
Abstract
Abstract
Urodele amphibians such as the axolotl (Ambystoma mexicanum) display a large capacity for tissue regeneration and remarkable resistance to cancer. As a model organism, axolotl thus offers a unique opportunity for cancer research and anticancer drug discovery, not to mention the discerning mechanisms that underpin controlled cellular growth and regeneration versus cancer. To the best of our knowledge, little is known on comparative gene expression changes during regeneration events such as wound healing in axolotl and humans. Using publicly available transcriptomics data and bioinformatics analyses, we examined the differential gene expression signatures in skin wound samples from axolotl and humans after skin biopsy punch injury, in comparison with intact (uninjured) control skin samples. We identified 95 genes exhibiting a reversal expression pattern between humans and axolotl during the wound healing/regeneration period. These genes were significantly associated with collagen biosynthesis, extracellular matrix organization, PI3K-Akt signaling pathway, immune system response, and apoptotic process. Furthermore, this new gene set exhibited high prognostic performance in discriminating the survival risk in skin-related cancers, including melanoma (hazard ratio [HR] = 8.14, p < 10−30), oral cancer (HR >100, p < 10−12), and head and neck carcinoma (HR = 5.29, p < 10−30). Moreover, considering these gene signatures, we repositioned 11 small molecules as potential anticancer drug candidates indicating reversal effects on upregulated human genes and downregulated axolotl genes or mimicking downregulated human genes and upregulated axolotl genes. We anticipate that this study offers new insights on gene signatures bridging regeneration mechanisms with tumorigenesis and cancer drug repositioning.
Introduction
The initial concepts that regeneration mechanisms and controlled cellular growth might offer clues on cancer biology and uncontrolled cellular processes have emerged in the first half of the 20th century (Waddington, 1935). Regeneration indicates a well-organized restoration of body parts that were lost functionally or physically. Thus, regeneration including proliferation necessitates signals for regulation of cell cycle to undergo cell cycle division and repair (Oviedo and Beane, 2009; Sánchez Alvarado, 2000). Regeneration of engaged cells are absolutely mentored to required zones, and after regeneration is finished, explicit prompts are necessary to report regenerative achievement and signal completion. Yet, under other conditions, proliferation may proceed toward uncontrolled growth, resulting in unwanted consequences for homeostasis (Oviedo and Beane, 2009).
Of note, one theory about regeneration is that if morphogenetic processes supersede the regeneration-related cellular proliferation, then regeneration is likely to inhibit abnormal growth and even reverse malignancies (Fior, 2014; Oviedo and Beane, 2009).
Urodele amphibians such as the axolotl (Ambystoma meaxicanum) display a large capacity for tissue regeneration and remarkable resistance to cancer. As a model organism, axolotl thus offers a unique opportunity for cancer research and anticancer drug discovery, not to mention the discerning mechanisms that underpin controlled cellular growth and regeneration versus cancer (Amamoto et al., 2016; Bryant et al., 2017; Chernoff et al., 2018; Erickson et al., 2016; Fior, 2014; Haas and Whited, 2017; Milyavsky and Dickie, 2017; Needham, 1936; Saad et al., 2018; Tazaki et al., 2017; Yokoyama et al., 2018).
Little is known on comparative gene expression changes during regeneration events such as wound healing in axolotl and humans. In the present study that harnesses publicly available transcriptomics data and bioinformatics analyses, we report the differential gene expression signatures in skin wound samples from axolotl and humans after skin biopsy punch injury, in comparison with intact (uninjured) control skin samples. In addition, we report our findings with a view to cancer drug repositioning by using a dataset of chemically perturbed expression profiles (Fig. 1).
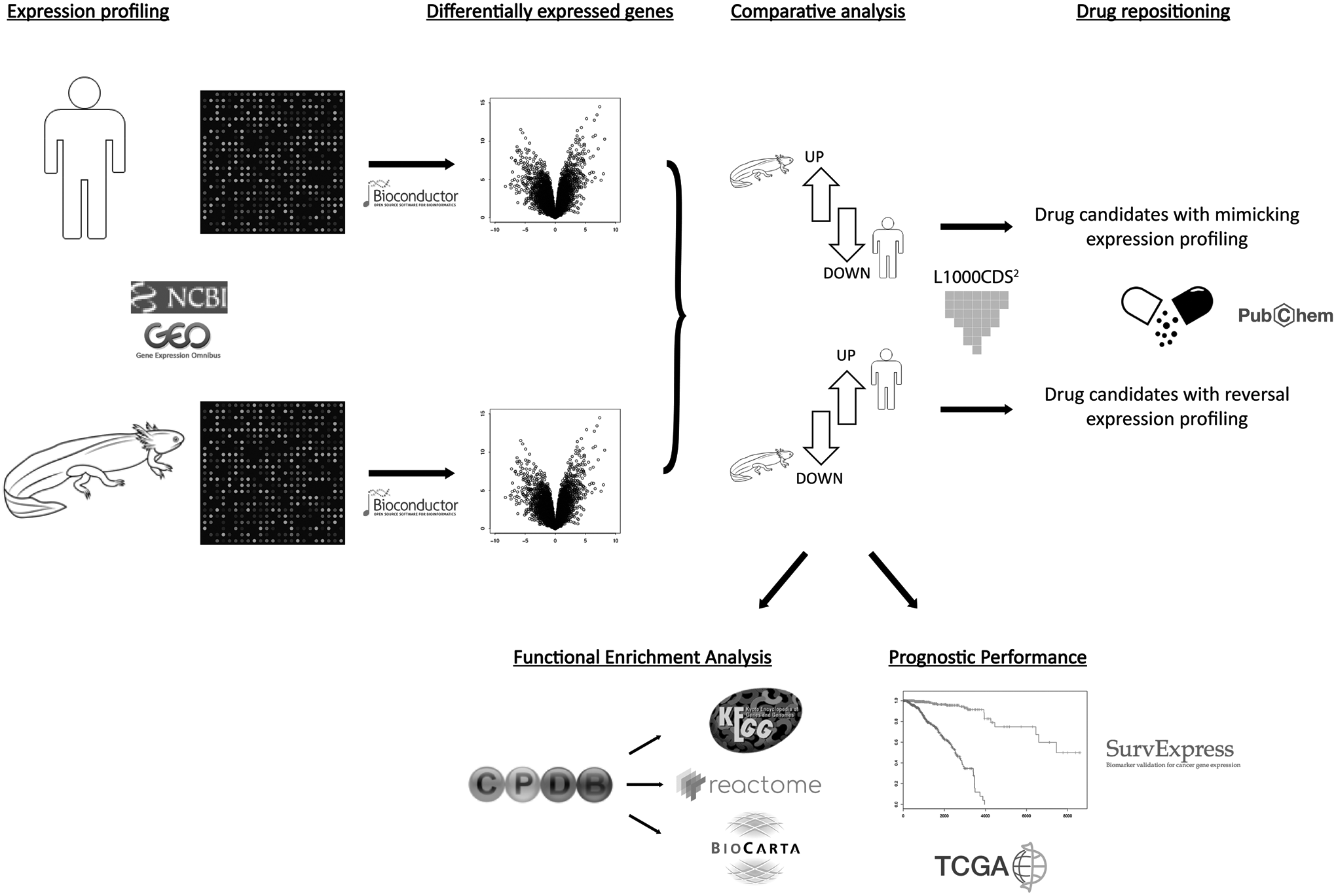
Flowchart on the study design. Gene expression datasets related to human and axolotl skin injury at three time points after injury (days 3, 7, and 14) were obtained from the Gene Expression Omnibus database. Each dataset was analyzed statistically using Bioconductor platform and the differentially expressed genes and their directions of regulation were identified through volcano plots. Comparative analyses were conducted to identify genes with reversal expression profiles between two organisms. To identify potential anticancer drugs, a drug repositioning algorithm was applied. Significant drugs with reversal effects on upregulated human gene signatures and downregulated axolotl gene signatures or mimicking downregulated human gene signatures and upregulated axolotl gene signatures, were collected as potential anticancer drug candidates.
