
Abstract
Abstract
Clarification of relationships among ploidy series of Cynodon accessions could be beneficial to bermudagrass breeding programs, and would enhance our understanding of the evolutionary biology of this warm season grass species. This study was initiated to elucidate polyploidization among Cynodon accessions with different ploidy series collected from Turkey based on chloroplast and nuclear DNA. Forty Cynodon accessions including 7 diploids, 3 triploids, 10 tetraploids, 11 pentaploids, and 9 hexaploids were analyzed using chloroplast DNA restriction fragment-length polymorphism (cpDNA RFLP), chloroplast DNA simple sequence repeat (cpDNA SSR), and nuclear DNA markers based on neighbor-joining (NJ) and principle component analyses (PCA). All three-marker systems with two statistical algorithms clustered the diploids apart from the other ploidy levels. Assuming autopolyploidy, spontaneous polyploidization followed by rapid diversification among the higher ploidy levels than the diploids is likely in Cynodon's evolution. Few tetraploid and hexaploid accessions were clustered with or closely to the group of diploids, supporting the hypothesis above. Eleven haplotypes as estimated by cpDNA RFLP and SSR markers were detected. This study indicated that the diploids had different organelle genome from the rest of the ploidy series and provided valuable insight into relationships among ploidy series of Cynodon accessions based on cp and nuclear DNAs.
Introduction
Genome doubling (polyploidy) is the presence of more than two chromosome sets in single individual and widespread angiosperms. It is major driving force in plant evolution and plant genomes harbor evidence of multiple rounds of past polyploidization events, often followed by massive silencing and elimination of duplicated genes (Adams and Wendel, 2005; Leitch and Leitch, 2008). Many polyploids experience extensive and rapid genomic alterations, some arising with the onset of polyploidy. This causes increased diversity in higher ploidy levels (Gulsen et al., 2009). Hence, polyploidy may increase adaptation of plant under adverse environmental conditions. Dubcovsky and Dvorak (2007) suggested that genome plasticity coupled with polyploidy was a key factor in wheat's (Triticum spp.) success under domestication. Hence, gene diversification in polyploids may lead to increased polymorphism detected by molecular markers, like SRAP, ISSR, RAPD, simple-sequence repeats, and restriction fragment-length polymorphism. Accordingly, Gulsen et al. (2005, 2009) and Budak et al. (2005) found higher molecular marker diversity in higher ploidy levels among bermudagrasses and buffalograsses [Buchloe dactyloides (Nutt.) Engelm.]. The genus Cynodon is comprised of a morphologically indistinguishable polyploid series of diploids, triploids, tetraploids, pentaploids, and hexaploids with a base chromosome number of x=9 (Wu et al., 2006). Relationships among the ploidy series of Cynodon is largely unknown. It is considered an autoploid (Zeven, 1979). Intra- and interploidy level crosses, except for those involving most triploid and pentaploid parents, can produce viable seeds (Taliaferro, 2003).
Harlan and de Wet (1969) suggested that an area from West Pakistan to Turkey is the center of evolutionary activity for the genus Cynodon, and Gulsen et al. (2009) reported all ploidy series in this region. The Cynodon accessions from far east including China and Korea were missing diploids (Kang et al., 2007; Wu et al., 2006). In Turkish and Chinese Cynodon accessions, tetraploids were the most frequent. Karaca et al. (2002) conducted a study to identify and characterize genetic relationships within and between forage bermudagrass ecotypes and varieties based on various molecular markers including cpDNA-SSR markers. Recently, Jewell et al. (2010) conducted a survey among 1200 Cynodon accessions using microsatellite markers, but provided little information of relationships among their ploidy series. Based on nuclear genome markers, Gulsen et al. (2009) reported that diploids were distinct from the other ploidy levels. Organelle genome-based analysis of ploidy series among the Cynodon accessions was scarce. In addition, Brysting et al. (2011) argued that constructing phylogenetic trees based on nuclear genome markers in polyploids is questionable because of general problems related to incongruent gene and organism lineage phylogenies and methodological challenges connected with retrieving sequence information from polyploids such as polymerase errors and differential amplification of homoeologs (PCR selection).
