
Abstract
Hepatitis B virus (HBV) infection has been causally linked to hepatocellular carcinoma (HCC) in more than 50% cases. MicroRNAs (miRNAs) play cross-cutting mechanistic roles in the complex interplay between viral pathogenesis, host survival, and clinical outcomes. The present study set out to identify etiologically significant human miRNAs associated with HBV infection in liver-related pathologies leading to HCC. In diverse tissue types, we assembled 573 miRNAs differentially expressed in HBV-associated liver pathologies, HBV infection, fibrosis, cirrhosis, acute on chronic liver failure, and HCC. Importantly, 43 human differentially expressed miRNAs (hDEmiRs) were regulated in serum/plasma and liver tissue of patients with HBV-positive conditions. However, only two hDEmiRs, hsa-miR-21-5p and hsa-miR-143-3p, were regulated across all disease conditions. To shortlist the functional miRNAs in HBV-induced HCC pathogenesis, a reverse bioinformatics analysis was performed using eight GEO datasets and the TCGA database containing the list of differentially regulated mRNAs in HCC. A comparative study using these data with the identified targets of hDEmiRs, a set of unidirectionally regulated hDEmiRs with the potential to modulate mRNAs in HCC, were found. Moreover, our study identified five miRNAs; hsa-miR-98-5p, hsa-miR-193b-3p, hsa-miR-142-5p, hsa-miR-522-5p, and hsa-miR-370-3p targeting PIGC, KNTC1, CSTF2, SLC41A2, and RAB17, respectively, in HCC. These hDEmiRs and their targets could be pivotal in HBV infection and subsequent liver pathologies modulating HCC clinical progression. HBV infection is the largest contributor to HCC, and the present study comprises the first of its kind compendium of hDEmiRs related to HBV-related pathologies.
Introduction
Hepatocellular carcinoma (HCC) constitutes more than 90% of all primary liver malignancies and is reported in approximately 85% of individuals diagnosed with cirrhosis (Asafo-Agyei and Samant, 2024; Llovet et al., 2021). HCC currently ranks as the fifth most common cancer worldwide (Fahmy et al., 2024; Xu et al., 2018). Chronic viral infections such as hepatitis B virus (HBV) and hepatitis C virus (HCV) are chief etiologies in HCC development, wherein alcohol consumption, aflatoxin-like carcinogens, and genetic and environmental factors are also crucial. Notably, HBV infection contributes to more than 50% of HCC cases (Jiang et al., 2021). HBV is a hepatotropic stealth virus that enters host cells through the sodium taurocholate cotransporting polypeptide receptors and replicates in normal immune anergy (Tan et al., 2015; Zhang et al., 2016b). It transmits through unsafe injection practices, blood transfusions, sexual contact, and vertically from mother to infant (Sabeena and Ravishankar, 2022). HBV’s partially double-stranded relaxed circular DNA genome gets converted into covalently closed circular DNA using the host cell’s DNA repair mechanisms, forming viral mini-chromosomes that replicate along with the hepatocytes, making it difficult to eradicate from the host cell completely. Moreover, the integration of HBV DNA into the host genome, which is not usually seen in HBV-infected conditions, has often been observed in HCC patients, altering host gene expression (Yang et al., 2018).
Furthermore, HBV infection creates an inflammatory immune response within the liver, leading to a continuous flux of immune cells, such as T cells and macrophages, into the liver tissue (Chen and Tian, 2019). Persistent HBV infection, coupled with ongoing immune response, contributes to liver damage, overtime leading to cell death and fibrosis, the initial stage of liver scarring (Luedde et al., 2014). Prolonged fibrosis can progress to cirrhosis, characterized as permanent liver scarring that can potentially lead to liver failure (Lv et al., 2023). Liver fibrosis and cirrhosis are classified into five stages (S0–S4), indicating the disease progression and risk elements (Berzigotti, 2017; Rossi et al., 2007). The complex interplay between the HBV virus and the host immune response inducing these liver pathologies can eventually culminate into HCC over a few years to several decades (Liang, 2009).
MicroRNAs (miRNAs) orchestrate gene silencing by forming complementary bonds with the 3′ untranslated region of target messenger RNAs (mRNAs), initiating their posttranscriptional gene regulation. Each miRNA possesses the potential to target multiple mRNAs, and conversely, individual mRNAs can be regulated by multiple miRNAs (Li et al., 2008). The maladjustment of miRNA expression levels has a profound effect on the inception and progression of diverse neoplastic conditions and, by extension, is poised to serve as potential diagnostic and prognostic biomarkers. In addition to cellular miRNAs, secretory miRNAs are transported exosomally from their parent cell into the body fluids. These exosomal miRNAs also play a pivotal role in cancer progression and can also have utility as biomarkers (Li et al., 2022).