Materials and Methods
Axolotl and human microarray data
Axolotl and human skin datasets are available online through the Gene Expression Omnibus (GEO) Database (Barrett et al., 2013). We used the GSE35255 dataset for axolotl that included aquatic axolotl skin samples at day 3 (A3 group—four samples) and day 7 (A7 group—four samples) after skin biopsy punch injury, and four uninjured skin samples as control group (Seifert et al., 2012). The other axolotl dataset (GSE79299) included axolotl skin biopsy punch injured samples at day 14 (A14—three samples) and three uninjured samples as control (Erickson et al., 2016).
The human dataset, GSE28914, consisted of skin graft donor site wound healing samples taken at day 3 (H3 group—six samples) and day 7 (H7 group—five samples), and eight uninjured samples as control group (Nuutila et al., 2012). The other human dataset (GSE50425) included skin wound biopsy samples taken at day 14 (H14 group—four samples) and four uninjured control samples (Table 1).
Transcriptome Datasets Used in this Study
GEO, Gene Expression Omnibus.
Preprocessing of transcriptome datasets and differential gene expression analysis
A well-established statistical analysis procedure (Gov et al., 2017; Islam et al., 2018; Kori and Arga, 2018) was adopted in this study to analyze microarray data. The raw data for Affymetrix datasets (GSE28914, GSE79299, and GSE35255) were read into R statistical software using the Affy package (Gautier et al., 2004) and normalized using the Robust multiarray average (Bolstad et al., 2003) implemented in Bioconductor platform (version Rx64 3.0.2) (Gentleman et al., 2004). Data from Illumina dataset (GSE50425) were preprocessed using GEOquery package (Sean and Meltzer, 2007) and quantile normalization was used. The normalized expression levels of genes were compared by using Linear Models for Microarray Data (LIMMA) package (Smyth, 2005) to identify differentially expressed genes (DEGs). Benjamini–Hochberg method was used as to control the false discovery rate.
Axolotl microarray IDs were matched to human orthologous genes with GEOquery package. Each time point of each organism was compared with its own control (uninjured) sample group statistically and to determine the statistical significance of the orthologous DEGs, an adjusted p-value threshold of ≤0.05 (or ≤0.01) and fold-change cutoff ≥2 (for upregulation) or ≤0.5 (for downregulation) were used. Principle component analysis (PCA) was performed to analyze differential clustering of human and axolotl samples taken at different time points. Venn diagrams and heat maps were created using Jvenn plug-in (Bardou et al., 2014) and pheatmap R package (Kolde, 2019), respectively.
Functional enrichment analysis
The functional enrichment analysis of the gene sets was performed by ConsensusPathDB (Kamburov et al., 2013). In Gene Ontology (GO) overrepresentation analyses, GO terms at levels 4 and 5 were screened and enriched GO terms with p ≤ 0.01 were accepted as statistically significant. Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa et al., 2019; https://www.kegg.jp/) BioCarta (https://cgap.nci.nih.gov/Pathways/BioCarta_Pathways) and REACTOME (Fabregat et al., 2018; http://reactome.org) databases were used as the data resource in pathway enrichment analyses, and p ≤ 0.05 was used to describe statistical significance.
Preclinical validation of prognostic performance of the modules
The prognostic capabilities of the gene signatures were identified using RNA-Seq dataset for melanoma (n = 335), oral cancer (n = 86), and head and neck cancer (n = 502) obtained from TCGA (Weinstein et al., 2014) and Cox proportional hazards regression analysis by SurvExpress validation tool (Aguirre-Gamboa et al., 2013). In SurvExpress, the samples were partitioned into low- and high-risk groups according to their prognostic index. The prognostic capabilities of the gene signatures were characterized through Kaplan–Meier plots, hazard ratios (HR), and the log-rank test p-values. In addition, prognostic performances of the gene signatures were investigated across TCGA-originated RNA-Seq datasets associated with other cancers including breast cancer (n = 962), ovarian cancer (n = 247), colorectal cancer (n = 466), lung cancer (n = 475), liver cancer (n = 361), and prostate cancer (n = 497).
Drug repositioning based on mimicking or reversal of gene signatures
To identify potential candidate drugs, L1000CDS2 (LINCS L1000 characteristic direction signature search engine) was used (http://amp.pharm.mssm.edu/L1000CDS2). L1000CDS2 enables to find consensus L1000 small molecule signatures (Subramanian et al., 2017) that match user input signatures (i.e., down and upregulated genes). The Kolmogorov–Smirnov statistic and random permutation tests were applied to calculate the similarity scores and statistical significance (p-value) of drugs. Significant drugs (p ≤ 0.05), with reversal effects on upregulated human gene signatures and downregulated axolotl gene signatures or mimicking downregulated human gene signatures and upregulated axolotl gene signatures, were collected as potential anticancer drug candidates in this study. Information on drugs (indication, mechanism of action, etc.) was based on NCBI-PubChem database (Kim et al., 2019).
Results
Differential gene expression profiles in human and axolotl skin biopsies
To display gene expression alterations during regeneration in humans and axolotl, gene expression levels in injured skin samples were analyzed at three different time points (i.e., at days 3, 7, and 14) and compared with those in uninjured skin samples in each organism. Volcano plots were constructed at each time point and DEGs were identified considering adjusted p-value and fold change thresholds (Fig. 2). The number of DEGs was decreasing over time, whereas the upregulated and downregulated genes were almost evenly distributed at each time point (Table 2).
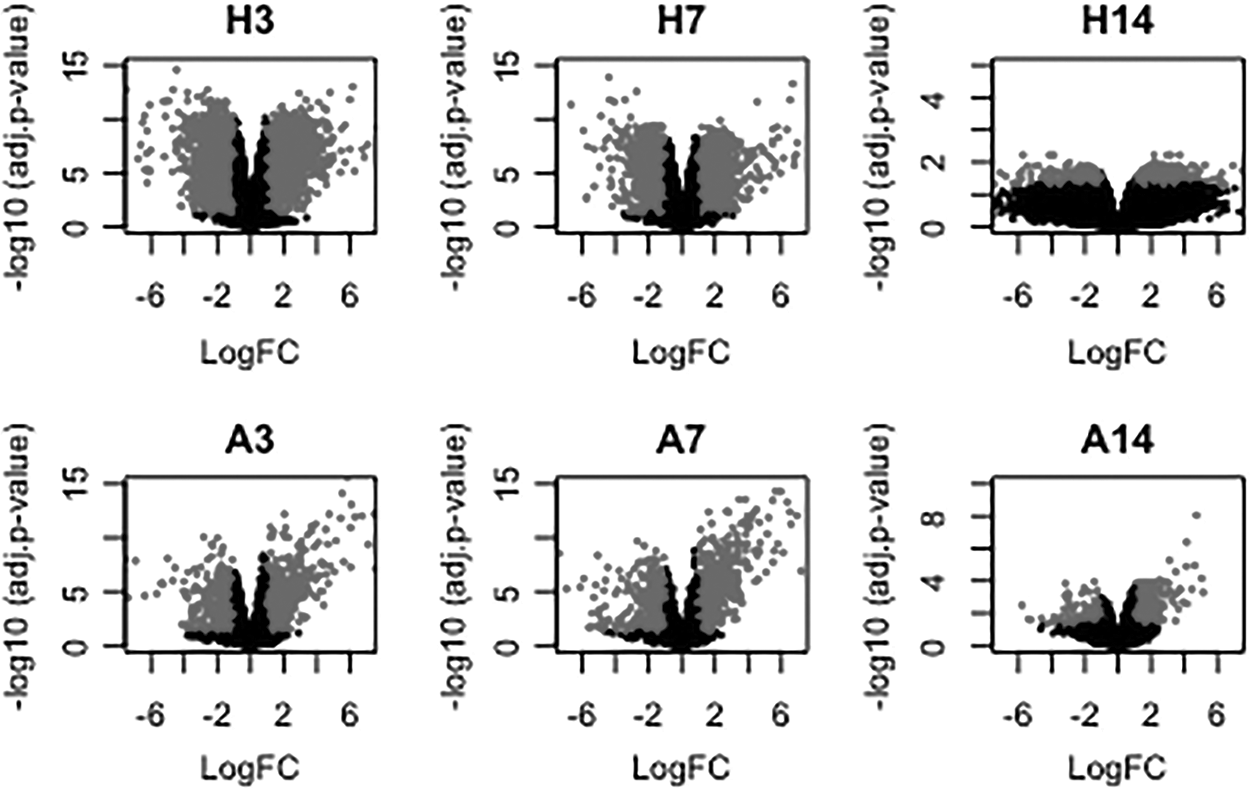
Volcano plots displaying differential gene expression profiles at each time point. H3, human skin profile at day 3; H7, human skin profile at day 7; H14, human skin profile at day 14; A3, axolotl skin profile at day 3; A7, axolotl skin profile at day 7; A14, axolotl skin profile at day 14; FC, fold change.
