant, which switches to CAM metabolism under salt stress. This switch helps in conserving water, as the stomata remain closed during daytime. There is also induction of genes involved in the synthesis of CAM enzyme proteins. Induced expressions of PEPCase, NADP malate dehydrogenase, and NADP malic enzyme have been reported in M. crystallinum in response to salt stress. Genes encoding glyceraldehydes-3-phosphate dehydrogenase and phosphoglycero mutase in response to salt stress have been reported in M. crystallinem (Foresthofel et al., 1995).
Gene expression
Some of the salt-responsive genes are those that encode proteins involved in the regulation of other salt-responsive genes. Salt response regulatory genes are mostly transacting factors (Urao et al., 1993) and protein kinases (Mizoguchi et al., 1996). In A. thaliana, the expression of receptor-like protein kinase gene is induced in response to salinity and ABA. These receptors transduce an extracellular signal across the membrane to activate cellular signal transduction pathways. Frandsen et al. (1996) reported an ABA, salt and desiccation-induced gene encoding a protein containing a conserved Ca2+ binding site, suggesting that Ca2+-linked signaling occurs in osmotically stressed plants. A gene encoding a phosphatidyl inositol-specific phospholipase-C (PI-PLC) is expressed in response to salinity and desiccation (Hirayama et al., 1995). This gene is also responsive to ABA, suggesting involvement of Ca2+-linked signaling in the mediation of ABA and osmotic stress responses. Mizoguchi et al. (1995) reported a gene in A. thaliana encoding components of signal transduction and identified it to be MAPK. Other similar genes are MAPKK kinase and a ribosomal S6 kinase, which all function in the MAPK cascade. The expression of these genes increases under salt stress.
Metabolomic Approach
Plant metabolites are characterized by an enormous chemical diversity, every plant having its own complex set of metabolites. This variety poses analytical challenges, both for profiling multiple metabolites in parallel and for the quantitative analysis of selected metabolites. In industrial crops, the content of the economically important metabolite is more important than the yield of the plant part containing the metabolite as it determines the cost of its extraction. It was found that the secondary metabolite production in Catharanthus roseus increased under salinity stress conditions (Jaleel, 2009). State-of-the art genomics tools can be combined with metabolic profiling to identify key genes that could be engineered for the production of improved crop plants. Our limited knowledge of stress-associated metabolism remains a major gap in our understanding; therefore, comprehensive profiling of stress-associated metabolites is most relevant to the successful molecular breeding of stress-tolerant crop plants. Unraveling additional stress-associated gene resources, from both crop plants and highly salt- and drought-tolerant model plants, will enable future molecular dissection of salt-tolerance mechanisms in important crop plants (Vinocur and Altman 2005).
Osmolytes
Compatible osmolytes (Table 3), as osmoprotectants, play a role in counteracting the effects of osmotic stress. Although accumulation of compatible solutes contributes to maintenance of cell growth under conditions of increased ion concentration, many organisms have also developed efficient methods to keep the ion concentration in the cytoplasm at low levels. Membranes, their integral and associated components necessary for the uptake and distribution of ions and solutes, are considered determinants in developing stress-resistant crops.
Proline
Proline (Pro) is one of the most common compatible osmolytes in water-stressed plants. Higher plants accumulate free proline in response to external salt and drought stress. There are reports suggesting that proline at higher concentration acts as an osmoticum, a protective agent of enzyme and cellular structure and a storage compound of reducing nitrogen for rapid growth after stress. The role of proline accumulation and its metabolism viz. tolerant to salinity and drought, therefore need to be critically examined (Tani and Sasakawat, 2006). The accumulation of Pro in dehydrated plants is caused both by activation of the biosynthesis of Pro and by inactivation of the degradation of Pro. In plants, L-Pro is synthesized from L-glutamic acid (l-G1u) via pyrroline-S-carboxylate (P5C) by two enzymes: P5C synthetase (P5CS) and P5C reductase (P5CR). L-Pro is metabolized to l-G1u via P5C by two enzymes: pro-line dehydrogenase (oxidase) [ProDH; EC 1.5.99.8 (EC)] and P5C dehydrogenase [P5CDH; EC 1.5.1.12 (EC)]. Such metabolism of Pro is inhibited when Pro accumulates during dehydration and it is activated when rehydration occurs. Under dehydration conditions, when expression of the gene for P5CS is strongly induced, expression of the gene for ProDH is inhibited. By contrast, under rehydration conditions, when the expression of the gene for ProDH is strongly induced, the expression of the gene for P5CS is inhibited. Thus, P5CS, which acts during the biosynthesis of Pro, and ProDH, which acts during the metabolism of Pro, appear to be the rate-limiting factors under water stress. Therefore, it is suggested that levels of Pro are regulated at the level of transcriptional the genes of these two enzymes during dehydration and rehydration. Moreover, it has been demonstrated that Pro acts as an osmoprotectant and that overproduction of Pro results in increased tolerance to osmotic stress of transgenic tobacco plants. Genetically engineered crop plants that overproduce Pro acquire osmotolerance, namely, the ability to tolerate environmental stresses such as drought and high salinity (Yoshiba et al., 1997).