Organelle DNAs (cpDNA and mtDNA) is a valuable tool for diversity and evolution studies because of the uniparental mode of inheritance of organelle genomes (Curtis and Clegg, 1984; Palmer and Stein, 1986). Of 235 angiosperm species evaluated by Corriveau and Coleman (1988), 192 showed maternal inheritance for cpDNA, the rest of the species showed paternal or biparental inheritance. CpDNAs have been studied at the intra- and interspecies levels in numerous plant species: tomato (Solanum lycopersicum L.) (Sokolonko et al., 1998); Prunus spp. (Mohanty et al., 2000); rice (Oryza sativa L.) (Kumagai et al., 2010); grass family (Poaceae) (Davis and Soreng, 1993); kiwi (Actinidia spp.) (Cipriani et al., 1998); Abies (Parducci and Szmidt, 1999); and Citrus spp. (Gulsen and Roose, 2001); and buffalograss [Buchloe dactyloides (Nutt.) Engelm.] ) (Gulsen et al., 2005). These studies indicate that organelle DNA studies may successfully identify maternal lineages, and interspecies organelle DNA variation is higher than intraspecies variation. These studies used different procedures that vary in cost and efficiency.
Several methods have been used to study variation in cpDNA RFLPs. The first method was common during the 1980s and involves digestion of DNA with restriction enzymes, size separation of fragments on agarose gel, transfer of fragments to membranes, and hybridization with probes to detect specific cpDNA sequences (Lee et al., 1988). The second procedure is the same as the first, with the exception that that no probe-hybridization is required. Instead, after isolation of cpDNA from the nuclear and mitochondrial DNA, and digestion with restriction enzymes, restriction fragments are separated on gel and detected with one of the available DNA detection procedures (Pillay and Hilu, 1990). The third procedure uses “universal” primers anchored within coding sequences can amplify noncoding regions of the cpDNA across species (Taberlet et al., 1991). As a result, PCR-based cpDNA RFLPs emerged, and involves PCR amplification of cpDNA, digestion of PCR products with endonucleases, separation of fragments by electrophoresis, and detection of digested PCR products. CpDNA SSR polymorphism requires PCR followed by fragmentation (Chung and Staub, 2003). The last two procedures can readily use either specifically extracted cp or total DNA extractions via cpDNA gene specific primers that may amplify cpDNA from a great range of plants (Demesure et al., 1995). Recently, because sequencing technology is relatively being cheaper, it is more often used in plant diversity and evolutionary analyses. This study was initiated to elucidate polyploidization among Cynodon accessions with different ploidy series collected from Turkey based on chloroplast and nuclear DNA.
Materials and Methods
Plant materials
The plant materials used in this study were previously described by Gulsen et al. (2009). In brief, plugs of Cynodon accessions collected as representatives from naturally occurring populations in the ruins of gardens of historical sites and undisturbed lands of Turkey were sampled in 2006 and 2007. The 40 accessions were sampled from these, which covers a region bounded by 37°23′27″ to the north, 35°53′51″ to the south, 27°18′42″ to the east, and 36°34′50″ to the west. It ranges from 0 to 1,350 m from the sea level and covers the Mediterranean coastal region of Turkey that lies south of the Taurus Mountains from Izmir to Hatay provinces of Turkey. Geographic barriers in this area (costal Mediterranean part of Turkey) of sampling are ignorable due to free human and animal activities in this region as assessed with analysis of molecular variance (Gulsen et al., 2009). The Cynodon accessions used in this study included 7 diploids, 3 triploids, 10 tetraploids, 11 pentaploids, and 9 hexaploids as determined by Gulsen et al. (2009) (Table 1). The diploids and triploids were fewer due to insufficient numbers of Cynodon accessions available. Diploid species Cynodon transvaalensis Burt-Davy were used as control.
CpDNA RFLP and cpSRR analyses of Cynodon accessions
Single fresh leaf was used for DNA extraction. All genotypes were vegetatively cloned using a single stolon from each population. Total DNA was extracted from 100 mg fresh leaf tissue according to a modified CTAB DNA extraction procedure (Doyle and Doyle, 1990). DNA pellets were diluted with 300 μL of TE (10 mM Tris, 0.1 mM EDTA, pH 7.4), and their concentrations were measured with a microplate reader (PowerWave HT, BIO-TEK Instruments, Inc.,Winooski, VT), and 10 ng μL−1 DNA templates for PCR amplifications were made using double distilled water.
