
Abstract
Cancer as the leading cause of death worldwide has many issues that still need to be addressed. Since the alterations on the glycan compositions or/and structures (i.e., glycosylation, sialylation, and fucosylation) are common features of tumorigenesis, glycomics becomes an emerging field examining the structure and function of glycans. In the past, cancer studies heavily relied on genomics and transcriptomics with relatively little exploration of the glycan alterations and glycoprotein biomarkers among individuals and populations. Since glycosylation of proteins increases their structural complexity by several orders of magnitude, glycome studies resulted in highly dynamic biomarkers that can be evaluated for cancer diagnosis, prognosis, and therapy. Glycome not only integrates our genetic background with past and present environmental factors but also offers a promise of more efficient patient stratification compared with genetic variations. Therefore, studying glycans holds great potential for better diagnostic markers as well as developing more efficient treatment strategies in human cancers. While recent developments in glycomics and associated technologies now offer new possibilities to achieve a high-throughput profiling of glycan diversity, we aim to give an overview of the current status of glycan research and the potential applications of the glycans in the scope of the personalized medicine strategies for cancer.
Introduction
Cancer is the overriding cause of morbidity and mortality worldwide and represents a global health burden. In cancer, in addition to alterations in RNA, protein, and metabolite levels due to genome reprogramming, alterations in glycan compositions and/or structures such as glycosylation, fucosylation, and sialylation are also widely observed. In accordance with the development of glycan profiling approaches, implementation of glycan studies for cancer to determine potential cancer glycan biomarkers has increased day by day (Wang, 2013).
Glycosylation is the enzyme-catalyzed chemical reaction formed by the covalent attachment of a carbohydrate to a polypeptide, lipid, polynucleotide, or carbohydrate, or other organic compounds (Kunej, 2019). Glycosylation is a regular process and every enzyme is encoded by glycogens (the storage form of glucose in cells) (Wang and Jia, 1987). Glycosylation is a co-translational or posttranslational modification process and may result in different classes of glycans through their specific glycosylation process regarding the alterations on glycan molecules (Ohtsubo and Marth, 2006).
N-glycosylation is a process in which glycans attach to nitrogen of asparagine or arginine side chains. A 14-sugar precursor is first added to the asparagine in the target protein. The structure of this precursor is the same in most eukaryotes, containing three glucose, nine mannose, and two N-acetylglucosamine molecules. Through a complex sequence of reactions, this branched chain is attached to a carrier molecule called dolichol and then transferred to the appropriate location on the polypeptide as the target protein passes into the endoplasmic reticulum (ER) lumen to form N-linked glycans (Wang et al., 2013). O-linked glycans are attached to the hydroxyl oxygen of serine and threonine in general. O-glycosylation also occurs due to the binding of sugar residues to amino acids like tyrosine, hydroxylysine, or hydroxyproline side chains, as well as serine and threonine. It is one of the common modifications in mammalian cells.
In addition, C-glycosylation, glypiation, phosphoglycosylation result in other forms of glycans. C-linked glycans are rare glycans with a carbon added on a tryptophan side-chain as a result of C-glycosylation. Phosphoglycosylation process forms phosphoglycans that are linked through the phosphate of a phosphoserine. Finally, glypiation is one of the common posttranslational modifications of eukaryotic proteins involving the addition of glycosylphosphatidylinositol (GPI) anchor proteins to lipids through glycan linkages. GPI contains a conserved phosphoglycan that is modified in a cell- and tissue-specific manner (Thomas et al., 2020).
The glycosylation of glycoproteins can be significantly altered in cancer due to changes in the expression levels of glycosyltransferases as a result of oncogene-activated signaling pathways coupled with gain or loss in chromosome copy number (Abbott and Pierce, 2010). Collectively, these changes result in glycoproteins exported to the cell surface and extracellular region with altered glycan structures that can lead to significant changes in cell phenotype. Therefore, glycoproteins with altered glycan expression are highly likely biomarker candidates in cancer.
Fucosylation is the addition of fucose sugar to a molecule by fucosyltransferase enzymes (Costa et al., 2018; Shu et al., 2011), and sialylation is the covalent adjoining of sialic acid to the terminal end of glycoproteins by sialyltransferase enzymes (Mereiter et al., 2016; Vajaria et al., 2016). Elevated levels of fucosylation and sialylation have been studied in various pathological cases, including inflammation and cancer (Costa et al., 2018; Shu et al., 2011). Moreover, specific types of fucosylated glycoproteins, which distinguish fucosylated glycans such as sialyl Lewis a/x, have been suggested as cancer biomarkers (Saldova et al., 2008).
Tumors may have different underlying genetic causes and can be expressed differently in one patient versus another. This natural variability of cancer contributes to the growing sensitivity and field of personalized medicine. The password of personalized medicine stands for the right drug, for the right patient, at the right time, and at the right dose (Sadée and Dai, 2005). Since the glycans and their alterations can be observed in cancer cases, they are expected to be used in personalized medical approaches. Glycans are reported as dynamic biomarkers for predictive, preventive, and personalized medicine for several disorders, including cardiometabolic diseases (Wang et al., 2019), Type 2 diabetes (Adua, 2018), and chronic diseases (Adua et al., 2017; Wang et al., 2020).
Also, glycans are potential biomarkers of suboptimal health status studies recently covered for some of the population-based researches (Adua et al., 2019; Wang and Yan, 2012). Plus, the defects and/or alterations in protein N-glycosylation, protein O-glycosylation, combined glycosylation processes (multiple pathway involvement), and glycosphingolipid and GPI anchor synthesis could cause congenital disorders of glycosylation, which are clinically and genetically heterogeneous metabolic disorders (Grünewald et al., 2002; Wang et al., 2016a; Wang, 2019). Therefore, developments on glycan research might shed light on various diseases and are urgently needed. Researchers consider that to study just two biomolecules (nucleic acids and proteins) is mostly adequate in personalized medicine research and therefore tended to underappreciate carbohydrates (i.e., glycans), which are one of the building blocks of life (Özdemir et al., 2020).