Proline continues to be the most studied molecule in the decade (Das et al. 1990; Misra et al. 1996). Proline is often regarded as the stress marker. The proline and protein content were evaluated in the in vitro cultures of Chlorophytum borivilianum (Fig. 2A and D), Bacopa monierrri (Fig. 2B and E), Jatropha curcas, (Fig. 2C and F) after a period of salinity stress (Debnath, 2008; Debnath and Bisen, 2008). An increasing proline concentration on both drought and salinity stress was observed as the concentration of the mannitol and NaCl increased. There was also an increase in the proline concentration as the duration (5, 15, and 30 days) of stress increased. These results were found univocally in both salinity and drought stress plantlets. The effect of salt-stress on protein content depended on the concentration of NaCl. At lower levels of NaCl, there was an increase in protein content, but higher concentrations caused it to decline (Fig. 4). There was a similar response on mannitol treatment also. From a comparative analysis on the plants it was evident that Jatropha showed the highest tolerance towards both the stresses, whereas Chlorophytum showed the least tolerance (Debnath and Bisen, 2009).
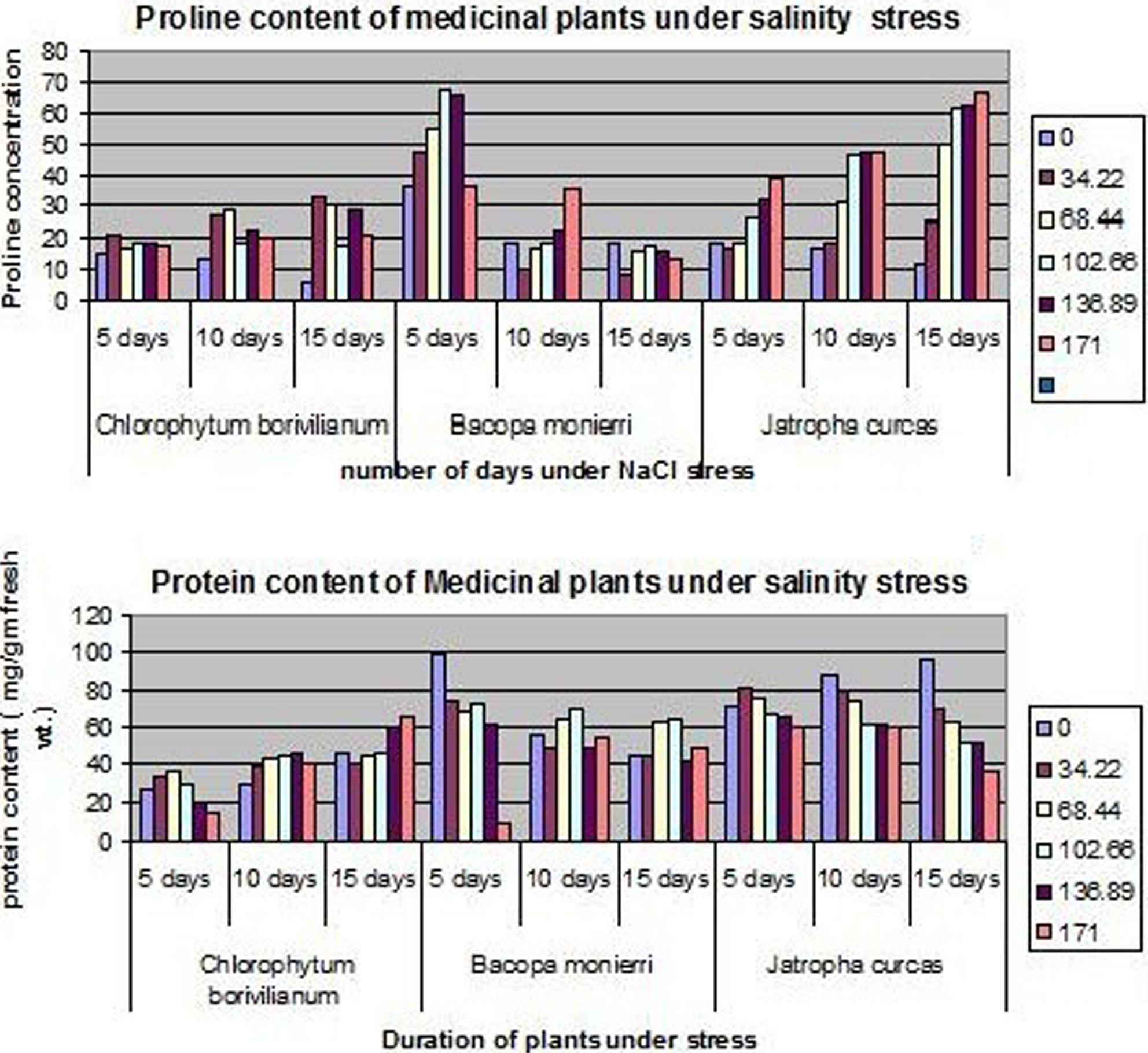
Comparative analysis of proline and protein content of medicinal plants under salinity stress.
In an interesting study by Jaleel et al. (2007a), investigation was conducted to determine whether CaCl2 increases Catharanthus roseus drought tolerance and if such tolerance is correlated with changes in oxidative stress, osmoregulation, and indole alkaloid accumulation. Drought-stressed plants showed increased LPO, H2O2, glycine betaine (GB), and PRO contents and decreased proline oxidase (PROX) activity and increased γ-glutamyl kinase (γ-GK) activity when compared to control, as well as an increase in total indole alkaloid content in shoots and roots when compared to drought stressed and well-watered plants.