For cpDNA RFLP study, five chloroplast primer pairs anchored at coding chloroplast DNA regions and tested previously for a range of plant species (Demesure et al., 1995) were used to amplify coding and noncoding regions (Table 2). CpDNA regions were amplified in a 50-μL reaction consisting of 30 pM of each of the primer pairs, 0.33 mM of each of dNTPs, 2.5–3 mM of MgCl2 as a final concentration, and 1 unit of Taq polymerase (Fermentas Inc., Glen Burnie, MD), and 10 ng of template. Cycling parameters included the following: one cycle of 2.30 min at 94°C, 35 cycles of 1 min at 94°C, 1min at 60°C, 2 min at 72°C, and for extension, one cycle 10 min at 72°C. Restriction enzymes used in this study are as follows: Mse I, EcoRI, Msp I, Hin1 II, Taq I, Dra I, BsuRI, Mbo I, Rsa I, and EcoRV. Restriction analyses were according to their manufacturer's manual (Fermentas Inc.). They were initially selected on the basis of observation of the rice chloroplast genome sequences as the closest relative among available complete sequences by the restriction analysis option of VECTOR NTI Suite 8 software (Invitrogen Corp., Carlsbad, CA). Restriction fragments were separated on 2% (w/v) agarose gels containing ethidium bromide, and were photographed.
Expected sizes were estimated based on organelle data of buffalograsses by Gulsen et al. (2005).
Expected sizes were estimated based on organelle data of tobacco by Chung and Staub 2003).
For the cpSSR study, 10 cpSSR primer sequences developed from tobacco (Nicotiana tabacum L.) chloroplast sequences (Chung and Staub, 2003) were used to screen cpDNA polymorphism in the Cynodon accessions (Table 2). CpSSRs were amplified in a 15-μL reaction. The reaction mixtures were the same as above. Cycling parameters included the following: one cycle of 5 min at 94°C, 35 cycles of 1 min at 94°C, 1 min at 55–65°C, 1 min at 72°C, and for extension, and one cycle of 6 min at 72°C. PCR products amplified with cpSSR primers were separated on 6% polyacrylamide gel stained by ethidium bromide and visualized and documented. Nuclear DNA profiles of the 40 Cynodon accessions were extracted from Gulsen et al. (2009) and reanalyzed for elucidating ploidy complex of the genus Cynodon.
Data analyses
Each restriction fragment was scored as present (1) or absent (0), and data were analyzed with the Numerical Taxonomy Multivariate Analysis System (NTSYS-pc) version 2.1 software package (Exeter Software, Setauket, NY) (Rohlf, 1993). A dissimilarity matrix was constructed on the basis of average taxonomic distance coefficient and used to construct a dendrogram based on the neighbor-joining (NJ). The NJ method by Saitou and Nei (1987) is a method for estimating phylogenetic trees and based on the idea of parsimony (it does yield relatively short estimated evolutionary trees). The algorithm starts with a matrix of distances among the OTUs (operational taxonomic units). The closest pair of OTUs is found, merged into a new HTU (hypothetical taxonomic unit) and the original pair of OTUs is deleted until the matrix is reduced to a single HTU and the tree is fully resolved. It produces an unrooted graph and estimates the length of each branch (edge) of the graph. These results in a tree that may not have equal branch length or path length may not be ultrametric.
Principal component analysis (PCA) was used to estimate clusters and relationships among the accessions on two-dimensional plot. First, dissimilarity matrix was produced using variance-covariance procedure with SIMINT module and, then used to construct two-dimensional scattered plot with based on EIGEN module. Finally, to produce the actual ordination of the genotypes, the PROJ module is used to project the observations onto the principal component axes.
Association between cpDNA- and nuclear DNA-based data was estimated, which may help understanding diversification of cpDNA RFLP and cpSSR regions. The MXCOMP module was used to calculate correlation coefficients between similarity matrix based on nuclear data previously produced by Gulsen et al. (2009) and average distance matrix based on cpDNA data in this study. This procedure is called the Mantel test, and indicates whether two data produce similar similarity matrices (Mantel, 1967).