Various cell and molecular factors such as miRNAs regulated by both the host and HBV play a significant role in HCC progression (Arbuthnot and Kew, 2001; Ligat et al., 2021). For instance, HBV infection potentially upregulates miR-802, which, in turn, targets SMARCE1C, consequently suppressing its counteractive role in generating HBsAg and HBeAg. Further, overexpression of miR-501 aids HBV survival by targeting HBXIP, an HBV replication inhibitor protein (Jin et al., 2013; Wang et al., 2019). Furthermore, the miRNAs such as miR-620, miR-1270, and miR-1236-3p are downregulated in HBV infection, leading to the upregulation of alpha-fetoprotein, an HCC biomarker, aiding its tumor-promoting intracellular functions (Zhang et al., 2016a). In addition, miRNAs such as miR-122 involved in lipid metabolism, miR-375 associated with glucose metabolism, and miR-132 implicated in inflammatory processes are frequently differentially regulated in HCC (Li et al., 2019; Morishita et al., 2021). There is a dynamic interplay between miRNAs and viral infections, where both the host cellular machinery and viral proteins contribute to changes in miRNA expression patterns, resembling a tug-of-war scenario (Skalsky and Cullen, 2010).
Our study aims to identify etiologically significant human miRNAs associated with HBV infection and related pathologies leading to HCC. We report new findings herein that comprise the first of its kind compendium of human differentially expressed miRNAs (hDEmiRs) in HBV-related pathologies—acute and chronic HBV infection, fibrosis, cirrhosis, acute on chronic liver failure (ACLF), and HCC. Furthermore, our bioinformatics analysis using the data on differentially regulated mRNAs from GEO datasets and TCGA datasets identified the functionally significant hDEmiRs in HBV pathology, which could be significant in developing prognostic and treatment strategies for HBV-induced HCC.
Materials and Methods
Data collection and curation
A PubMed search was carried out using the search term “HBV and microRNA” fetching 760 articles, which were screened for data on hDEmiRs in patients with HBV infection and HBV-induced liver pathologies using our unique curation criteria (Antil et al., 2022; Ramakrishnan et al., 2023; Ramesh et al., 2023). The disease conditions considered were acute/chronic/asymptomatic HBV infections, different levels of fibrosis and cirrhosis, ACLF, and HCC. The inclusion criteria had HBV infection as an essential factor in all pathologies, and data on liver diseases induced by other etiologies such as HCV infection were omitted.
Eighty-five articles were selected from the screened list as relevant to the study, and the hDEmiRs in each disease condition/context compared with the healthy control group along with the tissue type in which the study was conducted, the target of miRNAs, and other data connected to this were manually curated and segregated. The premature forms of miRNAs were omitted, and only the mature human miRNAs whose nomenclature starting as “hsa-miR” or “hsa-let”, wherein “hsa” stands for Homo sapiens, were gathered (Griffiths-Jones et al., 2006; Vishwakarma et al., 2024). The target mRNAs of these miRNAs were fetched from DIANA-TarBase and miRTarBase and used for further analyses (Hsu et al., 2011; Karagkouni et al., 2018).
A comparative analysis of hDEmiR targets with GEO Datasets and TCGA
The data on the differentially regulated mRNAs in HCC patients’ liver tissue compared with adjacent nontumor tissue were gathered from the GEO datasets. The Cancer Genome Atlas (TCGA) database contains the list of up- and downregulated mRNAs in dead and live liver cancer patients (Cancer Genome Atlas Research Network et al., 2013). This data were filtered to find those mRNAs exclusively up- or downregulated in dead and live HCC patients with a cutoff value of unidirectionally regulated in at least 10 patients.
The mRNAs differentially regulated in HCC patients from GEO datasets and TCGA were compared with the fetched targets of curated liver tissue hDEmiRs to study the effect of miRNAs on their targets in HCC condition.
HBV infection-specific hDEmiRs in HBV-infected HCC tissues
To analyze the subset of HBV infection-specific miRNAs in the curated hDEmiRs from liver tissue of HCC patients, the differentially expressed miRNAs upon HBV infection in cell lines were used from our previously published data (Ramakrishnan et al., 2023).
Results
A repertoire of hDEmiRs in HBV-induced liver pathologies
We carried out an extensive manual curation using our unique criteria, and 573 hDEmiRs in 35 different HBV-related liver disease conditions and 8 different tissue types, namely, serum, plasma, liver tissue, peripheral blood mononuclear cells (PBMCs), monocytes, CD4+ and CD8+ cells, and urine, were assembled from 85 articles. Due to lack of data, only the differential expression of miRNAs in serum, plasma, and liver tissue were analyzed in this study (Fig. 1). Categorization of disease conditions into five different contexts of HBV-induced liver pathologies—HBV infection, fibrosis, cirrhosis, ACLF, and HCC—was done for ease of data handling (Supplementary Table S1). A heat map was built with data on hDEmiRs validated using qRT-PCR and TaqMan PCR in different HBV-induced disease contexts (Fig. 2).