Number of Differentially Expressed Genes at Each Time Point (Days 3, 7, and 14) in Humans (H) and Axolotl (A)
DEGs, differentially expressed genes.
Comparative analysis of gene expression alterations between humans and axolotl
To identify the similarities and differences in gene expression alterations during regeneration in humans and axolotl, we comparatively analyzed the homologous genes, which were differentially expressed in humans and axolotl during the 14-day period. Comparative analysis of DEGs in axolotl and humans at each time point indicated that only a small portion of the DEGs (between 2.6% and 5.4%) were mutually expressed in two organisms at any time point (Fig. 3). Among these mutual DEGs, the majority (65.6% at day 3, 71.9% at day 7, and 87.0% at day 14) represented alterations in two organisms in the same regulatory direction (i.e., upregulated or downregulated in both organisms). On the contrary, we observed that a total 95 DEGs exhibited a reversal expression pattern between humans and axolotl during the whole time period (Fig. 4). At day 3, 69 DEGs exhibited a reversal expression pattern between humans and axolotl. Among those, 31 genes were downregulated in humans and upregulated in axolotl.
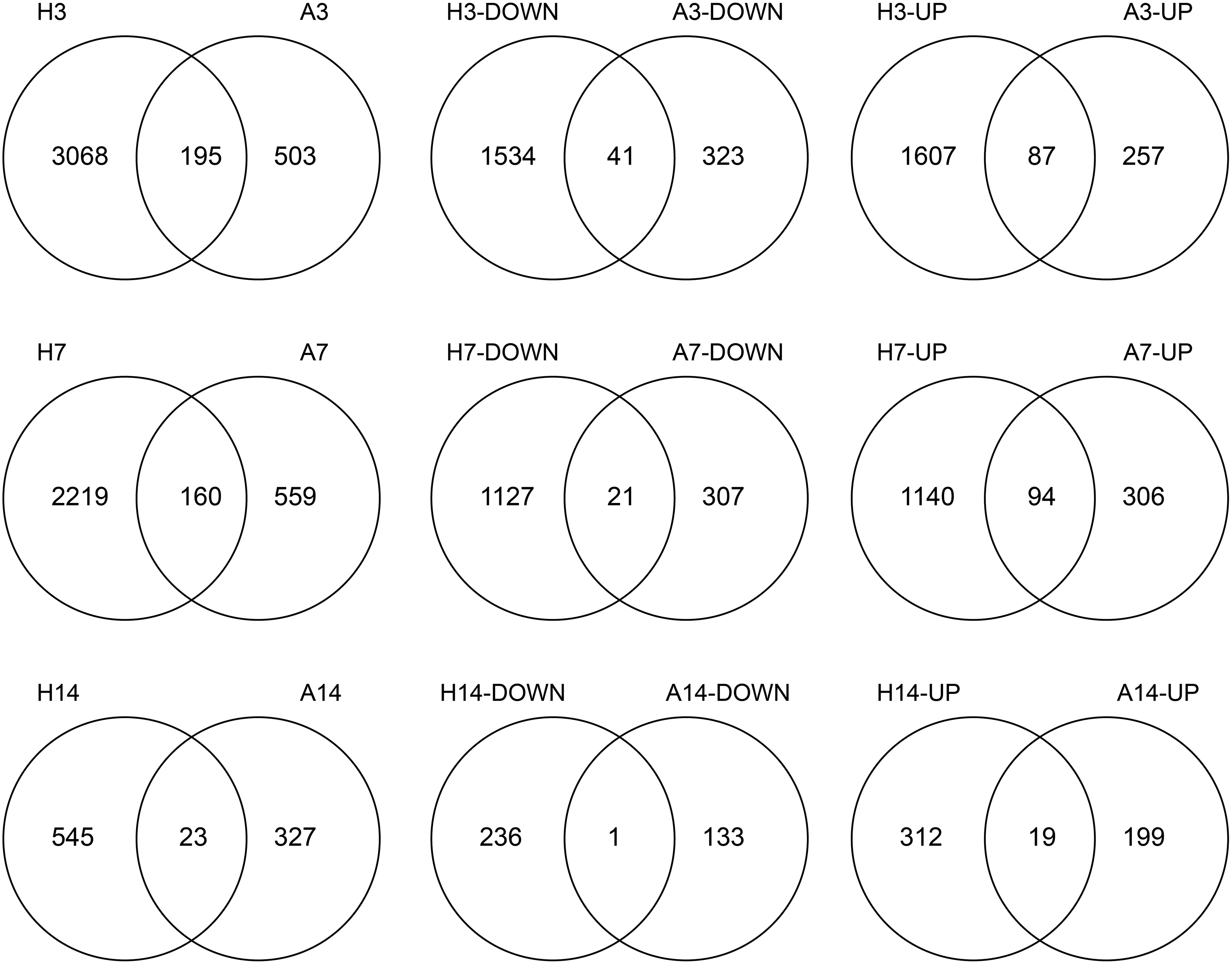
Venn diagrams of differentially expressed genes in each comparison.