Results
Five different regions of cpDNA were amplified with primer pairs targeting rbcL, psb-trns, trnHK, trnST, and 30S regions (Fig. 1). Annealing temperature was 60°C for all primer pairs. Sizes of amplified fragments obtained and expected were listed in Table 2. The amplified fragments were digested with 10 restriction enzymes as listed in Table 3. Cp DNA regions of 40 genotypes of bermudagrass accessions were amplified by using 5 cpDNA specific universal primers (rbcL, psb-trnS, trnHK, trnST, and 30S). Primer pairs usually produced expected size fragments (Table 2) and those fragments were 100% reproducible. To detect RFLPs in those regions, 10 restriction enzymes (Taq I, Rsa I, Hin1 II, BsuRI, EcoRI, EcoRV, Mbo I, Mse I, Msp I and Dra I) were used to digest amplified cpDNA fragments. All digestions of amplified fragments produced scorable bands. Of the 143 bands obtained from cpDNA-RFLP, 53 (% 37) were polymorphic among the 40 Cynodon accessions. The size of the restriction fragments ranged between 100–1900 bp. The polymorphism was the highest (100%) in the restriction analyses of the trnHK–DraI, trnHK–Msp I, rbcL–BsuRI, rbcL–Rsa I, 30S–Hin1 II fragment-restriction enzyme combinations.
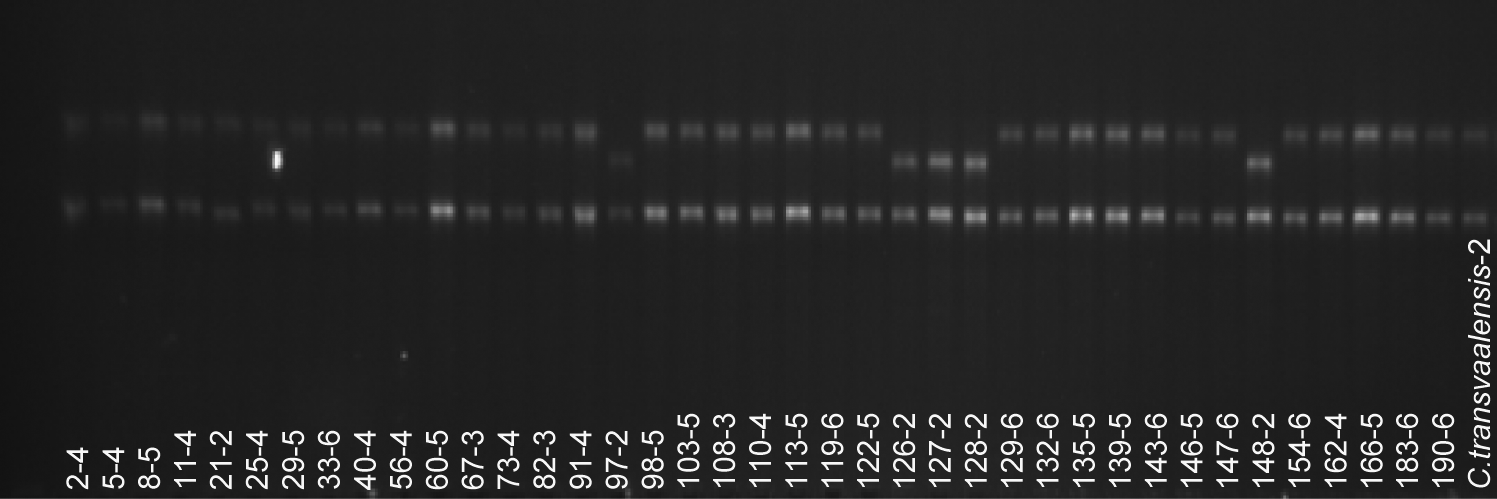
Gel images of 40 Cynodon accessions amplified with trnHK primer and digested with Taq I restriction enzyme. The second digit (after dash: ‘-’) indicates ploidy level of accession. Abbreviation C.transv-2 was used for diploid C. transvaalensis.
No restriction site.
In addition to five loci of cpDNA, 10 pairs of different cpDNA-SSR primers previously developed from tobacco cpDNA sequences were tested to detect cpDNA diversity in Cynodon accessions (Chung and Staub, 2003) (Table 2). Of the 10 primers tested, 8 produced reproducible and specific bands with close to the expected size of amplification product size when compared to size of tobacco SSR length (Table 4, Fig. 3). Annealing temperatures were between 46 and 59°C for 10 primer pairs. The level of the polymorphism was 87%, which was much higher compared to cpDNA RFLPs obtained in this study. The size of the obtained bands ranged between 190 and 240 bp. CpDNA-SSR-5 and 9 produced three different banding pattern among the genotypes studied. CpDNA SSR-4 and cpDNA SSR-15 primers produced single banding pattern among the genotypes studied. Considering all the Cynodon accessions screened in this study, cpDNA-SSR-5, 8, 12, 19, and 22 primers produced 100% polymorphism.