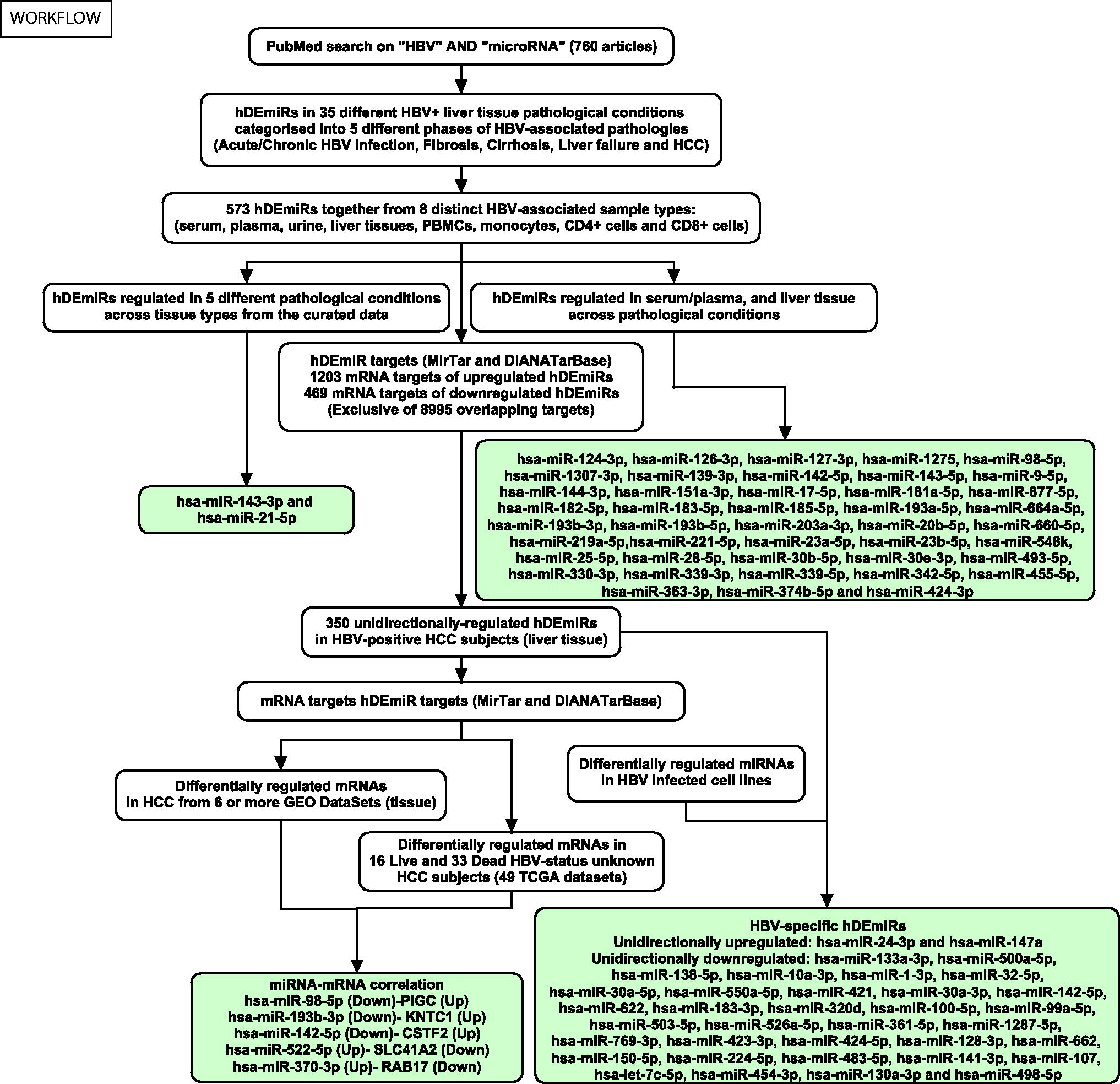
Workflow—the methods and datasets used for the study along with the results are depicted in the figure.
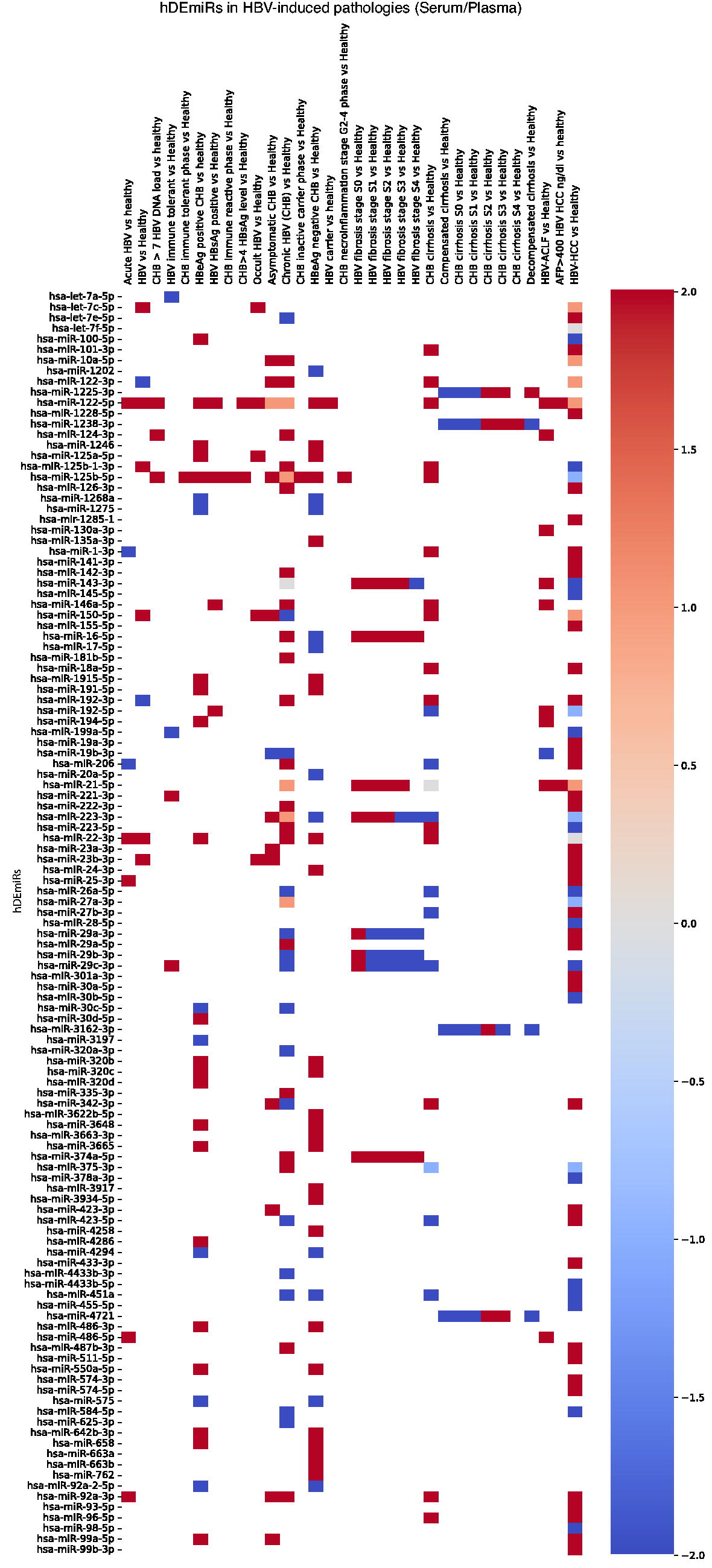
Heatmap of hDEmiRs—the hDEmiRs in all the HBV-induced pathologies validated using qRT-PCR and TaqMan PCR are illustrated as a heatmap showing their expression status in
(Continued).
Among the hDEmiRs, 43 were found to be differentially expressed across plasma/serum and liver tissue (Fig. 1, Supplementary Fig. S1A). However, hsa-miR-143-3p and hsa-miR-21-5p were found to be the only two hDEmiRs regulated across the five different contexts of “HBV vs healthy,” “fibrosis vs healthy,” “cirrhosis vs healthy,” “ACLF vs healthy,” and “HCC vs healthy,” irrespective of the tissue types. Their expression patterns in each dataset are gathered in Figure 1 and Supplementary Figure S1B. Thus, it could be inferred that these miRNAs might have significant functions in all the HBV-induced pathologies. Moreover, 30 hDEmiRs in the curation data were reported in extracellular vesicles cargos in HBV infection and related pathologies.
Comparative analysis of hDEmiR targets of HBV-induced HCC patients with GEO Datasets