However, it should be kept in mind that diverse glycans have long been known to play major metabolic, structural, and physical roles in biological systems. They appear to be ubiquitous to all cells in nature, and glycans are not different from other major macromolecular building blocks of life (nucleic acids, proteins, and lipids), simply more rapidly evolving and complex (Varki, 2017). Well, then why was the deserved attention not given to the glycans?
Although a variety of glycoprotein markers are routinely used in clinical laboratories, the general opinion is that most of the glycan markers do not still have enough sensitivity and specificity for early detection of cancers (except for well-known and widely used markers like CA125, PSA, and CA15–3), and are not recommended for clinical routine (Duffy, 1999; Meany et al., 2009; Wong et al., 2003). This is primarily because of the lower abundance of these glycoproteins. Since it is particularly challenging to detect lower abundant glycoproteins, it makes the situation more difficult and new avenues to overcome this pitfall are urgently needed. Moreover, the limited technology to detect and analyze glycans brings another bottleneck.
Besides their advantages, some tools can reduce the performance and limit analytical uses. Lectin-based assays, Western blotting (WB), mass spectrometry (MS), and high-performance liquid chromatography (HPLC) are the main technologies used in glycan analysis. Lectin-based assays have two main limitations that need to be overcome: (i) the weak affinity for their target glycan and (ii) the availability of lectins with uncommon/less-studied glycan structures (Haab, 2012). Difficulty in finding the primary antibody against the glycoprotein of interest is the biggest limitation for WB.
Moreover, many antibodies interact with other proteins and show off-target effects (Ghosh et al., 2014). For MS analysis, indispensable pretreatment operations (i.e., glycans that need to be separated and derived from proteins and lipids) prevent the direct application of glycan samples. Also, it is not possible to identify isomers and precise structural assignments based on the acquired mass spectra with MS. Similar to MS, in HPLC also, sample pretreatment is required and it is a relatively time-consuming method (Svarovsky and Joshi, 2014). Finally, the DNA and glycans are further complex and open ended than the sequence of a finite-length genome due to the reason that they do not synthesize by a template; that is why glycans are evaluated with different perspectives from other biomolecules (Özdemir, 2020).
Despite these limitations, it should not be forgotten that life needs more than nucleic acids and proteins, which can be completed by the integration of the glycans. In that regard, in this study, we aimed to depict hitherto reported glycan alterations in human cancers and present a catalog of clinically validated glycoproteins as cancer biomarkers. For this purpose, a detailed literature survey was performed for reported glycan alterations in human cancers and proposed glycoproteins as potential cancer biomarkers through electronic databases (i.e., Science Direct, Scopus, PubMed, and Web of Science).
The survey was based on several keywords such as “glycan biomarkers,” “O-glycans,” “N-glycans,” “glycosylation,” “sialylation,” and “fucosylation” and repeated independently by six researchers. In this review, the studies to be included were selected under the following criteria: (1) the articles should have been published between the years of 1985 and 2020, (2) the studies must be published as a review or an original research article, (3) the findings must be confirmed by wet-laboratory experimentation, (4) and the experiments should have been performed on human samples (tumor biopsy, body fluids, or cell lines). The literature survey resulted in 205 studies reporting 142 glycan alterations and 88 glycoprotein biomarkers associated with 26 human cancers.
Overview of Reported Glycan Alterations in Cancer
So far, a total of 142 glycan alterations (fucosylation, sialylation, and glycosylation) have been reported in human cancers (Tables 1–3). A summary of the reported alterations in human cancers and the experimental methodology employed during the studies is presented in Figure 1.
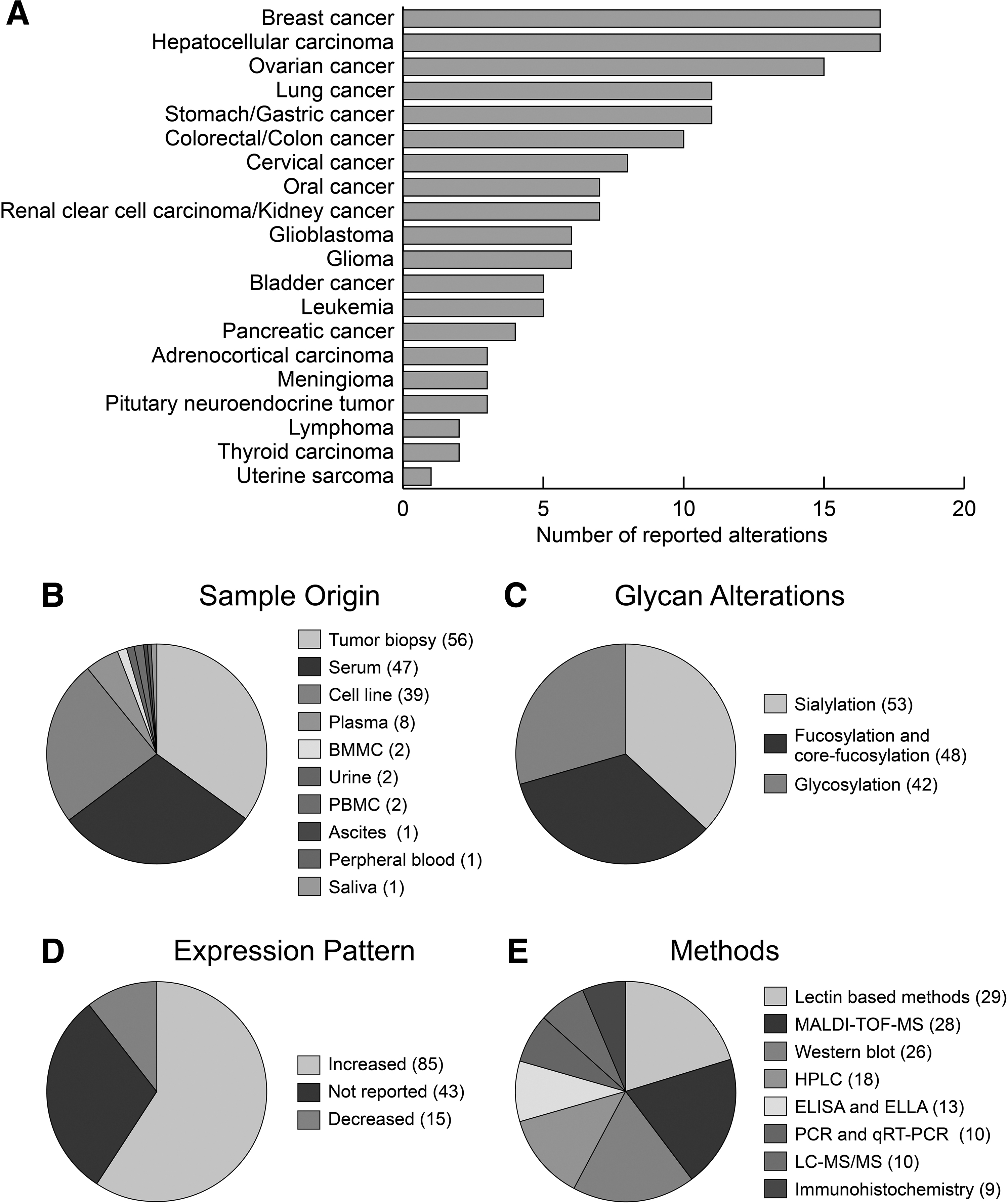
The conceptual summary of the conducted studies about glycan alterations for the 20 different types of cancers.