On exposure to water deficit or salinity stresses, plants lower the osmotic potential of the cell cytosol and accumulate compatible osmolytes. Proline improves the salt tolerance of plants by protecting the protein turnover machinery against stress-damage and upregulating stress protective proteins (Abdel et al., 2003). Inhibition of growth and a decrease in water content induced by water stress has been universally observed even in tolerant plants. This inhibition may be attributed to the observation that proline-induced cell wall bound peroxidases resulted in increased lignin synthesis, thereby stiffening the cell wall and reducing expansion (Lin and Kao, 2001).
Genes encoding most of the enzymes associated with the synthesis and degradation of proline were cloned and partially characterized, but the factors regulating the expression of these enzymes are largely in the last decade; several attempts were made to increase the level of proline accumulation in plants by transferring the genes associated with the biosynthetic pathway.
Mannitol
With the introduction of the Eshcherichia coli mtlD gene, tobacco plants have been modified. This gene is not produced by wild-type tobacco. Some plants synthesize and accumulate mannitol like transgenic tobacco, transgenic rice with the mtlD gene, and Arabidopsis thaliana with the bacterial mtlD gene (Thomas et al., 1996).
Pinnitol/ononitol
Pinnitol and ononitol are stored in a variety of species that are exposed to saline conditions. In transgenenic tobacco (Vernon et al., 1993), Mesembryanthemum (Vernon and Bohnert, 1992) and many similar plant species or accumulate in tolerant species inositol methyl transferase (Imt) have been obtained whereby the plant is under salinity conditions.
Sorbitol
This sugar alcohol helps in the osmoregulation. It is found in a variety of plant species as an important constituent of seed. Under saline conditions there is an increase in sorbitol concentration (Ahmad et al., 1979) and its accumulation suggest that it may contribute to the dessication tolerance of mature embryo.