Total length of the amplified region spans 9,230 bp (base pairs). Of the 9,230 bp, 2,230 bp corresponds to cpSSR, and the rest is the cpDNA gene regions used for cpRFLP. This represents about 5% of the chloroplast genome.
Considering all amplified fragments, the number of scored fragments ranged from 2 (EcoRV) to 24 (Mse I)) (Table 3). The numbers of scorable bands were the highest with 32 in trnsT gene region and the lowest in rbcL gene region with 20. Considering the restriction enzymes used for RFLP analysis, the numbers of scorable fragments were the lowest with EcoRV in all chloroplast DNA regions studied. Only the trnHK gene region had one restriction site and produced two fragments for that enzyme. Mse I, a four-base cutter enzyme, had six restriction sites and produced seven fragments in psb–trnS gene region.
The NJ approach was used to estimate relationships among the 40 Cynodon accessions for both separate and joint data sets of cpDNA RFLP and SSR (Fig. 2). CpDNA RFLP-based NJ tree indicated an average distance of 0.0–0.32. The 28 accessions, the largest group, were not distinguished from each other. This group consisted of all ploidy levels from triploid to hexaploid, excluding diploids. The second largest group included five diploids and one triploid. In this group accession 108-3 was clustered with C. transvaalensis. Accessions 91-4 and 147-6 were clustered in an intermediate position between the diploids and the other ploidy levels.
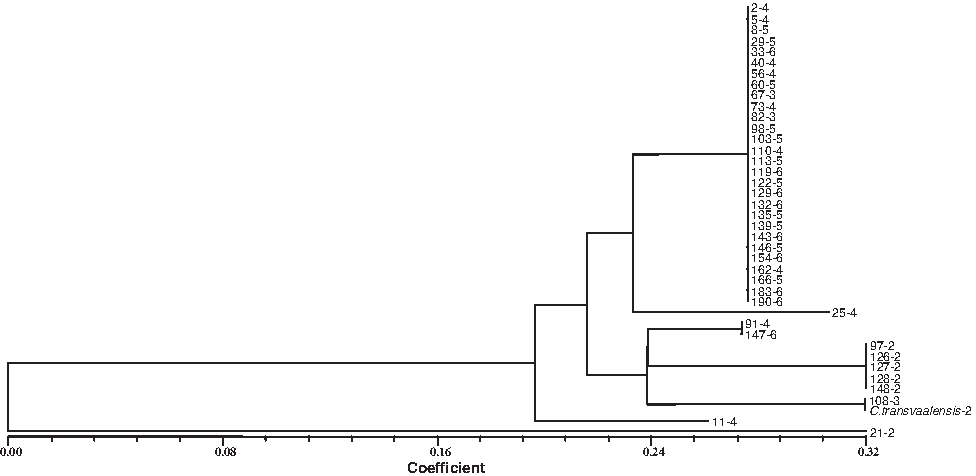
Neighbor-joining tree of the 40 Cynodon accessions constructed by using 145 cpDNA RFLP fragments obtained from restriction analyses five cpDNA regions.
The cpDNA SSR-based NJ analysis indicated slightly different results. Average distance among the accessions ranged from 0.0 to 0.41 (Fig. 3). The largest group included 21-2 accessions, which were not distinguished. Similarly, this group included all ploidy levels except for diploids. The second largest group consisted of four diploids and one hexaploid (147-6), which were not distinguished from each other. As revealed by both cpDNA data, accession 21-2 was uniquely different from the rest of the genotypes. Cynodon transvaalensis, 97-2, and 21-2 were intermediate between the largest group and diploids' group.