Altered Glycosylations in Various Tumors
IHC, immunohistochemistry; LA, lectin-based methods; FC, flow cytometry; HPLC, high-performance liquid chromatography; IB, immunoblotting; RT-PCR, reverse transcription polymerase chain reaction; IPREC, immunoprecipitation; WB, Western blot.
Altered Fucosylations (and Core Fucosylations) in Various Tumors
In terms of the reported number of glycan alterations, breast, hepatocellular, ovarian, stomach, and lung carcinomas were the leading types of cancer (Fig. 1A). In general, it has been observed that studies were focused on cancers more frequently observed in society. Either no study has been performed or very few studies have been reported on glycan alterations in tumors with low observation frequency or rare tumors.
In the detection of glycan alterations and preclinic and clinical validation studies, tumor biopsies, cell lines, and serum samples were preferred sample types in almost 90% of reports. A limited number of studies were carried out by the use of urine, saliva, ascites, bone marrow mononuclear cells, and blood constitutes such as plasma, peripheral blood mononuclear cells, or peripheral blood (Fig. 1B). Considering the frequent secretion of glycoproteins to the extracellular region, future studies should be extended to body fluids, which have been neglected so far.
The glycan alterations were primarily observed in terms of sialylation (37%). Further alterations in fucosylation and core fucosylation (34%) and glycosylation (29%) were observed (Fig. 1C). Although the expression patterns of the glycan alterations were varying, in general, the increased levels of fucosylation, sialylation, and glycosylation were encountered in cancers (Fig. 1D).
Numerous methods have been applied to the analysis of glycan profiles (Fig. 1E). However, a common strategy to identify glycan alterations was to use methods employing lectins, which are carbohydrate-binding proteins with variable affinities for specific glycan structures, and further separations using HPLC- and MS (MALDI-TOF-MS, and LC-MS/MS)-based technologies and validations using WB, polymerase chain reaction (PCR) and real-time quantitative reverse transcription PCR, enzyme-linked immunosorbent assay and enzyme-linked lectin assay, capillary electrophoresis, or immunohistochemistry.
When the altered glycosylations in human cancers were considered (Table 1), these alterations were primarily reported in hepatocellular carcinoma, which was commonly studied using cell lines, and the expression patterns (increased/decreased) were varying. No reported studies were found for leukemia, oral, and thyroid cancers. Out of 41 glycosylation studies, 12 of them used the LA technique. Moreover, MUC5AC, CD44, and CD133 were some of the affected proteins in terms of glycosylation alterations.
Breast and ovarian cancer were the two cancers in which sialylation alteration was mostly observed (Table 2). In both cancers, among reported expression patterns, all alterations were increased. No reported studies were found for adrenocortical, hepatocellular, pancreatic, and uterine carcinoma. Forty-seven percent of the sialylation studies were based on MS or MS-based (i.e., MALDI-MS, MALDI-TOF-MS, or nano-LC−MS) methods. ST3GAL, CA-125, and ST6GAL1 were some of the affected proteins in terms of sialylation alterations.
Altered Sialylations in Various Tumors
UHPLC, ultra-high-performance liquid chromatography; UPLC, ultra-performance liquid chromatography; BMMC, bone marrow mononuclear cell; PBMC, peripheral blood mononuclear cell.
Lung cancer was the most reported cancer type when the altered fucosylations (and core fucosylations) in various tumors were investigated (Table 3). Almost all of them had an increased expression pattern in lung cancer and were studied only in serum and tumor biopsy samples. No reported studies were found for adrenocortical, bladder, glioblastoma, leukemia, lymphoma, meningioma, pituitary neuroendocrine tumor, and uterine cancers. Of the 48 studies reported, 30 had an increased fucosylation (and core fucosylation) pattern, while only 6 had a decreased pattern. MALDI-TOF-MS was the quarter of the techniques applied in the fucosylation (and core fucosylation) studies. Some of the affected proteins associated with altered fucosylations (and core fucosylations) include haptoglobin and CD147.