Glycine–betaine
Salinity stress induces the activity of betaine–aldehyde dehydrogenase (Weretilnyk and Hanson, 1990). The elevated level of glycine–betaine in Poaceae species is correlated with salt tolerance. Genetic evidence that glycine–betaine improves salinity tolerance has been obtained for barley and maize (Grumet and Hanson, 1986; Rhodes et al., 1989). Transgenic rice plants expressing betaine–aldehyde dehydrogenase converted high levels of exogenously applied betainealdehyde to glycine–betaine than did wild-type plants. The elevated level of glycine–betaine in transgenic plants conferred significant tolerance to salt, cold, and heat stress.
Polyamines
A number of nitrogen-containing compounds accumulate in plants in response to environmental stress conditions (Rabe, 1990). The nitrogen-containing compounds that normally accumulate during stress conditions include amino acids (arginine, proline), amides (glutamine, asparagine), ammonium, quaternary ammonium (glycinebetaine), and polyamines (putrescine, spermidine, spermine).
Polyamines are low molecular weight polycations, and are present in all living organisms. Putrescine is the obligate precursor for spermidine and spermine in all systems studied so far. In higher plants, putrescine is made from ornithine, utilizing the enzyme ornithine decarboxylase (ODC). An alternative pathway in higher plants for the synthesis of putrescine involves arginine decarboxylase. The product of this reaction, agmatine, is then converted to putrescine in two additional enzymatic steps. They also have a possible role in plants under stress conditions. Recent experiments indicate that polyamine levels are not associated with growth inhibition caused by nitrogen deficiency, sucrose starvation, and NaCl stress. Putrescine accumulation seems to be a factor causing growth inhibition under potassium deficiency and phosphate deprivation. Spermidine and spermine seem to be involved in protection against paraquat toxicity in rice leaves (Kao, 1997).
ABA
Abiotic stress causes an increase in ABA biosynthesis, which is then rapidly metabolized following the removal of stress. Many stress-responsive genes are upregulated by ABA. ABA is a regulatory molecule involved in drought stress tolerance. The main function of ABA is to regulate osmotic stress tolerance via cellular dehydration tolerance genes and to regulate plant water balance. ABA is also induced by salt and to a lesser extent by cold stress. ABA-inducible genes have the ABA-responsive element (ABRE) (C/T) ACGTGGC in their promoters. Basic leucine zipper factors (bZIP) function in signal transduction by binding to the ABRE element in stress-inducible genes.
Many accumulated soluble metabolites have been identified by gas chromatography-mass spectrometry (GC/EI-TOF-MS)-based metabolite profiling, indicating a change of metabolic phenotype upon salt acclimation. Ionomic profiling by inductively coupled plasma-atomic emission spectrometry (ICP-AES) revealed salt stress-induced reductions in potassium, phosphorus, sulphur, zinc, and molybdenum. Microarray profiling using the Lotus Genechip® allowed the identification of 912 genes differentially expressed during salt acclimation. Metabolic changes were characterized by a general increase in the steady-state levels of many amino acids, sugars, and polyols with a concurrent decrease in most organic acids. Transcript and metabolite changes exhibit a stress–dose-dependent response within the covered range of NaCl concentrations. The combined observations suggest a successive and increasingly global requirement for the reprogramming of gene expression and metabolic pathways to maintain ionic and osmotic homeostasis during salt stress (Udvardi et al., 2007).
Improving and Understanding Abiotic Stress Tolerance
Plants are typically exposed during their life cycle to conditions of abiotic stress. Most plants have evolved strategies to protect themselves against these conditions. However, if the severity and duration of the stress conditions are too great, the effects on plant development, growth, and yield of most crop plants profound. Continuous exposure to stress causes major alterations in the plant metabolism. These great changes in metabolism ultimately lead to cell death and consequently yield losses. Several lines of evidence have indicated that molecular tailoring of genes has the potential to overcome a number of limitations in creating drought-tolerant transgenic plants (Umezawa et al., 2006).
Strategies for improving stress tolerance
Developing stress-tolerant plants is a strategy that has the potential to solve or mediate of these problems arising due to stress (McKersie and Leshem, 1994). However, traditional plant breeding strategies to develop new lines of plants that exhibit resistance (tolerance) to these types of stresses are relatively slow and require specific resistant lines for crossing with the desired line. Limited germplasm resources for stress tolerance and incompatibility in crosses between distantly related plant species represent significant problems encountered in conventional breeding. Additionally, the cellular processes leading to drought, cold, and salt tolerance are complex in nature and involve multiple mechanisms of cellular adaptation and numerous metabolic pathways (McKersie and Leshem, 1994). This multicomponent nature of stress tolerance has not only made breeding for tolerance largely unsuccessful, but has also limited the ability to genetically engineer stress tolerance plants using biotechnological methods.