miRNAs are used in personalized/precision medicine approaches as biomarkers and therapeutic targets so as to inform the development of future individually tailored treatments (Szczepanek et al., 2022). However, not all the target mRNAs are modulated by miRNA dysregulation in a disease condition. To study the impact of the hDEmiRs on their target genes, we have done a comparative study of the targets of hDEmiRs in liver tissue of HBV-induced HCC patients with the data on mRNA expression in HCC patients irrespective of their etiologies from GEO dataset.
Through this analysis, we observed that 4 mRNAs, targeted by 7 upregulated hDEmiRs, were indeed downregulated, whereas 30 mRNAs, targeted by 5 downregulated hDEmiRs, upregulated in GEO dataset (Fig. 3, Supplementary Table S2, Supplementary Fig. S2A). For example, the mRNA Midline 1 interacting protein 1 (MID1IP1) was upregulated in seven GEO datasets and seen as the target of exclusively downregulated miRNAs: hsa-miR-17-5p and hsa-miR-196b-5p. This finding aligned with the current knowledge on MID1IP1, which aids metastasis by promoting FRA1-mediated-MMP9 signaling. It is also involved in hepatic lipogenesis by activating acetyl-coenzyme A carboxylase. The miRNA hsa-miR-17-5p targeting MID1IP1 was downregulated in different HBV-related pathologies, including HCC, and hsa-miR-196b-5p was downregulated in HCC. Moreover, miR-17-5p upregulation inhibited postoperative HCC metastasis (Chiu et al., 2023; Liu et al., 2020).