Glycoproteins as Cancer Biomarkers
Glycoproteins are frequently secreted from cells or released from cell surfaces by hydrolytic cleavage, and therefore, glycoproteins with altered glycan expressions are likely candidates as biomarkers that can be involved in the prediction or monitoring the states of various diseases, including cancer. As a result of a systematic survey, we end up with 88 different glycoproteins as clinically validated glycoprotein biomarkers in various tumors (Table 4).
Glycoprotein Biomarkers Clinically Validated in Various Tumors
In terms of the reported number of glycoprotein biomarkers, hepatocellular carcinoma, lung cancer, and ovarian cancer were the leading types of cancer (Fig. 2A). Tumor biopsy, serum, and plasma (total 92%) were preferred primarily as sample origins in preclinic and clinical validation studies. A limited number of studies were carried out by the use of bone marrow, urine, blood, saliva, and ascitic fluid (Fig. 2B).
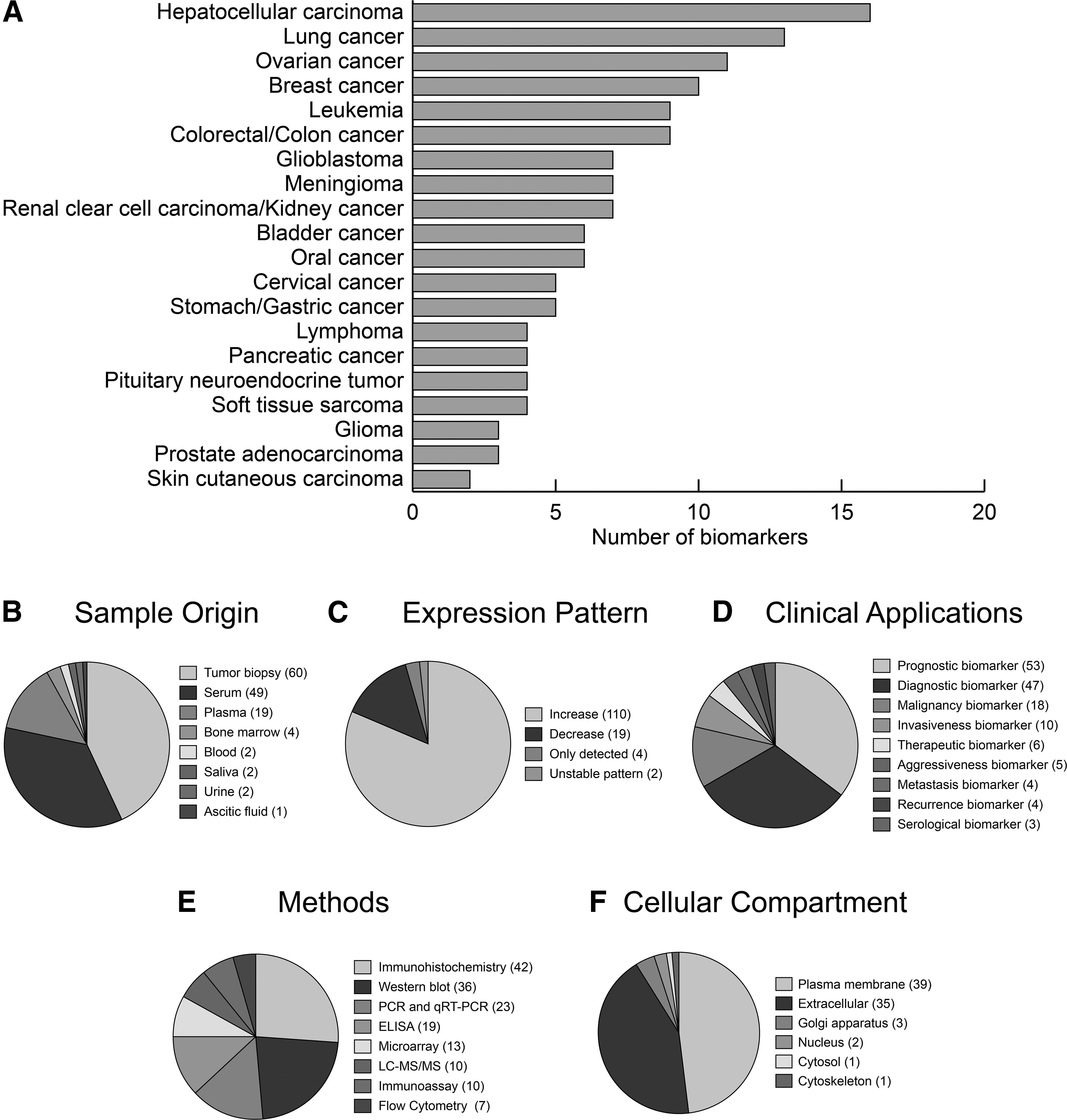
The conceptual summary of the conducted studies about glycoprotein biomarkers for the 20 different types of cancers.
When the expression patterns of glycoproteins were examined, it was observed that ∼82% of glycoproteins have increased expressions in cancer. If the expression pattern stated as only detected, the direction of expression was not examined in that study. Also, if the expression pattern is stated as an unstable pattern, the direction of expression was considered both increased and decreased (Fig. 2C).
The reported glycoprotein biomarkers have been proposed for numerous clinical applications such as prognosis (35%), diagnosis (31%), and malignancy (12%). Moreover, only 4% of the reported glycoprotein biomarkers were suggested as therapeutic targets for the treatment of cancers using various experimental methodologies (Fig. 2D, E).
Based on the cellular compartmentalization information provided by GeneCards database (Stelzer et al., 2016) and the Metascape toolbox (Zhou et al., 2019), the majority of the reported glycoprotein biomarkers (>90%) were either localized on the plasma membrane or secreted to the extracellular region (Fig. 2F). A limited number of listed glycoprotein biomarkers were involved in the Golgi apparatus, nucleus, cytosol, and cytoskeleton. This issue may be originated from the limitations of the detection techniques and analytical sensitivity. Since the detection of tiny amounts of glycoproteins is an uneasy procedure, the observations provided above might represent the localization of the glycoproteins that are available in high amounts in the plasma membrane and/or extracellular space.