The work on genetic engineering of tolerance to abiotic stresses began within a decade of the molecular understanding of pathways induced in response to one or more of the abiotic stresses. In most of the cases the transgenes expressed faithfully but only a limited level of tolerance was provided under stress conditions compared to the nontransformed wild-type plants. In many cases the transgenic plants had morphological abnormalities and slower growth under nonstressed environment. The level of many compatible osmolytes responsible for osmotic adjustment was too low to be effective per se in providing the required water retention and osmotic adjustment. The use of multiple tolerance mechanisms for one or more of the abiotic stresses through stepwise or cotransformation may help to achieve high levels of tolerance for commercial exploitation. The QTL mapping of stress tolerance in certain species, comparative mapping and map-based cloning in plants may be used to screen genes that function under stress as well as those induced and expressed in response to stress.
Molecular understanding of the stress perception, signal transduction, and transcriptional regulation of abiotic stress responsive genes (Table 4) may help to engineer tolerance for multiple stresses. In such studies, transcription factor polynucleotide and polypeptides incorporated into nucleic acid constructs, including expression vectors, have been introduced into plants and were ectopically expressed or by effecting expression of PAP activity. Transgenic plants transformed with many of these constructs have been shown to be more resistant to disease (in some cases, to more than one pathogen), or more tolerant to an abiotic stress (in some cases, to more than one abiotic stress).
One recent advance in transgenic technology of potential value to pharmacognosy is an application of transgenic organ cultures such as hairy roots and shooty teratomas to overproduction and biotransformation of secondary metabolites. The hairy roots induced by Ri plasmid of Agrobacterium rhizogenes have been proved to be an efficient means of producing secondary metabolites that are normally biosynthesized in roots of differentiated plants. So far, the specific metabolites produced by hairy root cultures and/or plants regenerated from hairy roots of 63 species have been analyzed and reported. As an alternative means of producing metabolites normally produced in leaves of plants, the shooty teratomas incited by the tumor-forming Ti plasmid or a shooty mutant of Agrobacterium tumefaciens have been used for the de novo biosynthesis and biotransformation of some specific secondary products. A second and more direct way to manipulate secondary pathways is performed by transferring and expressing specifically modified genes into medicinal plant cells with Agrobacterium vector systems. The genes encoding neomycin phosphotransferase and beta-glucuronidase have been used as model genes under the transcriptional control of appropriate promoters. Recently, some specific genes that can eventually modify the fluxes of secondary metabolism have been integrated and expressed in medicinal plant cells (Saito et al, 1992).
Plant adaptation to environmental stress is dependent upon the activation of cascades of molecular networks involved in stress perception, signal transduction, and the expression of specific stress-related genes and metabolites. Consequently, engineering genes that protect and maintain the function and structure of cellular components can enhance tolerance to stress. Our limited knowledge of stress-associated metabolism remains a major gap in our understanding; therefore, comprehensive profiling of stress-associated metabolites is most relevant to the successful molecular breeding of stress-tolerant crop plants. Unraveling additional stress-associated gene resources, from both crop plants and highly salt- and drought-tolerant model plants, will enable future molecular dissection of salt-tolerance mechanisms in important crop plants.
Signal transduction pathways
The signal transduction pathways in plants under environmental stresses have been divided into three major types: (1) osmotic/oxidative stress signalling that makes use of MAPK modules; (2) Ca2+-dependent signaling that leads to activation of LEA-type genes such as dehydration responsive elements (DRE)/cold responsive sensitive transcription factors (CRT) class of genes, and (3) Ca2+-dependent salt overly sensitive (SOS) signaling that results in ion homeostasis (Kaur and Gupta, 2005). The MAP kinase pathways are intracellular signal modules that mediate signal transduction from the cell surface to the nucleus. These kinases seem to be widely used as osmolarity signaling modules.