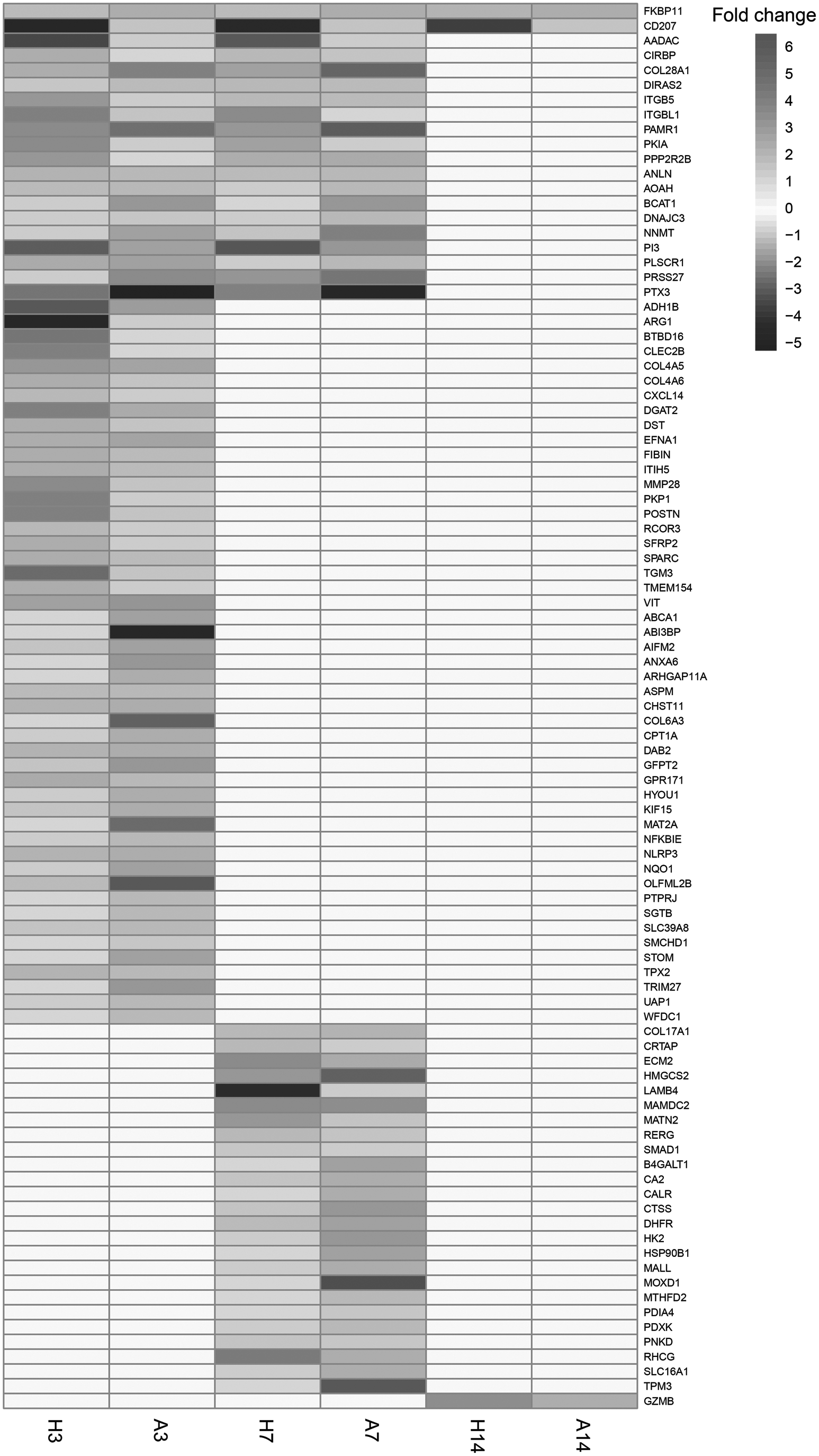
Heat map of 95 homologous genes exhibiting a reversal expression pattern between humans and axolotl. H3, human skin profile at day 3; A3, axolotl skin profile at day 3; H7, human skin profile at day 7; H14, human skin profile at day 14; A7, axolotl skin profile at day 7; A14, axolotl skin profile at day 14.
On the contrary, 38 genes were acting in reverse manner (i.e., upregulated in humans, and downregulated in axolotl). At day 7, the number of DEGs exhibiting a reversal expression pattern between two organisms decreased to 45, among which 19 genes were downregulated in humans and upregulated in axolotl, and 26 genes were acting in reverse manner. At day 14, the differences in expression patterns almost disappeared, because a single gene (CD207) was downregulated in humans and upregulated in axolotl, whereas only two genes (FKBP11 and GZMB) were upregulated in humans and downregulated in axolotl (Fig. 4).
We also performed a PCA taking into consideration the expression levels of 95 genes to analyze the discrimination efficiency of these genes in clustering the humans and axolotl samples at different time points. Considering the first two principle components (PC1 and PC2, explaining the 76% the 17% of the total variance, respectively), we showed a clear separation of axolotl and human samples at days 3 and 7 (Fig. 5). The samples at day 14 were clustered together as expected, because the differences in expression patterns almost disappeared at day 14 (Fig. 4).
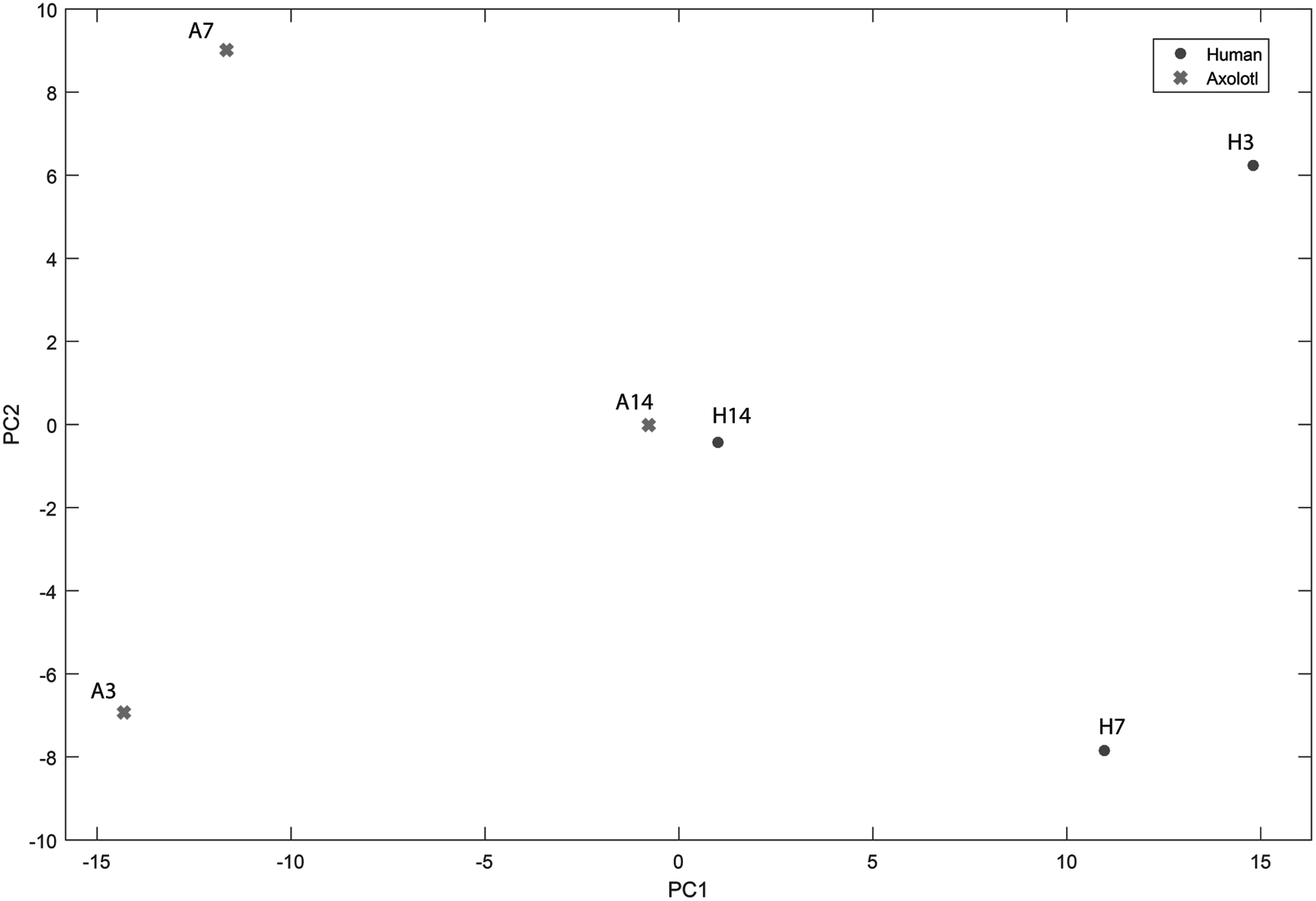
Plot of principle component analysis based on expression profiles of 95 genes in axolotl and human samples. H3, human skin profile at day 3; H7, human skin profile at day 7; H14, human skin profile at day 14; A3, axolotl skin profile at day 3; A7, axolotl skin profile at day 7; A14, axolotl skin profile at day 14.