Among 88 different glycoproteins, mucin-1 and syndecan-1 were mutually associated with six different cancer types. CD44 antigen and chondroitin sulfate proteoglycan 4 were common for five and four different cancer types, respectively. Six glycoprotein biomarkers (alpha-1-acid glycoprotein, decorin, endocan, haptoglobin, mucin-16, and zinc-alpha-2-glycoprotein) were common for three different cancer types. Also, 13% of the glycoprotein biomarkers were common for two different cancer types, whereas 75% of glycoprotein biomarkers were specific to a certain cancer type.
Considering that proteins do not function alone, and major biological processes are mediated through protein interactions, we aimed to map the physical interactions of glycoprotein biomarkers and reconstruct a protein-protein interaction (PPI) network around them. For this purpose, we employed gene symbols associated with the glycoprotein markers from UniProtKB (The UniProt Consortium, 2019) and GeneCards: The Human Gene Database (Stelzer et al., 2016), and the high confidence human protein interactome (with confidence score ≥0.8) (Karagoz et al., 2016).
The reconstructed PPI network was visualized by Cytoscape (v3.5.0) (Shannon et al., 2003) (Fig. 3). The reconstructed PPI subnetwork consisted of 961 nodes (i.e., 88 glycoprotein biomarkers and their physically interacting first neighbors) and 1743 edges (i.e., physical PPIs between these proteins). The proteins that play central roles in the modular organization and information flow within the network was identified by the topological analysis (using degree metric) of the reconstructed subnetworks. The top three central proteins were the epidermal growth factor receptor (EGFR, degree = 674), the proto-oncogene c-Fyn (FYN, degree = 470), and the cyclin H (CCNH, degree = 73).
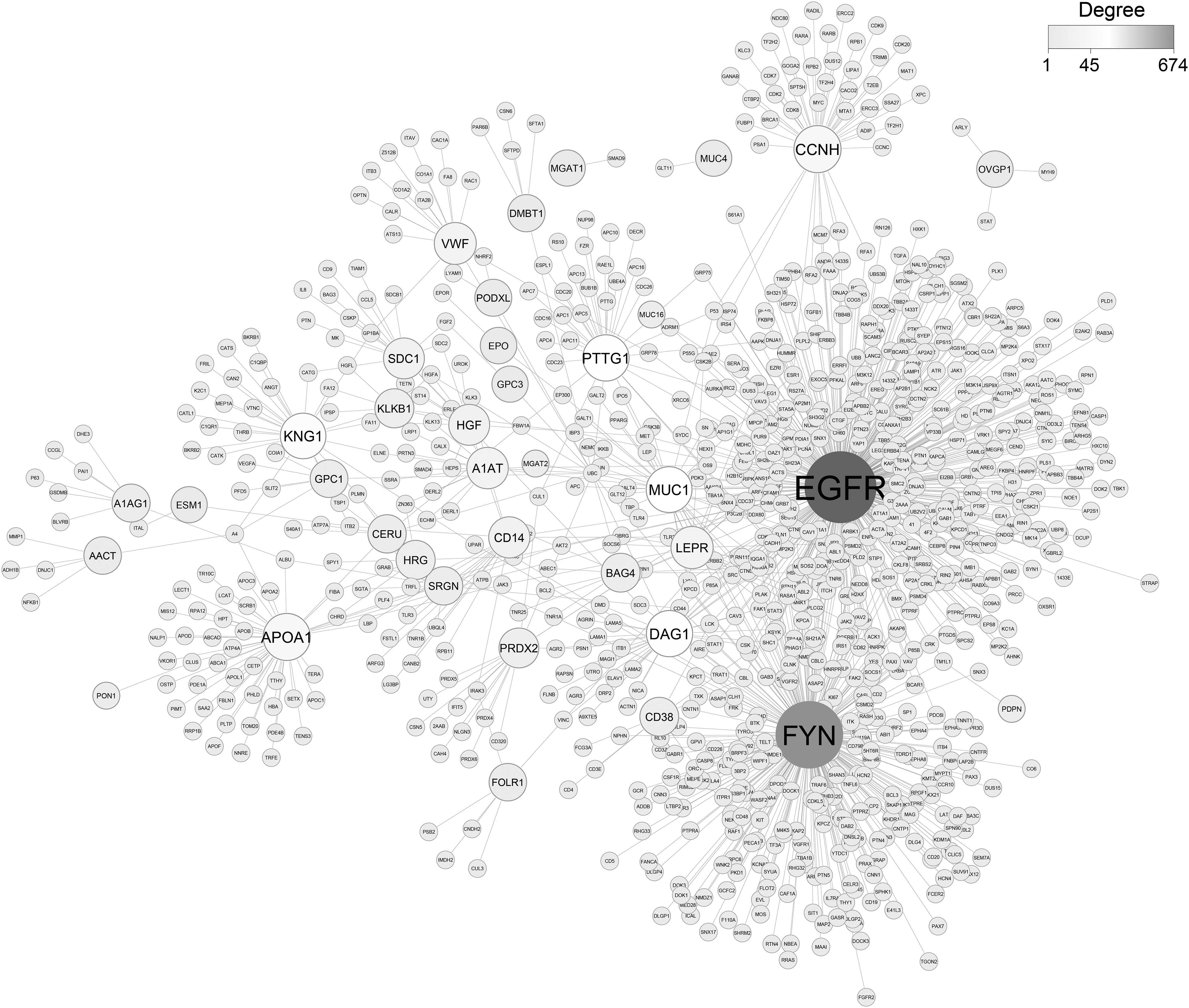
Network consisting of protein-protein interactions reconstructed around glycoprotein biomarkers. Proposed glycoprotein biomarkers are represented in larger nodes and dark gray color. As represented in the legend, nodes are sized and darkened in gradient based on the degree which glycoprotein biomarkers have.