MAP kinases are involved in mediating environmental stress responses in plants. Stress adaptation in plants is also known to be mediated by ABA-dependent and ABA-independent processes. Using specific peptide antibodies that were raised against different alfalfa MAP kinases, it was found exclusive activation of p44MMK4 kinase in drought- and cold-treated plants. Although transcript levels of the MMK4 gene accumulated after drought and cold treatment, no changes in p44MMK4 steady-state protein levels were observed, indicating a posttranslational activation mechanism. Although ABA rapidly induced the transcription of an ABA-inducible marker gene, MMK4 transcript levels did not increase and p44MMK4 kinase was not activated. These data indicate that the MMK4 kinase pathway mediates drought and cold signaling independently of ABA (Jonak et al., 1996).
The signaling pathways for abiotic stresses have been reported to converge at multiple steps. This was based on a comprehensive mutational analysis in which Arabidopsis single-gene mutation was found to affect responses to all or a combination of these signals. It appears there are multiple pathways of signal-transduction systems operating at the cellular level for gene regulation. Salts first decrease the osmotic potential of soil solution, creating a water stress in plants. The loss of water from the cells, one of the initial events of water deficit, may affect turgor and bring about changes in size and membrane properties. Although the components of the signal–transduction pathway are difficult to identify, ABA is well known as one such components acting in one of the signal transduction pathways.
Jasmonic acid also acts as an important component in expression of numerous genes and influences specific aspects of plant growth, development, and responses to abiotic and biotic stresses. The Jasmonic acid signal–transduction pathway is mainly unknown. It is presumed that jasmonate interacts with receptors in the cell that activates a signaling pathway resulting in changes in transcription, translation, and other responses (Creelman et al., 1997). Usage of regulatory genes is a more effective approach for producing more salt stress-tolerant plants.
The expression pattern of genes encoding putative transcription factors in ethylene signal transduction was investigated, after treatment of tomato (Lycopersicon esculentum cv. UC-82) seedlings with solutions of differing NaCl concentration for 12, 24, and 48 h. The role of ethylene signaling in tomato seedlings adapting to salt stress is discussed. Le-EIL1 gene expression in tomato seedlings was promoted after 12 h of treatment under different NaCl concentrations. The amount of Le-EIL2 transcript increased after 12 and 24 h in 0.08 mol/L NaCl, and the same effect was observed for Le-EIL3 gene expression in tomato seedlings grown in 0.04 and 0.08 mol/L NaCl. After 12 h, Le-ERF1 gene expression was significantly enhanced by 0.04 and 0.08 mol/L NaCl treatments. Le-ERF2 gene transcriptional expression increased with the application of 0.04, 0.08, and 0.16 mol/L NaCl to seedlings. However, Pti4 gene expression did not change remarkably. These results suggest that different temporal gene expression patterns exist in Le-EIL and Le-ERF family members after NaCl stress induction (Ying et al., 2004).
Calcium-dependent protein kinases (CDPKs) sense the calcium concentration changes in plant cells and play important roles in signaling pathways for disease resistance and various stress responses as indicated by emerging evidences. Among the 20 wheat CDPK genes studied, 10 were found to respond to drought, salinity, and ABA treatments. Consistent with previous observations, one CDPK gene was shown to respond to multiple abiotic stresses in wheat, suggesting that CDPKs could be converging points for multiple signaling pathways. Among the 12 wheat CDPK genes that were responsive to Blumeria graminis tritici (Bgt) infection or the treatment of hydrogen peroxide (H2O2), eight also responded to abiotic stresses, suggesting a crosstalk between biotic and abiotic stress signaling pathways. Phylogenetic analysis indicated that some of these genes were closely related to CDPKs from other species, whose functions have been partially studied, suggesting similar functions in wheat CDPK genes. Combining the up-to-date knowledge of CDPK functions and our observations, a model was developed to project the possible roles of wheat CDPK genes in the signaling of biotic and abiotic stress responses (Li et al., 2008).
Regulation of abiotic stress
Although abiotic stress affects plant growth and development, demonstration of its direct effect on regulation of the components of DNA replication machinery is largely unknown. The subsistence of plants depends on the rapid regulation of gene expression in order to adapt their physiology to abiotic stresses. The expression of nuclear genes is highly regulated at both the transcriptional and posttranscriptional level. Posttranscriptional regulation of gene expression occurs at the levels of premessenger RNA (mRNA) processing (capping, splicing, and polyadenylation), mRNA stability, and mRNA translation.