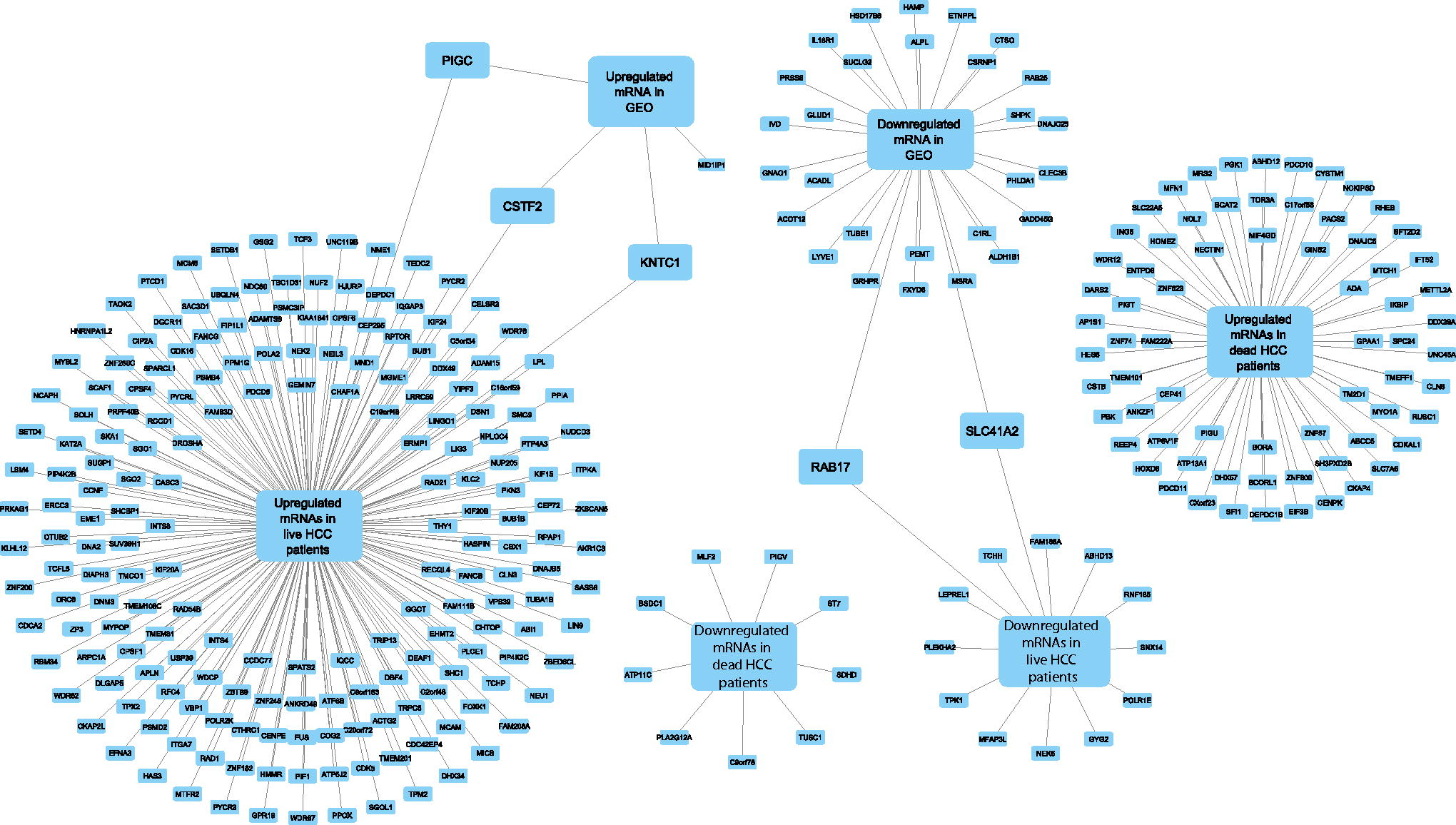
GEO dataset and TCGA data analysis—the hDEmiR target mRNAs that are regulated in accordance with the expression status of these miRNAs in GEO datasets and TCGA data and their overlap are shown in the figure. mRNA, messenger RNA; miRNA, microRNA.
Comparative analysis of hDEmiR targets of HBV-induced HCC patients with TCGA
In addition, to study the effect of hDEmiRs on HCC prognosis, we compared the TCGA data with the targets of hDEmiRs in HBV-induced HCC tissue (Supplementary Fig. S2B; TCGA overlap). Although there were overlaps between mRNAs with opposite expression patterns between TCGA and the hDEmiR targets, we have used only those target mRNAs that showed a unidirectional expression pattern. Such 213 mRNAs (199 upregulated and 14 downregulated) were found to overlap between the hDEmiR targets and differentially expressed mRNAs in live patients. Similarly, 79 mRNAs (70 upregulated and 9 downregulated) overlapped between the hDEmiR target and differentially regulated mRNA in dead patients (Fig. 3).
The miRNAs that regulate these mRNAs were mapped from our curated data, and it was found that 89 hDEmiRs targeted the differentially regulated mRNAs in live patients. Among these, 83 were found to be targeting the upregulated and four were targeting the downregulated mRNAs, respectively, in live patients. Meanwhile, 55 miRNAs were found to regulate the mRNAs differentially expressed in dead patients (Supplementary Table S3). Similarly, there were 51 in the upregulated target regulating miRNA list and 4 in the downregulated list in dead patients’ data. Among these hDEmiRs targeting mRNAs differentially regulated in HCC dead/live patients, 17 belonged to the list of 43 common differentially expressed hDEmiRs in liver tissue, serum/plasma samples of patients suffering from HBV-induced pathologies.