Biological evaluation of differential gene signatures
The 95 DEGs exhibiting differential regulatory pattern among axolotl and humans were further analyzed in terms of their roles in biological processes and molecular pathways. Several pathways associated with collagen biosynthesis, extracellular matrix (ECM) organization, PI3K-Akt signaling pathway, and immune system response were significantly enriched (Table 3). Furthermore, GO term enrichment analysis indicated association of these genes with ECM organization, chondrocyte differentiation and development, oxoacid and coenzyme metabolic processes, development and morphogenesis of connective tissue, bone and skeletal system, secretion, cell adhesion and migration, response to nutrient levels such as lipids, immune system response, regulation of transport, and apoptotic process (Table 4).
Significantly Enriched Pathways Associated with 95 Gene Signatures (Number of Associated Genes ≥5, Adjusted p ≤ 0.05)
ECM, extracellular matrix; KEGG, Kyoto Encyclopedia of Genes and Genomes.
Significantly Enriched Biological Processes Associated with 95 Gene Signatures (Number of Associated Genes ≥5, Adjusted p ≤ 0.05)
Among the 95 genes, 11 genes were previously identified as either oncogene or tumor suppressor gene (TSG) by comparing them with the TSG (Zhao et al., 2013, 2016) and oncogene (Liu et al., 2017) lists in literature. According to this comparison, the tumor suppressors SPARC and TGM3 were downregulated in humans and upregulated in axolotl at day 3, whereas there were no significant differences in their expression levels at days 7 and 14.
Similarly, among TSGs, ARG1, CXCL14, and SFRP2 were underexpressed in early phase (day 3) in humans, and overexpressed in axolotl. Other TSGs, namely disabled homolog 2 (DAB2), methionine adenosyltransferase II alpha (MAT2A), PTPRJ, SMCHD1, and WFDC1 represented a reversal profile, that is, they were upregulated in humans and downregulated in axolotl at day 3. Oncogenes and TSGs were not differentially expressed at days 7 and 14 either in humans or axolotl, with a single exception being HSP90B1, which was upregulated in humans and downregulated in axolotl at day 7 (Fig. 6).
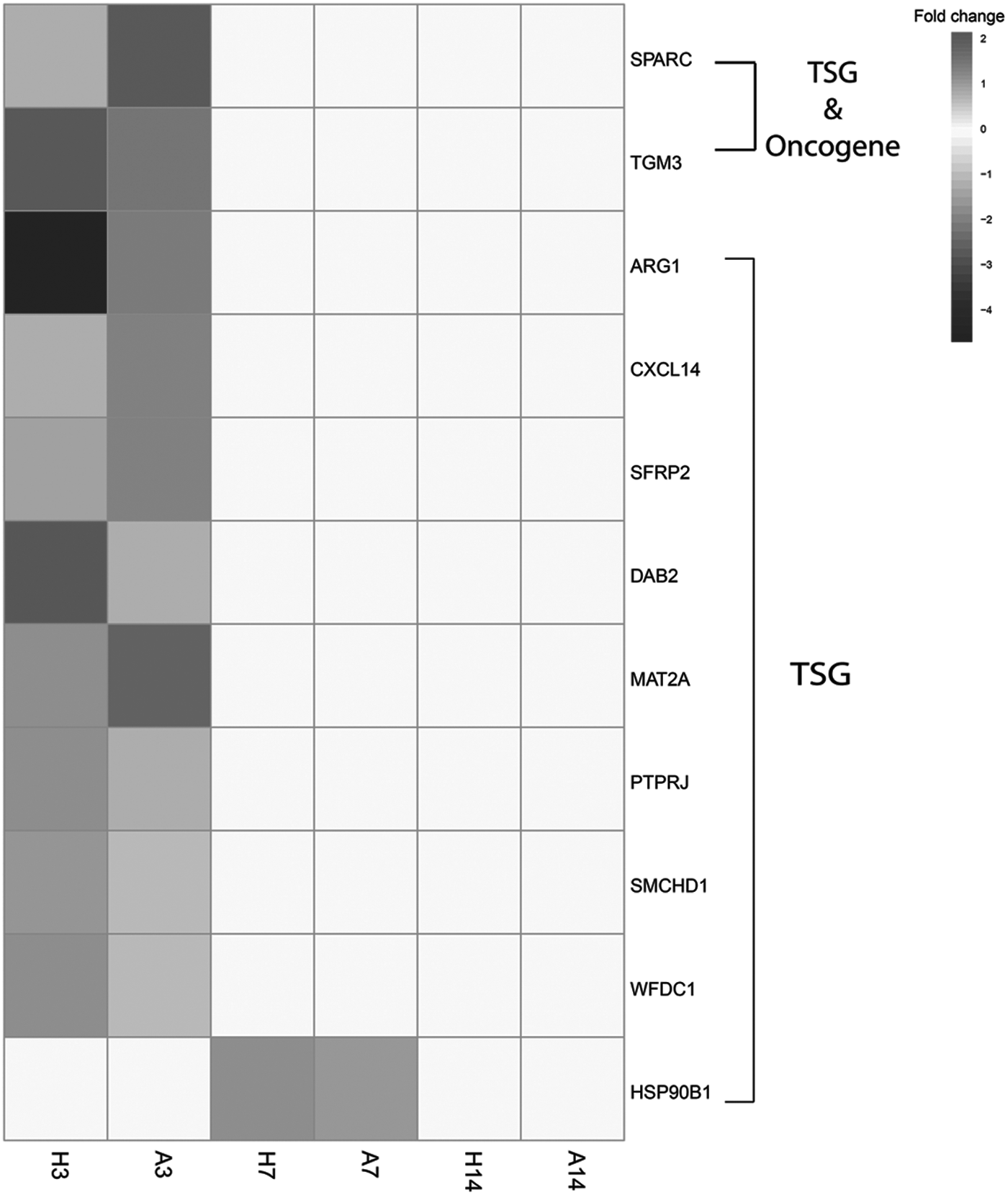
Heat map showing the expression levels of oncogenes and tumor suppressor genes in axolotl and human samples. H3, human skin profile at day 3; H7, human skin profile at day 7; H14, human skin profile at day 14; A3, axolotl skin profile at day 3; A7, axolotl skin profile at day 7; A14, axolotl skin profile at day 14; TSG, tumor suppressor gene.
The prognostic performance of these 95 gene signatures were also investigated in several human cancers using TCGA-originated RNA-Seq datasets through disease-free survival and Cox proportional hazards regression analyses (Fig. 7). The gene set indicated high performance in discriminating the survival risk in melanoma (HR = 4.86), oral cancer (HR = 3.04), and head and neck carcinoma (HR = 3.25). Furthermore, the prognostic performance of this gene set was not specific to these skin cancers, because the gene set also indicated significantly high performance (p < 10−3) in discriminating the survival risk in breast cancer (HR = 7.06), ovarian cancer (HR = 7.36), colorectal cancer (HR = 5.61), lung cancer (HR = 4.54), liver cancer (HR = 4.68), and prostate cancer (HR >100).