Differential mRNA translation occurs in response to numerous environmental stimuli such as heat stress (Horiguchi et al., 2000), salt stress (Hua et al., 2001), water deficit (Wood et al., 2000), oxygen deprivation, pathogen infection (Bailey-Serres, 1999), and sucrose starvation (Nicolai et al., 2006). In one these studies, leaves of A. thaliana have been placed under normal or dehydration conditions. The proportion of individual mRNA in polysomes has been measured for over 2,000 genes. The majority of mRNAs show a significant decrease in polysomes association in response to dehydration stress. Although some transcripts are upregulated under stress, their association with polysomes is maintained at the same level as in normal conditions. These results suggest that the effect of dehydration on the translational level varied between mRNA species (Kawaguchi et al., 2004).
Highly efficient methods are available now to study complex alterations at the level of transcripts, proteins, and metabolism, completing our current knowledge. A wealth of data has been recorded recently for changes caused by various abiotic stress inducers and is ready to be analyzed by bioinformatics. All these data show that numerous genes and pre- as well as posttranscriptional and/or -translational factors and processes are involved to achieve appropriate plant responses to environmental circumstances, including mechanisms for stress avoidance as well as tolerance. However crucial to the success of this challenging idea of in silico experimental studies on stress adaptation will the equality and completeness of the data used. Modeling the potential of plants requires a maximum of input of regulatory networks working in plants to cover high adaptability of plants to environmental conditions and the plasticity of the plant reactions. It is importance to understand all levels of regulatory mechanisms. It includes sequence analysis of genes and nongene DNA and studies of transcriptomics and posttranscriptional regulation as well as metabolomics. In addition, profound knowledge on the functioning of the regulatory genes, like transcriptional factors, seems critical to understand the abiotic stress responses (Arnholdt-Schmitt, 2004).
A vital mission of computational stress genomics is identifying genes that respond to diverse environmental stimuli. Regulation, which is determined by chromatin structure, the binding of transcription factors, and cis-regulatory DNA sequences, can be inferred computationally by mining transcript profiles and the regional structure and distribution of short response elements in corresponding promoter regions. Several databases for the analysis of plant promoters exist (e.g., http://www.dna.affrc.go.jp/PLACE and http://intra.psb.ugent.be:8080/PlantCARE). Reconciling stress-regulated genes with promoter structures and then extracting rules about regulation in a cell and tissue context can, in principle, rely on the increasing number of microarray datasets but genome-wide analyses are still few. These analyses have focused on regulatory networks for which response elements, such as dehydration-responsive element binding protein (DREB) or abscisic acid-responsive element (ABRE) in plants, or cAMP-responsive element binding protein (CREB) in yeast, are well known (Proft et al., 2005; Zhang et al., 2005). The relative paucity of results suggests that additional techniques for the identification of promoter-binding proteins like chromatin immunoprecipitation, protein binding to synthetically generated promoters, and in vivo recordings of promoter activity with fluorescent markers (Gibbons et al., 2005; Tran et al., 2004) can be applied. Cell-, tissue-, and treatment-specific transcript profiles from ablated or laser dissected cells or sorted fluorescently painted cells will be essential on the way toward generating a stressed plant in silico (Birnbaum et al., 2005).
Integration of Omic Approach
Some integration of approaches has been achieved by combining transcript analysis and metabolite profiling, reflecting the rationale that downstream results of altered transcription might be most easily documented in this way (Cook et al., 2004; Gong et al., 2005; Hirai et al., 2005; Vinocur and Altman, 2005). The growing attention to metabolite analysis is obvious in the development of metabolite sensors and databases that unite metabolite and transcript profiling (Blank et al., 2005; Deuschle et al., 2005; Fernie et al., 2005; Nikiforova et al., 2005; Thimm et al., 2004). A recognized deficiency at present is the lack of data on metabolite flux parameters (Fernie et al., 2005), which might be alleviated by the introduction of powerful new instruments and protocols for NMR imaging (Krishnan et al., 2005). In contrast, high-throughput protein analyses on the stress proteome are few. Instead, analyses have concentrated on the behaviour of individual proteins or protein families (Feilner et al., 2005; Obrdlik et al., 2004). The requirement for the annotation and integration of data has stimulated work on databases and statistical methods (Thimm et al., 2004). Although not specific to the abiotic stress responses, these tools are essential for the analysis of plant stress data. For example, a superior clustering method, fuzzy k-means (Gasch and Eisen, 2002) assumes that genes could be a part of several pathways, at different weights, and therefore provides flexibility in data analysis that is superior to that provided by classical data clustering. Various profiling platforms can assemble information on abiotic stress-dependent mechanisms that must then be incorporated by bioinformatics into a plant wide stress database to generate new phenotypes, and to allow the information to be exported to crop species in an iterative fashion.