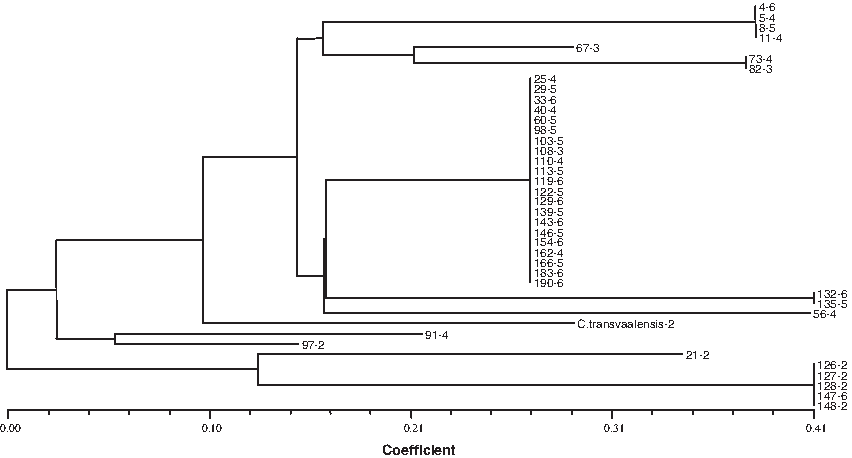
Neighbor-joining tree of the 40 Cynodon accessions constructed by using 16 cpSSR fragments obtained from PCR amplifications 10 cpDNA regions.
From our previous study (Gulsen et al., 2009) of 192 accessions, nuclear data of the 40 accessions were reused to construct NJ-based dendrogram (Fig. 4). Similar results were obtained from the nuclear data. Average distance was between 0.0 and 0.38. The largest group contained 31 accessions including all ploidy levels excluding diploids. The second largest group included five diploids. The lower group included one triploid (108-3) and one tetraploid (91-4) along with C. transvaalensis and 21-2.
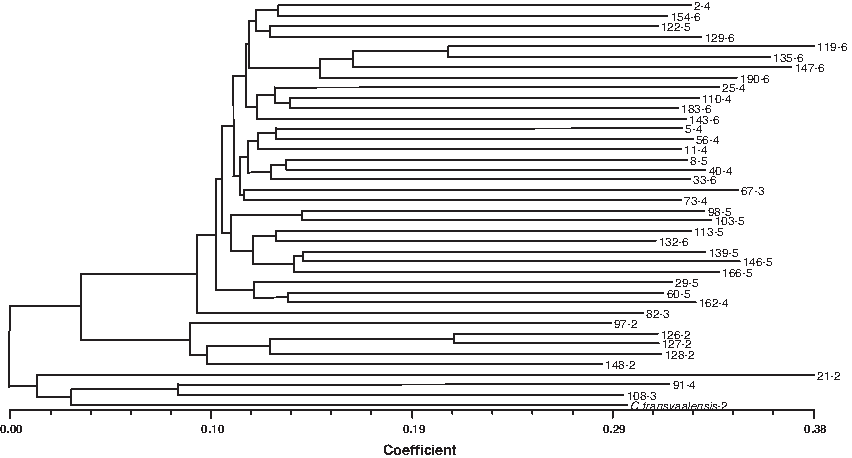
Average distance-based neighbor joining tree of the 40 Cynodon accessions using 407 nuclear DNA markers previously reported by Gulsen et al. (2009).
The PCA was performed to obtain better resolution among the 40 Cynodon accessions (Fig. 5). The first three eigen vector revealed 82% of the total variation observed in the Cynodon accessions, which means that it was reliable. The largest group included 30 accessions including all ploidy levels, excluding diploids. The second largest group included five accessions, one hexaploid (147-6) along with four diploids (126-2, 127-2, 128-2, 148-2). The PCA analysis revealed an intermediate group between the two main groups. This intermediate group included two diploids (C. transvaalensis and 97-2) and one tetraploid (91-4).
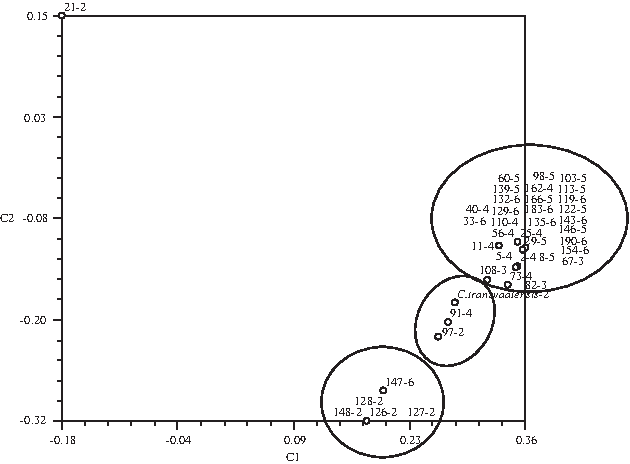
Principle component analysis of 40 Cynodon accessions using two-dimensional scatter plot based on 145 cpDNA RFLP and 16 cpSSR markers. C. transvaalensis is abbreviated with 201-1.