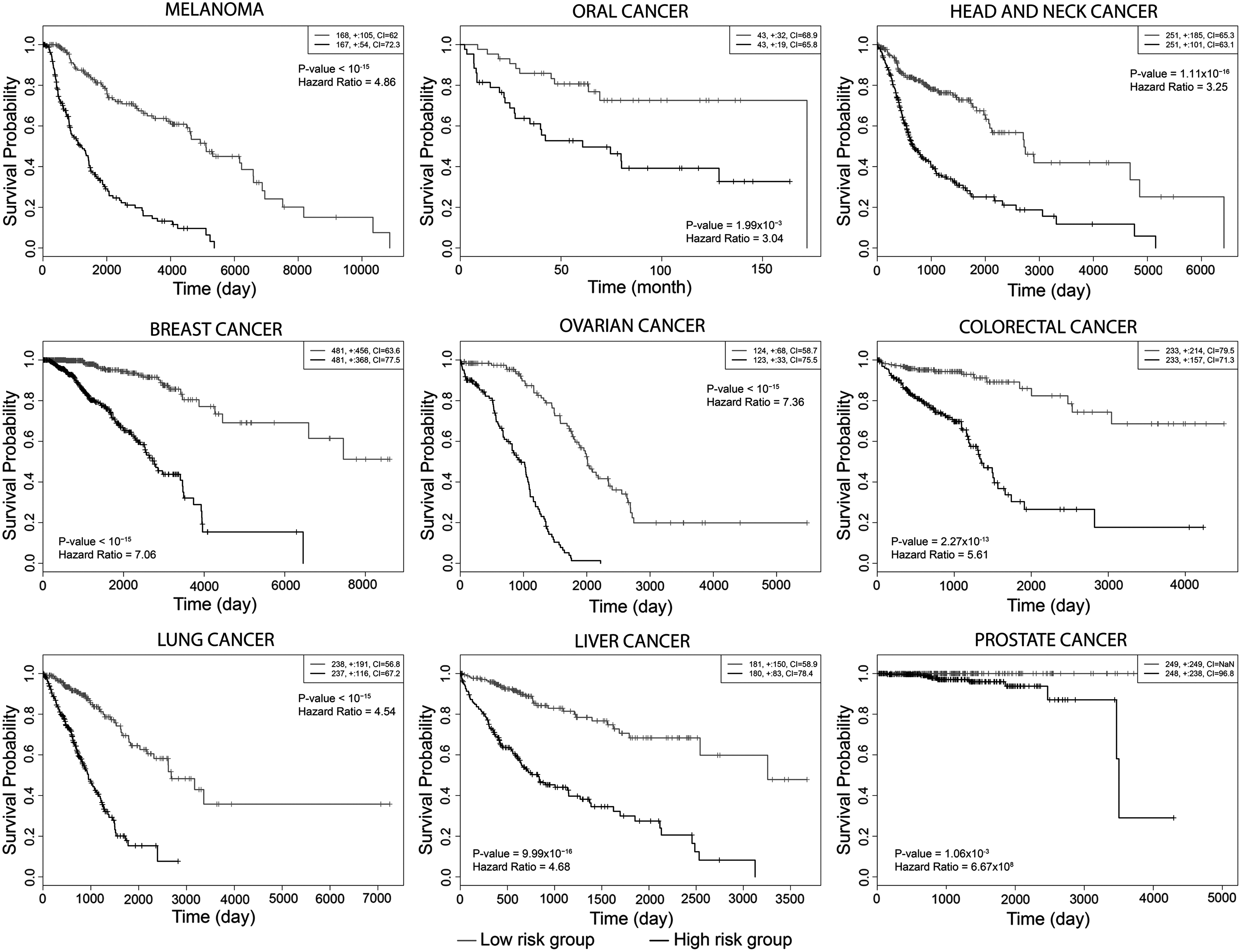
Kaplan–Meier plots representing the prognostic performance of 95 genes in several human cancers.
Drug repositioning based on reversal gene signatures
To identify potential candidate drugs, we used the L1000 dataset, which include the chemically perturbed expression profiles for 33,197 mRNAs in 62 unique cell lines in response to 3924 different small molecules (Subramanian et al., 2017). Using the L1000CDS2 characteristic direction signature search engine and 95 gene signatures with reversal expression profiles between humans and axolotl, 40 drugs indicating reversal effects on upregulated human gene signatures and downregulated axolotl gene signatures or mimicking downregulated human gene signatures and upregulated axolotl gene signatures with overlap ≥0.08 were identified (Table 5). The majority (29 of 40) of the drugs were antineoplastic agents that were already proposed for treatment of cancers.
Repositioned Drug Candidates (Overlap ≥0.08)
On the contrary, BJM-ctd2-9 (targeting the metabolism of xenobiotics by cytochrome P450), UNC0321 (epigenetic regulator of frataxin), CGP-60474 (antiviral used in inhibition of Flavivirus infection), PP-110 (inhibitor of Src and Abl protein tyrosine kinases), AG14361 (antigout and antihyperuricemic), BRD-K35716340 (antibiotic), aminopurvalanol A (antiasthmatic), emetine (antiemetic and anthelminthic), BRD-K19724398, HG-5-113-01, and tremulacin were repositioned as potential anticancer drug candidates in this study.
Discussion
The reality that tumorigenesis is observed at very low frequencies upon carcinogenic exposure to regenerative tissues in animals with highly regenerative capacities including axolotl oriented researchers to elucidate the associations between regenerative mechanisms and tumorigenesis. One theory about regeneration is that if morphogenetic processes supersede the regeneration-related cellular proliferation, then regeneration is likely to inhibit abnormal growth and even reverses malignancies that are commonly detected in animals with highly regenerative capacities (Fior, 2014; Oviedo and Beane, 2009). Thanks to the development of next-generation sequencing technologies, it is now possible to analyze these types of associations at molecular level (Tekir et al., 2009).
Previously, several studies reported genome reprogramming during regeneration of injured skin samples in humans (Nuutila et al., 2012) and axolotl (Erickson et al., 2016; Seifert et al., 2012). However, this process was not compared broadly between axolotl and humans. On the contrary, this type of comparison would give possibility to determine mechanisms and proteins with cancer conferring or cancer protective function, and allow development of new therapeutic strategies.
To explore potential genes that associate regeneration process with cancer biology, we compared the gene expression profiles in skin samples after skin biopsy punch injury with respect to uninjured skin samples in axolotl and human at three different time points during 14-day period. We observed that number of DEGs exhibiting a reversal expression pattern between humans and axolotl (downregulated in humans and upregulated in axolotl, or upregulated in humans and downregulated in axolotl) were decreasing with time and the differences in expression patterns almost disappeared at day 14.
Such a reversal expression pattern between two organisms was characterized by 95 DEGs during the whole regeneration period. These gene signatures were associated with several biological processes and molecular pathways such as collagen biosynthesis, ECM organization, PI3K-Akt signaling pathway, and immune system response, which were already associated directly or indirectly with cancer hallmarks (Fang et al., 2014; Gonzalez et al., 2018; Martini et al., 2014; Walker et al., 2018). Furthermore, 11 of them were previously identified as either oncogene or tumor suppressor. All these results together with high prognostic performances of these genes in discriminating the survival risk in human tumors (such as melanoma, oral cancer, and head and neck cancer) supported our hypothesis that distinct gene signatures during the regeneration period in humans and axolotl may be associated with regulation of tumorigenesis.
In our study, CD207 protein was found to be downregulated at each time point for humans, whereas it was upregulated at each time point for axolotl. CD207 (Langerin, NP_056532) is a type II transmembrane protein that is abundantly expressed in Langerhans—the epidermal dentritic cells specialized in antigen presentation (Shevchuk et al., 2014) and can promote tumor-specific immunity (Benencia et al., 2012; Dyduch et al., 2017). Langerhans cells are distinct with their Birbeck granules and CD207 is an endocytic receptor that has a potential inducer role in Birbeck granule formation (Valladeau et al., 2000). The mutations in CD207 gene result in inhibition of Birbeck granules that make it a precious molecule in skin cancers.