Excess of EGFR is frequently related to high tumor grade, also causes poor treatment results, including chemotherapy and hormonal therapy resistance. EGFR overexpression has been determined as a powerful marker in head and neck, bladder, esophageal, ovarian, and cervical cancers, while it is a modest marker in gastric, endometrial, and colorectal cancers (Correia et al., 2014). Fyn has various functions associated with immune and neurological processes. Furthermore, Fyn attended in biological processes such as growth and proliferation, cell-cell adhesion, cell cycle entry, mitogenic signaling, and integrin-mediated interactions (Saito et al., 2010).
The upregulation expression pattern of Fyn was reported in several cancers, including melanoma, glioblastoma, breast cancer, prostate cancer, and squamous cell carcinoma (Elias and Ditzel, 2015). A recent study on CCNH suggested that its upregulation is associated with poor prognosis and promotes the growth of ovarian cancer (Peng et al., 2020)
To further elucidate the intertwined roles of these glycoproteins in signaling, metabolic, and disease pathways, we also performed pathway enrichment analysis through the Metascape (Zhou et al., 2019) using data from Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa et al., 2007), Gene Ontology (The Gene Ontology Consortium, 2019), and Reactome (Fabregat et al., 2018) databases.
The statistical significance of the enrichment results was represented by adjusted p-values obtained through Fisher's Exact Test and Benjamini–Hochberg correction procedure. Pathway enrichment analyses of the listed glycoproteins resulted in the following pathways (Supplementary Table S1): diseases of glycosylation, extracellular matrix (ECM) organization, matrisome-associated pathway, proteoglycans in cancer, and integrin 3 pathway, whereas gene ontology biological process enrichment analysis of glycoproteins resulted in the following terms: regulated exocytosis, platelet degranulation, regulation of cell adhesion, tissue remodeling, blood vessel development, and so on (Fig. 4A).
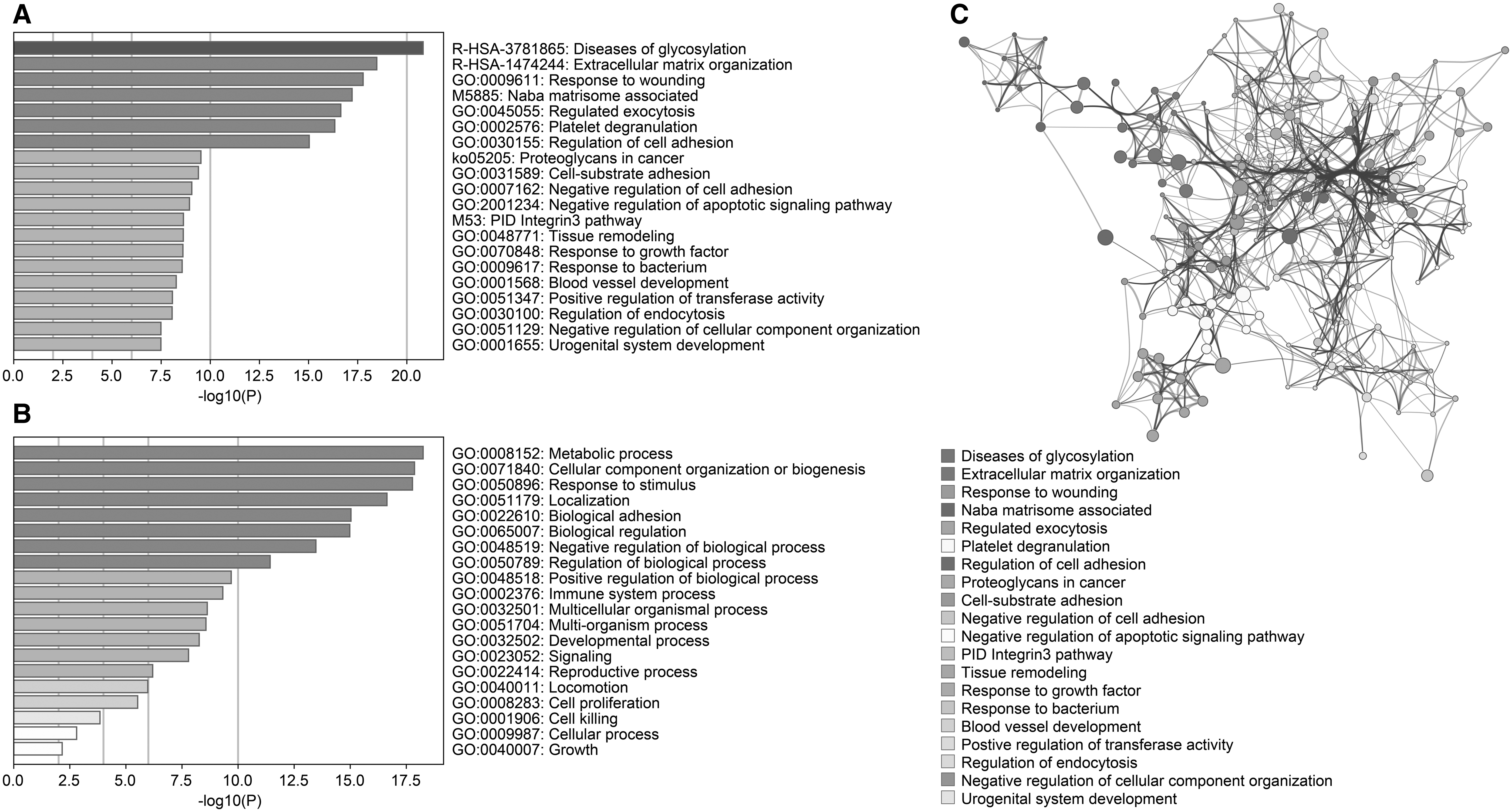
The enrichment results, including pathway and GO terms, of the genes encoding glycoprotein biomarkers.