Association between cpDNA-based and nuclear DNA-based data was estimated by using the MXCOMP module nested in NTSYS software. The Mantel test indicated that correlation coefficient (r) between two data sets was 0.70. This level of association was meaningful, indicating moderate association between the two data sets.
Discussion
In this study we aimed genetic analysis of turf type Cynodon accessions with varying ploidy levels. Thus, this was the first study to clarify genetic relationships among Cynodon genetic accessions with ploidy series. We studied 5 loci with 10 restriction enzymes for cpDNA RFLPs and ten loci for cpDNA SSRs of the 40 Cynodon accessions (Tables 1 and 2 and Fig. 1). The samples included diploids, triploids, tetraploids, pentaploids, and hexaploids, and this study aimed to resolve relationships among the ploidy series of the Cynodon accessions. The results were valuable for both bermudagrass breeding programs and understanding of its evolution.
CpDNA-RFLP-based NJ tree estimated that the largest group consisted of all ploidy levels from triploid to hexaploid, excluding diploid. The second largest group included five diploids and one triploid. The cpDNA SSR-based NJ analysis indicated slightly different results. Average distance among the accessions ranged from 0.0 to 0.41. The largest group included 21 accessions with ploidy levels except for diploids. The second largest group consisted of four diploids and one hexaploid, which were not distinguished from each other. C. transvaalensis, accessions 97-2 and 21-2 were intermediate between these two groups. The two-dimensional PCA based on organelle genome markers supported the findings above. The largest group included 30 accessions including all ploidy levels, excluding diploids. The second largest group included five accessions, one hexaploid (147-6) along with four diploids (126-2, 127-2, 128-2, 148-2). Intermediate group between the two main groups included two diploids (210-2 and 97-2) and one tetraploid (91-4). From our previous study (Gulsen et al., 2009) of 192 accessions, nuclear data of the 40 accessions were reused to construct NJ-based dendrogram (Fig. 4). The similar results to organelle DNA analyses were obtained from the nuclear data and average distance was between 0.0 and 0.38. The largest group contained 31 accessions including all ploidy levels excluding diploids. The second largest group included five diploids. The third group included two diploids (C. transvaalensis-2, 21-2, 108-3, 91-4).
Both organelle and nuclear genome analysis produced consistent grouping pattern. Based on these findings, we can hypothesize that (1) diploids were highly different from the triploids, tetraploids, pentaploids, and hexaploids; (2) with assumption of autoploidy, after polyploidization higher ploidy levels had rapid diversification resulting in increased genetic distance between diploids and the other ploidy levels. Adams and Wendel (2005) discussed that many polyploids experience extensive and rapid genomic alterations, some arising with the onset of polyploidy. The results of both nuclear and organelle genome data of the Cynodon accession were consistent with their suggestion.
The cpDNA analyses were reproducible (Fig. 1) and provided DNA markers that were useful in analyzing the maternal genome and phylogenetic relationships among the Cynodon accessions (Figs. 2 and 3). Based on organelle DNA markers, 3 and 6 out of the 40 Cynodon accessions were differentiated in this, and Karaca et al. (2002) discriminated similar proportion (3 out of 31) of forage type bermudagrasses. This may indicate similar organelle DNA polymorphism among the forage type bermudagrasses. In addition, Gulsen et al. (2009) discriminated all accessions from each other based on nuclear genome markers. It can be concluded that the differentiation power of the cpDNA analyses was lower than that of the nuclear genome markers as reported by Karaca et al. (2002) and Gulsen et al. (2009). This is likely due to uniparental inheritance of organelle genome in most plants (Corriveau and Coleman, 1988) and more frequent occurrence of chromosomal recombination and higher rate of point mutations in nuclear genome DNAs) (Curtis and Clegg, 1984).
Annealing temperatures were 60°C for all cpDNA RFLP primers and PCR fragments were highly reproducible. We obtained similar results with cpDNA SSR polymorphism. This indicates that organelle DNA primers were readily transferable (Tables 2 and 3).