Previously, this molecule was found to be expressed considerably lower in basal cell carcinomas compared with normal human epidermis (Plzáková et al., 2004), and again lower in cutaneous basal and squamous cell carcinomas compared with normal skin (Shevchuk et al., 2014). Moreover, malignant skin lesions had a lower number of Langerhans cells compared with benign skin indicating the disparity in immune response pattern about the cutaneous tumors (De Melo et al., 2006).
We also detected that FKBP11 protein, which is a potential early hepatocellular carcinoma (HPC) marker (Lin et al., 2013) is reverse-regulated (up in humans and down in axolotl) at two time points (D7 and D14). This protein was first characterized by Rulten et al. (2006) and its mRNA is plentiful in secretary tissues. Lin et al. demonstrated in their study that FKBP11 expression level rises as the severity of the carcinoma increases. Thus this gene could possibly be an early marker for carcinoma. In addition, this group proposed FKBP11 to be an early response gene to liver damage events that demonstrate ER stress as a result of metabolic disorders (Lin et al., 2013). FKBP11 low expression is correlated with survival rate human melanoma according to human protein atlas (https://www.proteinatlas.org/ENSG00000134285-FKBP11/pathology).
Although secreted protein acidic and rich in cysteine (SPARC) and TGM proteins were found to be tumor suppressors and oncogenes depending on the study in the literature, we propose these two proteins to be tumor suppressors according to our results that indicated both proteins to be downregulated in humans and upregulated in axolotl at our very early time point of regeneration (D3). TGM3 is an epidermal transglutaminase widely expressed in supra basal layer of the stratified squamous epithelium and is fundamental for the epidermal final differentiation (Eckert et al., 2005). In literature, this protein is downregulated in head and neck squamous cell carcinoma and exogenous expression of it prevents the proliferation. It also reinforces the apoptosis of the cancer cells in vitro and represses tumorigenesis in vivo (Wu et al., 2013).
Furthermore, it is downregulated in oral squamous cell carcinoma (Nayak et al., 2018; Negishi et al., 2009) and esophageal carcinomas (Chen et al., 2000; Liu et al., 2006). TGM3 is important in the transformation of the keratinocytes to corneocytes by apoptosis and therefore, downregulation of this protein in carcinomas may intrude on the possibly crucial initiation of apoptosis, thereby supporting tumor cell survival (Uemura et al., 2008).
Similarly, SPARC, which is downregulated in human samples, is upregulated in axolotl samples, is a matrix-associated protein, and is principally expressed in tissues experiencing repair or remodeling (Socha et al., 2015). Yiu and his friends found that this protein leads to apoptosis only in cancer cells, when it is applied to ovarian cancer cells and ovarian surface epithelial cells, demonstrating the downregulation of this crucial for carcinogenesis (Yiu et al., 2001). In addition, in one study, this protein was found to be downregulated in ovarian cancer through aberrant promoter hypermethylation and exogenous addition of this protein inhibited proliferation of cancer cells (Socha et al., 2015).
In our study, the TSGs ARG1, CXCL14, and SFRP2 were underexpressed in early phase (day 3) in humans, and overexpressed in axolotl. ARG1, encoding the protein Arginase 1, is believed to be a crucial metabolic protein encoding gene, responsible for hydrolysis of arginine to ornithine and urea and is important for important metabolic pathways. You et al. (2018) found that ARG1 is underexpressed in HPC and was correlated with prognosis of the patients.
Similarly, the chemokine CXCL14, which inhibits angiogenesis (Kondo et al., 2016), is proved to be silenced by DNA hypermethylation in various cancers such as acute myeloid leukemia (Sonnet et al., 2014), stomach cancer (Hu et al., 2013), colorectal cancer (Cao et al., 2013), and lung cancer (Tessema et al., 2010). Likewise, SFRP2, encoding the secreted frizzled-related protein 2, belongs to the family of Wnt pathway antagonists and is downregulated in human gastric cancer (Cheng et al., 2007), malignant pleural mesothelioma (Cheng et al., 2017), and silenced by hypermethylation in colorectal cancer (Naini et al., 2016).
Among the other TSGs, DAB2, MAT2A, PTPRJ, SMCHD1, and WFDC1 were upregulated in humans and downregulated in axolotl at day 3 and HSP90B1 was upregulated in humans and downregulated in axolotl at day 7. Although DAB2 is respected as a TSG previously, Xie et al. (2015) showed that overexpression of this protein increased cancer cell migration and invasion, regulates the cell migration-related genes, suggesting this protein has a crucial role in prostate cancer and metastasis. Likewise, MAT2A was overexpressed in gastric cancer (Zhang et al., 2013), colon cancer (Chen et al., 2007), and HPC (Liu et al., 2011). The overexpressions of PTPRJ and SMCHD1 genes were not reported in any cancer.
In addition, WFDC1 gene encodes for a whey acidic protein four-disulfide core member protein that its upregulation in a stromal/cancer cell xenograft model in prostate cancer leads to tumorigenesis because of increased angiogenesis (McAlhany et al., 2003). Finally, the heat shock protein 90B1 (HSP90B1) that was overexpressed in humans and underexpressed in axolotl at day 7 was found to be overexpressed in the studies carried out by Xu et al. (2016) in nonsmall cell lung cancer and Cawthorn et al. (2012) in breast cancer. In the latter study, overexpression of this protein was also found to be associated with decreased overall survival and distant metastasis.
Considering the 95 gene signatures as drug candidates and using a comprehensive library of chemically perturbed expression profiles for 3924 different small molecules (Subramanian et al., 2017), we identified 11 novel drugs as potential anticancer therapeutics indicating reversal effects on upregulated human genes and downregulated axolotl genes or mimicking downregulated human genes and upregulated axolotl genes. The anticancer drug candidates were BJM-ctd2-9 (targeting the metabolism of xenobiotics by cytochrome P450), UNC0321 (epigenetic regulator of frataxin), CGP-60474 (antiviral used in inhibition of Flavivirus infection), PP-110 (inhibitor of Src and Abl protein tyrosine kinases), AG14361 (antigout and antihyperuricemic), BRD-K35716340 (antibiotic), aminopurvalanol A (antiasthmatic), emetine (antiemetic and anthelminthic), BRD-K19724398, HG-5-113-01, and tremulacin (Fig. 8).