From this analysis, we found that many, but not all, differentially expressed mRNAs known to have roles in HCC progression could be regulated by hDEmiRs. For instance, ERCC3, which was upregulated in HCC patients (TCGA data) and in the hDEmiR target list, was targeted by the exclusively downregulated miRNA hsa-miR-192-5p. ERCC3 upregulation is correlated with the inhibition of MAP9 tumor suppressive effect and increased tumor progression (Zhang et al., 2020).
HBV infection-specific hDEmiRs in HBV-positive HCC liver tissue
Differentially regulated miRNAs in HBV-infected cell lines were taken from our previously published data. Specifically, 362 miRNAs were studied in the context of hDEmiRs, and it was found that 113 miRNAs were downregulated in both, of which 34 showed unidirectional expression. Two miRNAs were found in the unidirectionally upregulated list among the 36 upregulated miRNAs in both HBV-infected cells and the hDEmiR list (Fig. 4, Supplementary Table S4).
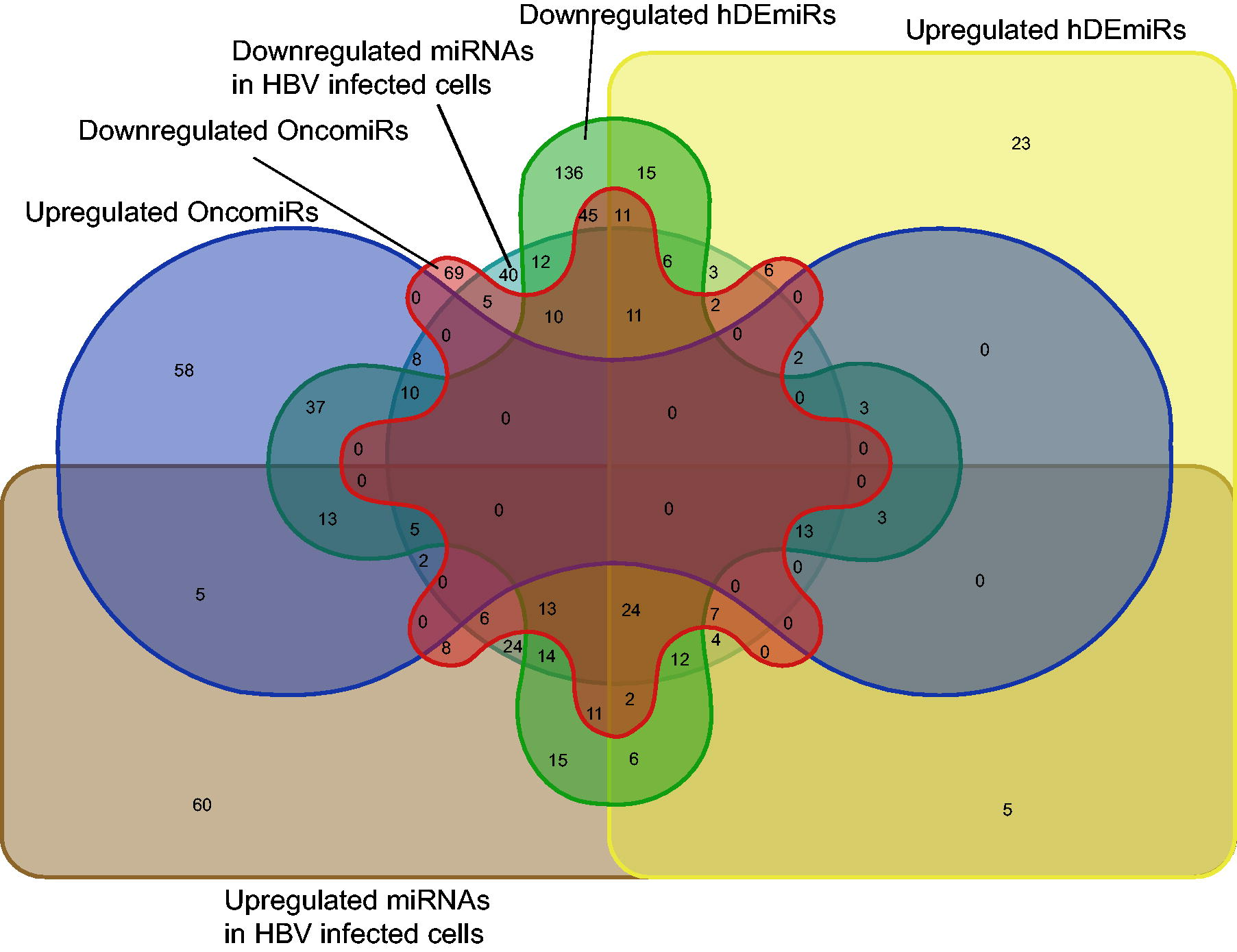
HBV infection-specific hDEmiRs in HBV-positive HCC tissues—the comparison between the hDEmiRs and the differentially regulated miRNAs in HBV-infected cells. HCC, hepatocellular carcinoma.