The two following pathways, diseases of glycosylation and proteoglycans in cancer, clearly indicated that listed glycoproteins are literally addressing the human diseases, including cancer. The response to the wounding process was also associated with cancer recently and suggested that hallmarks of cancer are also the hallmarks of wound healing (MacCarthy-Morrogh and Martin, 2020). The study reports how tissue repair and cancer have common cellular and molecular patterns that are regulated in a wound, but misregulated in cancer. Listed glycoproteins were enriched with the response to wounding process. It wounding process thought to be like as MacCarthy-Morrogh and Martin scenario, tumors can be accepted as wounds that failed to heal (Dvorak, 2015).
The ECM organization and matrisome associated pathways were also enriched (Fig. 4A). When we consider the invasion and the metastasis of the tumors, ECM is the fundamental factor that influences tumor microenvironment and migration. The ECM provides not only signaling pathways through integrins and other membrane receptors but also supporting surrounding tissue and organs (Lu et al., 2012). Cancer cells can disrupt ECM fibers by matrix metalloproteinases that cause the migration of cells through collagens and basement membranes of the blood and lymph vessels that led to tumor invasion. The invasion triggers the cytokine stimulation in nearby tissues, which is an urgent call for the recruitment of tumor-associated macrophages to stop invasion through paracrine growth factor signaling loop (Conklin et al., 2011).
The enriched pathways and processes, including regulation of exocytosis, regulation of cell adhesion, ECM organization, tissue remodeling, response to growth factor, blood vessel development, and regulation of endocytosis, were all associated with the tumor microenvironment, migration, and invasion processes. Therefore, understanding the associations between the ECM and cancer cells may unveil possible therapeutic approaches in controlling cancer (Pal et al., 2020). The interactions of each pathway and process elements are represented in Figure 4B. The intertwined elements display how these pathways and processes are closely related to each other.
The biological processes of listed glycoproteins were identified as metabolic process, cellular component organization, response to stimulus, immune system process, developmental process, signaling, locomotion, cell proliferation, and cell killing (Fig. 4C). These processes are also associated with cancer hallmarks, primarily the tumor microenvironment, invasion, and migration.
Glycoproteins were also investigated for their tumorigenic potentials as tumor suppressors or oncogenes considering the information provided by Lawrence et al. (2014), which presents a comprehensive catalog, including a total of 1217 tumor suppressor and 803 oncogenes. Among the glycoprotein biomarkers, 6 of them were reported as tumor suppressors (CDH1, DCN, DMBT1, GPC3, HRG, and VIM) and 8 glycoproteins as oncogenes (AXL, CDH1, EGFR, FYN, MUC1, MUC4, PTTG1, and VIM).
Moreover, screening of the drug candidates considering these glycoproteins as potential drug targets through gene-drug interactions provided through Drugbank (Wishart et al., 2018), DGIdb (Edwards et al., 2011), GeneXpharma (Turanlı et al., 2017), and Comparative Toxicogenomics Database (CTD) (Davis et al., 2009) resulted in antineoplastic agents (docetaxel, fluorouracil, sorafenib, bevacizumab, capecitabine, carboplatin, cisplatin, dasatinib, and doxorubicin), corticosteroids (dexamethasone and methylprednisolone), immune system suppressors (methotrexate, tacrolimus, and prednisone), an alkaloid (colchicine), a cardioprotective agent (dexrazoxane), a monoclonal antibody drug (rituximab), and antitumor antibiotics (streptozocin and mitomycin) as potential repositioned drug candidates (Supplementary Table S2). These drugs mostly interacted with the plasma membrane and extracellular space glycoproteins.
Future Perspectives
Many studies discovered the glycan alterations and glycoprotein biomarkers in different cancer types as we reviewed. Except for cancer, glycans are also highly dynamic and alterations are observed in other common complex diseases as well. For instance, the aberrant immunoglobulin G (IgG) glycome composition or changes in IgG glycosylation were associated with the systemic lupus erythematosus (Vučković et al., 2015) and the remission status of rheumatoid arthritis (Sebastian et al., 2016).
Moreover, N-glycosylation of IgG was proposed as a potential biomarker of Parkinson's disease (Russell et al. 2017) and was associated with both chronological and biological aging (Yu et al., 2016), as well as the pathogenesis of hypertension (Wang et al., 2016b). Liu et al. (2018c) reported that loss of galactose and sialic acid impresses dyslipidemia development. The potential of variations in the composition of the N-glycome as biomarkers for several diseases such as metabolic syndrome (Lu et al., 2011), type 2 Diabetes Mellitus (Ge et al., 2018), and hematological cancers (Lauc et al., 2013) was already reported providing evidence for considering glycan alterations and glycoprotein biomarkers as potential candidates for the prognoses and treatment of virtually every disease.
The main bottleneck in glycan-based studies is using tumor biopsies as samples (Tables 1, 2). Since invasive operations are not patient friendly, there is an urgent need for noninvasive alternatives. Human serum, plasma, feces, hair, saliva, and urine hold the promise of being noninvasive samples for the identification of novel biomarkers, which can be detected in routine clinical diagnosis, prognosis, and targeted cancer therapies. Using urine, saliva, or blood-based test for early detection of cancer is a longstanding dream of cancer scientists. Such a discovery would make moving forward the war against cancer and would be more patient friendly.
The most common bottleneck in cancer research that scientists have been faced with is the elucidation of stable biomarkers, which can be routinely detectable in easily collectible samples mentioned above (Ren et al., 2020). There are just a few studies that consider noninvasive glycan biomarkers for distinguishing early stages of the diseases (Gressner et al., 2009; Qin et al., 2017; Yamasaki et al., 2015). Although a limited number of studies were conducted about noninvasive glycan biomarkers, it is not quixotic to dream about a future glycodiagnostic biomarker discovered from a single hair, urine sample, or saliva, since it is now possible to integrate the data from genomics, proteomics, metabolomics, and glycomics with pathway analysis.