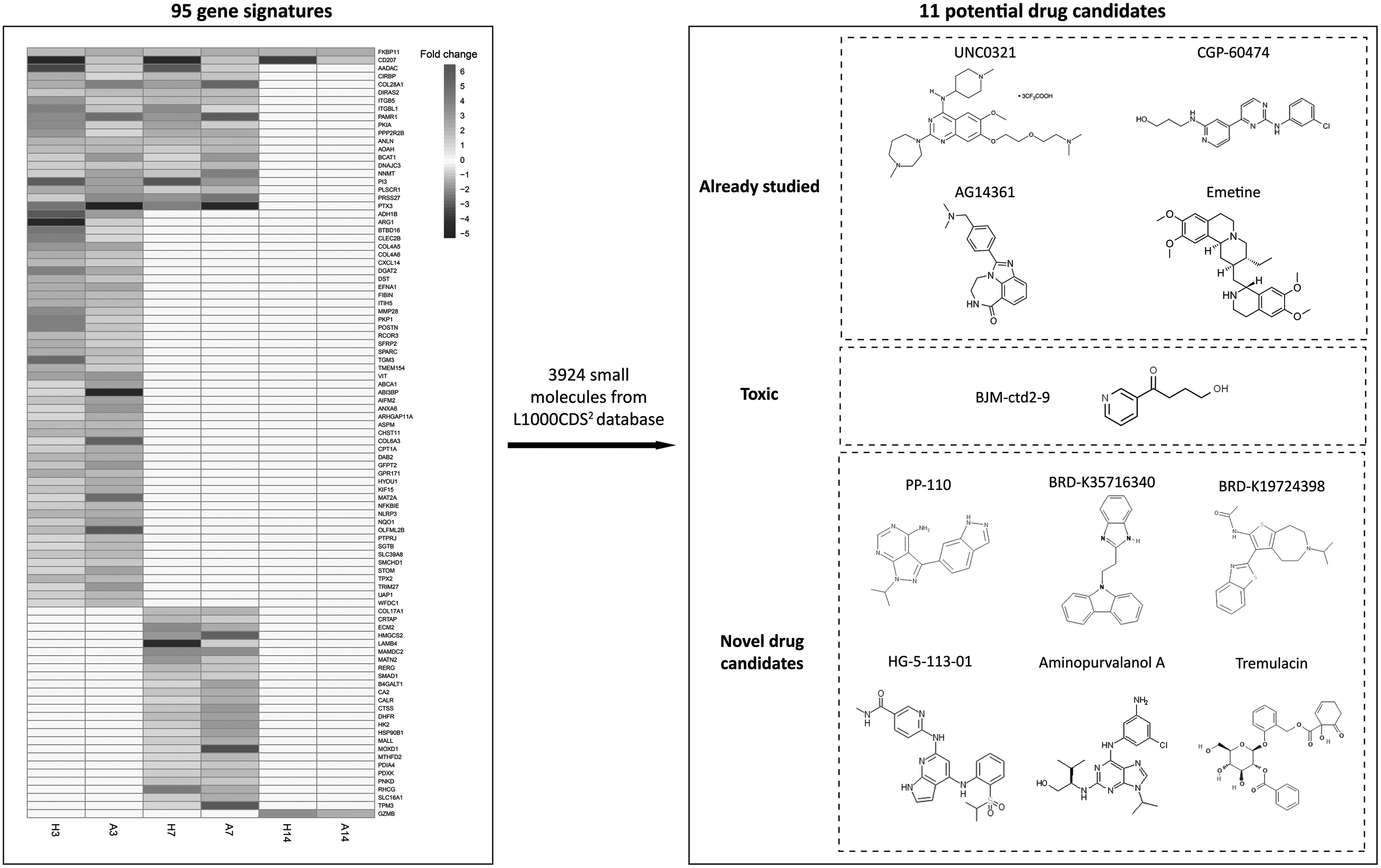
The graphical abstract of the results. Drug repositioning based on the expression profiles of 95 genes resulted in 11 potential small molecules, 6 of which were proposed as novel drug candidates for treatment of cancers.
Among them, UNC0321, CGP-60474, AG14361, and emetine are the potential cancer treatment molecules studied in literature. UNC0321 (epigenetic regulator of frataxin) is a protein lysine methyltransferase G9a inhibitor that is known to prevent cancer cell growth (Liu et al., 2010). The other molecule, CGP-60474, a cyclin-dependent kinase inhibitor, was classified as an effective drug for small cell carcinoma cells (Wildey et al., 2014). AG14361 (antigout and antihyperuricemic) is a PARP (poly(ADP-ribose) polymerase-1) inhibitor and is used for enhancing the effects of chemotherapy and radiotherapy in cancer treatment (Calabrese et al., 2004; Veuger et al., 2003). There exist various studies and applications of this molecule with other cancer drugs because it can avoid chemosensitization and radiosensitization (Curtin et al., 2004; Murai et al., 2012; Sajish and Schimmel, 2015; Smith et al., 2005). Specifically, this molecule was also found to be effective in BRCA2-deficient tumors (Vazquez-Ortiz et al., 2014).
Finally, emetine is known as an alkaloid produced by ipecac roots and used for antiparasitic and contraception purposes (Cuyas et al., 2015; Mehrotra et al., 2004). Emetine is also used as a small molecule inhibitor of protein synthesis and has toxicity in all types of cells; thus, this feature of emetine is crucial in heterogeneous cancer cells within a tumor (Akinboye et al., 2017). Specifically, emetine was recently proposed as a novel cancer drug for the treatment of breast cancer by decreasing estrogen receptor α level in breast cancer cells (Busonero et al., 2017).
BJM-ctd2-9, known as 4-hydroxy-1-(3-pyridyl)-1-butanone (HPB), is a N′-Nitrosonornicotine (NNN) derivative (Liu et al., 2016). NNN and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) were suggested to be the most carcinogenic tobacco-related molecules with forming DNA adducts or interacting with cytochrome p450 (Hang, 2010; Peterson, 2010). Because of its potential toxic effect, we concluded that this molecule could not be used as a potential cancer treatment drug. In addition, any studies related to the molecules PP-110, BRD-K35716340, BRD-K19724398, and HG-5-113-01 could not be found in literature. We believe that the effect of these molecules as anticancer molecules should be tested with in vitro and in vivo experiments.
Finally, two molecules, aminopurvalanol A and tremulacin are the drugs used for other purposes in literature. Aminopurvalanol A is a CDK1/cyclin B inhibitor that has been first proposed as an inhibitor in sea urchin while arresting embryos at the G2/M transition (Le Breton et al., 2003). Because aminopurvalanol A also selectively inhibits casein kinase I (CK1) from Toxoplasma gondii or any other parasites that was unexpected to happen, antiparasitic and antibiotic usage of this molecule have been proposed in several literature (Artz et al., 2011; Donald et al., 2005).
Besides, antiangiogenic effect of aminopurvalanol A has been proposed in one study (Zahler et al., 2010). Nonetheless, we could not find any specific studies relating aminopurvalanol A molecule with cancer or cancer treatment. Tremulacin is a glucoside that was isolated from Populus tremula by Thleme and Richter. It has also been demonstrated to cause inhibition of carrageenan-induced paw edema in rats and croton oil-induced ear edema in mice (Cheng et al., 1994). However, there are no known antitumor effects or any literature that relate tremulacin with cancer.
The drug repositioning procedure used in this study lacks experimental validation and therefore is classified as level 0 according to the drug repositioning evidence level classification scheme that ranges from level 0 referring to predictions that lack any in vitro or in vivo experimental validation, to level 4 assigned to approved drugs for a new indication (Oprea and Overington, 2015; Vogrinc and Kunej, 2017).
On the contrary, several success stories based on similar workflows are recently reported in prostate cancer (Turanli et al., 2019a) and breast cancer subtypes (Turanli et al., 2019b), where the effect of repurposed drugs were validated using an in vitro cell assay and evidences were provided for the high correlation of multiomics-based in silico predictions with in vitro assays (Turanli et al., 2018). As such, by means of further in vitro and in vivo experiments to demonstrate the antitumor effect of repositioned molecules, this study has the potential enhancing the impact of existing therapeutics by identifying novel drug molecules in the context of regeneration and cancer.
Conclusions
In this study, we compared axolotl wound healing to humans that would give possibility to figure out cancer-inhibiting or cancer-leading mechanisms and proteins that might decipher gene signatures bridging regeneration mechanisms with tumorigenesis. Six molecules came into prominence as novel potential drug candidates. Two of them, namely aminopurvalanol A and tremulacin are the drugs used for cancer nonrelated purposes in literature. The other four drugs, which are PP-110, BRD-K35716340, BRD-K19724398, and HG-5-113-01 could not be found in literature for any treatment purposes. The repositioned drugs conferred in this study deserve attention for further in vitro and in vivo studies because they point out potential candidate biomarkers therapeutics to treat cancer. We believe that elucidating the related mechanisms more deeply between regeneration and cancer would guide researchers in developing more therapeutic strategies from already approved noncancer drugs in the future.
Footnotes
Acknowledgments
The authors thank the 100/2000 YOK Doctoral Fellowship Program provided to Gizem Gulfidan.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.