Discussion
Despite the diversity of HCC etiologies, HBV is the primary causative factor contributing to more than 50% of cases (Michielsen and Ho, 2011). However, HBV infection is infamous for its insidious nature, resulting in late diagnosis. Thus, precision diagnosis and therapeutic strategies against HBV infection and associated pathologies progressing into HCC are much needed, and miRNAs offer promise toward this goal (Ardekani and Naeini, 2010; Wang and Wei, 2020).
To unpack the functional role of miRNAs in HBV-induced HCC development, a comprehensive list of 573 hDEmiRs in HBV-induced pathologies in different tissue types, ranging from HBV infection, fibrosis, cirrhosis, ACLF, and HCC, was analyzed in the present study. Two hDEmiRs, namely, hsa-miR-21-5p and hsa-miR-143-3p, were found to be modulated in all the five pathological contexts and 43 hDEmiRs across serum/plasma and liver tissue. Our analysis of differentially regulated miRNAs in HBV-infected cell lines identified 2 upregulated and 34 downregulated miRNAs as HBV infection-specific hDEmiRs in HCC tissues implying their significant contribution to these pathologies. Their expression status and, at times, function vary with the disease conditions, even in the same etiology. For instance, the hsa-miR-21-5p was found to be both up- and downregulated in HBV infection and HCC. The miRNA expression patterns change temporally but do not always correlate with their target mRNA expression levels (Bozkurt et al., 2021; Lindholm et al., 2019). Therefore, the expression of miRNAs is dynamic in each of the pathology and warrants further study.
Using a reverse bioinformatics strategy, the publicly available datasets, GEO and TCGA datasets, were analyzed along with the targets of liver tissue-specific hDEmiRs in HBV-induced HCC patients. Through the comparative study with GEO datasets, unidirectionally modulated genes such as MID1IP1, which was upregulated in HCC datasets, were found to be targeted by hsa-miR-17-5p and hsa-miR-196b-5p, that are exclusively downregulated hDEmiRs in HBV-induced HCC tissue. The hsa-miR-17-5p also showed similar expression in HBV-infected patients’ serum. MID1IP1 is known to be upregulated in HCC and many other cancer types inducing metastasis whose depletion attenuates c-MYC, a protumorigenic gene, and activates p21, an inducer of cell-cycle arrest (Chiu et al., 2023; Huang et al., 2022; Jung et al., 2020; Lin et al., 2010). Moreover, hsa-miR-17-5p overexpression was found to inhibit HCC metastasis, signifying the role of these miRNAs in HBV-induced HCC (Liu et al., 2020). In contrast, the target mRNAs of hsa-miR-196b-5p were mostly downregulated in dead patients opposing its unidirectional effect in live tissue, demonstrating the complexity of miRNA regulation in these pathologies demanding further validation.
An analysis of TCGA datasets found a significant overlap between the differentially expressed mRNAs in patients with the targets of hDEmiRs. These target mRNAs and their corresponding miRNAs might have vital functions in HBV-induced HCC disease progression. Unidirectional hDEmiRs such as the downregulated miRNA hsa-miR-192-5p targeting the tumor promoting gene ERCC3, upregulated in HCC patients (TCGA data), were studied here. Interestingly, ERCC3 is transcriptionally regulated by TP53, which is usually suppressed by HBx protein, reinstating its importance in HBV infection and HBV present HCC cases (Levine, 1997; Wang et al., 1994; Yang et al., 2014). Moreover, hsa-miR-192-5p is considered to be a cancer stem cell biomarker whose downregulation is associated with HCC carcinogenesis (Fu et al., 2021). Our bioinformatic analyses suggest that the ERCC3 expression change in HCC could be due to the function of hsa-miR-192-5p. The reason behind ERCC3 dysregulation in HCC could be attributed to the downregulation of hsa-miR-192-5p. However, further in vitro studies are required to validate this mechanism.
A set of unidirectionally regulated mRNAs, PIGC, KNTC1, CSTF2, SLC412A, and RAB17, and the hDEmiRs targeting them, hsa-miR-98-5p, hsa-miR-193b-3p, hsa-miR-142-5p, hsa-miR-522-5p, and hsa-miR-370-3p, respectively, were identified from both the GEO and the TCGA data (Fig. 5). Among them, hsa-miR-98-5p, hsa-miR-193b-3p, and hsa-miR-142-5p are anti-tumorigenic miRNAs in HCC with a role in cell migration (Fei et al., 2020; Lou et al., 2017; Tsang et al., 2015; Wang et al., 2020). KNTC1, PIGC, and CSTF2, which are targets of downregulated hDEmiRs, were similarly upregulated in both GEO and TCGA datasets. The function of these genes could be aiding in HCC development, and hence, targeting them could inhibit HCC progression. KNTC1 overexpression has been tied to poor prognosis and higher cell proliferation through PI3K/Akt pathway regulation (Tong et al., 2023). In contrast, PIGC overexpression promotes proliferation and induces cell migration in HCC (Guo et al., 2021). Meanwhile, CSTF2 expression promotes HCC through enhanced aerobic glycolysis (Chen et al., 2022). On the contrary, SLC41A2 and RAB17, the targets of upregulated hDEmiRs, were downregulated in both TCGA and GEO datasets. In this context, overexpression of RAB17 is considered to inhibit HCC progression through G1 cell cycle arrest (Wang et al., 2015). However, more studies are required to understand the particular role of SLC41A2 in HCC. Quite surprisingly, in addition to HCC, all these five hDEmiRs were expressed in a similar pattern in HBV infection, firmly establishing the role in HBV infection advancing to HCC progression.