Association between cpDNA-based and nuclear DNA-based data was estimated by using the MXCOMP module nested in NTSYS software. The Mantel test indicated that the correlation coefficient (r) between two data sets (cpDNA and nuclear DNA markers) was 0.70. This means moderate association between two data sets. Karaca et al. (2002) found lower Mantel correlation coefficiencies (0.21–0.27) between matrices based on cpDNA (cpDNA-SSR) and nuclear DNA markers (AFLP, RAPD, and DAMI markers). This may be caused by differential genotype and genome sampling by two studies. For example, turf-type genotypes were sampled in our study, whereas forage-type bermudagrasses were used in their study.
Total length of the amplified region spans was 9,230 bp, and this corresponds to about 5% of the chloroplast genome. The cpDNA RFLP primers used in this study amplified 9,230 bp of cpDNA of bermudagrass and had similar sizes in Citrus and buffalograss (Gulsen and Roose, 2001; Gulsen et al., 2005). This suggests cpDNA RFLP regions in these three species were highly conserved in size. However, cpDNA polymorphism was considerably higher in buffalograsses compared to bermudagrasses. The polymorphism level in buffalograsses was 84%, whereas it was only 37% in bermudagrasses. These results indicated that bermudagrasses had narrower cpDNA diversity compared to buffalograsses. This result may also be an indication of later polyploidization of bermudagrass in their evolution, most probably due to molecular o'clock theory in which the level of diversity increases over time.
Accession 21 was the most different as estimated by both organelle and nuclear DNA studies and two statistical procedures. Its morphological characteristics were also different, having longer internodes and slower growth rate and density.
We previously suggested that existence of diploid genotypes supports this area as evolutionarily active as stated by previous researchers (Gulsen et al., 2009). In this study cpDNA RFLP and SSRs also supported this statement. The Cynodon accessions used in this study included all ploidy series from diploids to hexaploids. The diploid bermudagrasses indicated little polymorphism based on cpDNAs. Accession 21-2 and C. transvaalensis based on cpDNA RFLP and accession 21-2, 97-2 and C. transvaalensis based on cpDNA SSR markers were distinguished from the other diploid bermudagrasses. The nuclear DNA analysis differentiated all diploids, and diversity among the diploids was similar to the samples with the other ploidy levels.
Several different studies about chloroplast microsatellites have shown considerable levels of diversity compared to those of chloroplast RFLPs (Provan et al., 1997). A fourfold increase in levels of diversity was revealed when the same species were studied using chloroplast microsatellites in a study carried out on a range of rice species, which had previously been examined using chloroplast RFLPs. Likewise, a study on barley (Hordeum vulgare) revealed six chloroplast microsatellite haplotypes among 12 H. vulgare ssp. spontaneum accessions, whereas a previous RFLP study on 245 samples generated only three haplotypes (Provan et al., 1999).
CpDNA RFLP and SSR results were evaluated separately and together to show phylogenetic relationships among the Cynodon accessions with varying ploidy levels. The number of branches in a phylogenetic tree can be used as an indication of detection power of polymorphism. cpDNA RFLP and cpDNA SSR polymorphism estimated 6 and 11 haplotypes based on the NJ tree, respectively. This result was consistent with the findings above. The joint data analysis of cpDNA RFLP and SSR markers based on PCA detected 11 different haplotypes, identical to cpDNA RFLP haplotypes.
Conclusion
Harlan and De Wet (1969) suggested that an area from West Pakistan to Turkey is the center of evolutionary activity, and Gulsen et al. (2009) reported all ploidy series in this region. In this study we combined two cpDNA (cpDNA RFLP and SSR) and four nuclear DNA (SRAP, POGP, ISSR, and RAPD) markers to clarify relationships among the ploidy series of the 40 Cynodon accessions. Thus, the results were likely to be conclusive: (1) the diploids were the most different from the rest of the ploidy levels, and (2) assuming autopolyploidy in bermudagrasses, higher ploidy levels than the diploids went rapid diversification after polyploidization. This study was the most comprehensive to deduce relationships among the ploidy series of the Cynodon accessions.
Footnotes
Acknowledgments
This research was supported by the Scientific and Technical Research Council of Turkey under the project code TUBITAK TOVAG: 105 O 586 and Erciyes University Scientific Research Project under the project code FBY9-1117.
Author Disclosure Statement
The authors declare that no competing financial interests exist.