The molecular response of plants to abiotic stresses has been often considered as a complex process mainly based on the modulation of transcriptional activity of stress-related genes. Nevertheless, recent findings have suggested new layers of regulation and complexity. Upstream molecular mechanisms are involved in the plant response to abiotic stress, above all in the regulation of timings and amount of specific stress responses. Posttranscriptional mechanisms based on alternative splicing and RNA processing, as well as RNA silencing, define the actual transcriptome supporting the stress response. Beyond protein phosphorylation, other posttranslational modifications like ubiquitination and sumoylation regulate the activation of preexisting molecules to ensure a prompt response to stress. In addition, crossconnections exist among these mechanisms, clearly demonstrating further and superimposed complexity levels in the response to environmental changes. Even if not widely identified, the targets of these mechanisms characterized so far are mainly regulatory elements of the stress–response pathways. The network of posttranscriptional and posttranslational modifications ensures temporally and spatially appropriate patterns of downstream stress-related gene expression. Future attempts of plant engineering could exploit insights from a deeper comprehension of these emerging sites of regulation of stress responses to develop stress-resistant plants.
Conclusion
The beginning of the 21st century is marked by global scarcity of water resources, environment pollution, increased salinity of soils, and scarcity of waters. Abiotic stress is already a major limiting factor in plant growth. Traditional breeding on environmental stress tolerance succeeded by ingenious selection procedures at the population as well as single-plant level in various environmental backgrounds (years and locations), and by calculating genetic as well as environmental components. These methods automatically considered the complexity of plant organisms (selection at the whole-plant level), as well as the complexity of stress phenomena (testing above years and locations), and have been very successful, although limited by nature. If modern breeding on abiotic tolerance wants to exceed natural limits, a better understanding of the complex interaction between plant and environment is urgently required and has to include research on the adaptive and coordinating capacities of global genome regulation.
Successful transgene strategies to improve abiotic stress have been rare so far, although obviously straightforward. The presence of a transgene sequence in the plant genome cannot be expected to account solely for success or failure in molecular breeding on abiotic stress tolerance. Nevertheless, investigation of all factors involved in global genome regulation of both successful and failing transgenic plants could certainly improve our understanding of complex molecular mechanisms. In this sense, creation of transgenic plants could prove, once again, its worth in leading basic research a significant step forward. Additionally, transgenic RNAi knockout plants will help to understand causal links between global genome regulation and stress adaptation.
The list of genes whose transcription is upregulated in response to stress is rapidly increasing. Functions for some of these polypeptides are close to being identified, and their likely role in stress physiology is being determined. The understanding of mechanisms that regulate gene expression and the ability to transfer genes from other organisms into plants will expand the ways in which plants can be utilized. The exploitation of cloned genes to alter the function of gene products in transgenic plants provides novel opportunities to assess their biological role in a stress response. The molecular analysis of stress responses has arrived at a stage where research can build upon a large collection of characterized genes. Identification of quantitative trait loci for abiotic stress resistance may well be an effective analytical tool. This approach is promising, considering that saturated DNA marker maps are now available for both genetic model plants and crop plants. The use of novel approaches combining genetic, physiological, and molecular techniques should provide excellent results in the near future. Conventional breeding has been of some success only in the case of wheat and rice.
Future progress in systems biology and virtual experiments will no doubt help us to understand how plants work to overcome abiotic stress. However, because models always carry the danger of multiplying underlying minor errors according to the rules of chaos, it will be wise to remember limitations and the potential for misleading. An interdisciplinary glimpse at globally applied computer simulations and their limitations in reliably predicting future perspectives in economy, though based on very well-studied data of the economy markets, may serve as a powerful warning.
Footnotes
Acknowledgments
This work is supported under the scheme of Career award for young teachers AICTE to the first author.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.