Since all cells have a dense glycocalyx, the most secreted proteins are glycosylated (Svarovsky and Joshi, 2014). This common nature of glycan abundances highlights the crucial roles performed by them not only for normal morphological and physiological duties but also for abnormal disease-related pathological processes like cancer (Ohtsubo and Marth, 2006). Glycosylation is constituted on surfaces of the cell, which is the primary point of contact in cellular interactions through extracellular matrices (Adamczyk et al., 2012) when any disturbances occur. Hence, the causes of disease states on glycan biosynthesis may be more conspicuous than disease-associated aberrations in proteins. Thus, gaining knowledge about glycans and glycosylation patterns has become a promising window that allows researchers to identify potential biomarker candidates for different cancer types.
It is well established that in tumor cells, glycans on cell surface glycoproteins are essentially altered, and as a result, those cells have a diverse ‘glycan coat’ when compared to healthy cells. Therefore, pioneers of the glycobiology field should envisage the construction of a broad atlas of cell-specific glycomes for the whole body of the human as providing a database or the amalgamation of protein glycosylation features in gene and protein databases. Access to databases for glycomics and glycoproteomics is essential for the glycan-based code of human disease research (Schjoldager et al., 2020). Genomic and proteomic databases can be searched for the expression of glycosylation-related genes to unveil glycosylation signatures related to cancer development, progression, invasion, and metastasis (Rodriguez et al., 2018).
Although aberrant glycosylation has arisen as a hallmark of cellular oncogenesis, the understanding of a tumor-associated glycome is fundamentally associated with the elucidation of the metastasis-associated processes (Häuselmann and Borsig, 2014). The translational and clinical applications of altered glycan structures in cancer have not yet been fully achieved (Cagnoni et al., 2016). However, in recent years, lectins were studied for deciphering the glycome and cracking the glyco-code for cancer therapy as a logical target.
There are effortful studies that shed light on the glycol-code of cancers with the focus of developing therapeutic strategies using lectin–glycan interactions (Dalziel et al., 2014). If the increasing amount of data on glycomics and glycoproteomics can be combined with the advances of other omics technologies, it will have a major impact on revealing therapeutic targets and patient stratification. Also, glycoengineering of the glycan structures is another important issue that should be mentioned. The re-designing of the known glycans through glycoengineering strategies can serve as the production of therapeutic proteins, including antibody-based therapeutics and immunotherapeutics (Buettner et al., 2018; Dicker and Strasser, 2015; Jefferis, 2009).
As cancer researchers make great strides for detection, prognosis, and therapeutic targeting, the amount of data will be increased exponentially, which allows us to see advanced analysis using artificial intelligence (AI) and machine learning (ML). Researchers have started to use AI algorithms to gather not only information on glycans but also glycan-based modifications (glycosylation, fucosylation, sialylation, etc.) to elucidate a comprehensive prediction of the detection of cancer.
Currently, glycan marker discovery is carried out mostly by analyzing high-throughput data producer platforms such as MS types and lectin microarrays (Lai et al., 2018; Manimala et al., 2006). However, these approaches are high dimensional and have chaotic patterns with a high level of noise sourced from measurements, disease heterogeneity, and biological variability (Tang et al., 2010). Attuned computational methods are needed to overcome a problem from such high-throughput data producer platforms in the way of discovery of biomarkers.
ML methods such as support vector machines (SVM) and deep neural networks have been employed for the marker selection in the above-mentioned methods (Hwang et al., 2020; Lai et al., 2018; Tang et al., 2010). This review might provide the learning data for such methods. The success of future glycodiagnostics will eventually base upon the utility of new and improved artificial intelligence methods. The improvement of these techniques might reveal better identification of crucial cancer biomarkers and accelerate the unveiling of glycodiagnostics, glycoprognostics, and glycotherapeutics.
Concluding Remarks
As a final remark, it seems that glycan alterations and glycobiomarkers will be outstanding for efficient biologics to personalized medicine treatments of cancer in the context of heterogeneity and specificity/sensitivity in the near future. We have summarized the major impacts of glycosylation, fucosylation, and sialylation alterations on understanding cancers. Glycoproteins have been valuable vehicles not only for the elucidation of pathomechanisms of the cancers but also for developing therapeutic strategies for cancers.
The system biomedicine approach believes that there is a holistic harmony of many crucial life events at different levels (i.e., nucleic acids, proteins, or metabolites) and argues with the traditional approaches that believe in single gene/single protein/single molecule. With this context, glycan biomarkers can be potential candidates, but a more robust cluster that is constituted from different omics level biomarkers is needed to eliminate patient variability, biomarker characteristics from many ethnicities, and differences coming from daily life routines.
From a translational medicine point of view, improvements in the new gene-editing methods are now providing more extensive analyses and genome-wide scans of functions of glycans to elucidate possible glycan interactions and functions and engineer design of glycoproteins that allow discovering and evaluating advanced drug and vaccine designs. If glycomics information can be combined with the other omics levels (i.e., transcriptomics, proteomics, and metabolomics), the personalized medicine strategies that will be applied to the patients will be splendid. The integration of the knowledge that comes from different omics levels will be amalgamated to yield clinically practicable glycan-based cancer diagnostics and therapeutics. Improvements in this intriguing field are eagerly awaited.
Footnotes
Acknowledgments
The scholarships under the TUBITAK 2211-C Doctoral Fellowship Program provided to Medi Kori and Busra Aydin; the scholarships under the TUBITAK 2211-A Doctoral Fellowship Program provided to Gizem Gülfidan Yıldız; and the scholarships under the YOK 100/2000 Doctoral Fellowship Program provided to Medi Kori, Busra Aydin, and Gizem Gülfidan Yıldız are greatly acknowledged.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.