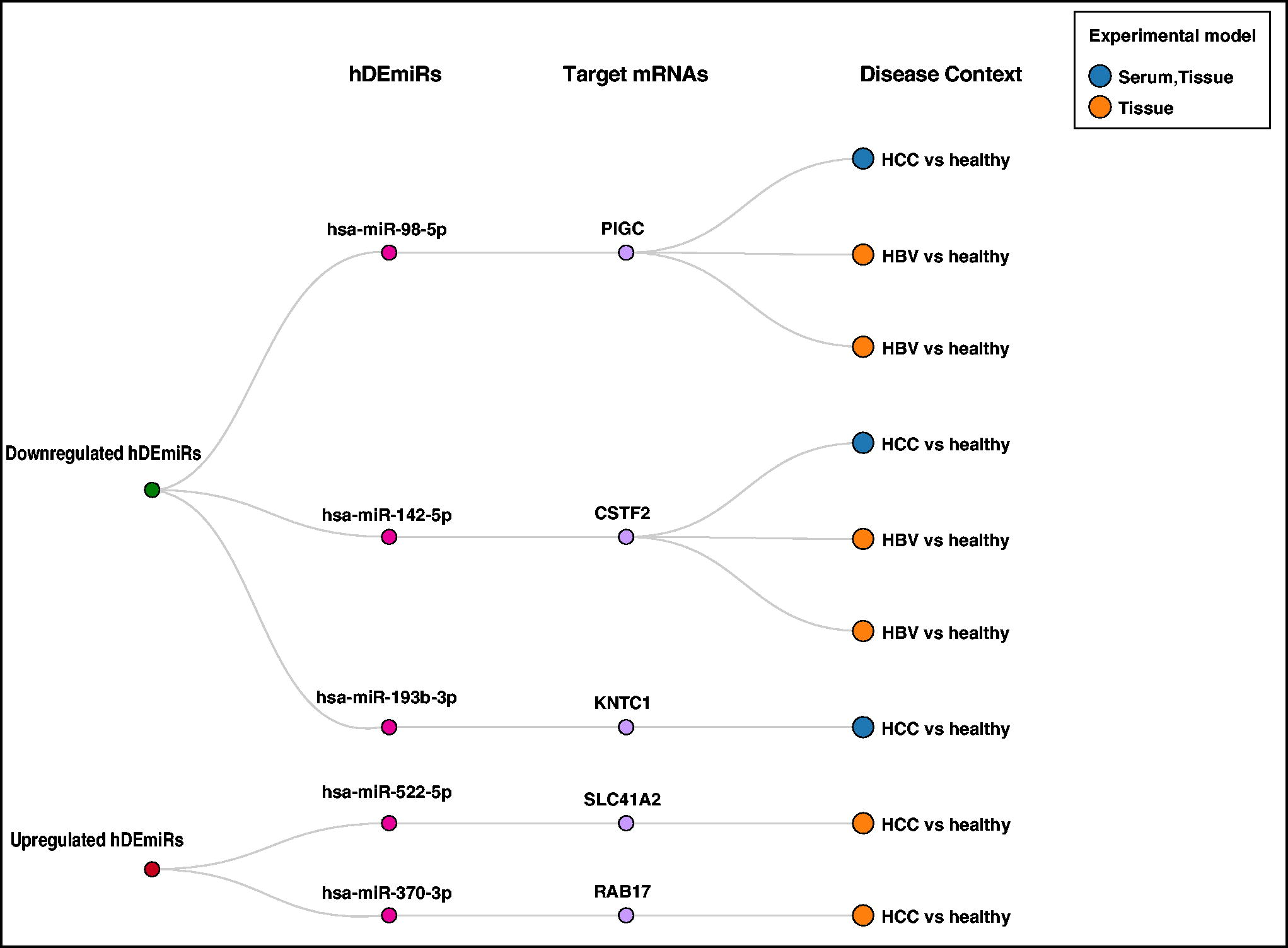
GEO-TCGA analysis—the hDEmiRs targeting those mRNAs unidirectionally regulated in GEO and TCGA are depicted along with the disease context in which the hDEmiRs are studied, the experimental model, and their expression pattern.
This study illuminates the intricate relationships between the miRNAs and their target genes in fine-tuning the liver pathologies’ overall mechanism. Such a cassette of miRNA expression in different disease conditions offers promise for biomarker development for HBV-induced pathophysiologies and to develop miRNA-based therapeutics.
Currently, many tools and databases explain the roles of miRNAs in cancer types and their differential expressions. However, our analysis showed that not all the miRNA perturbations have a downstream effect on modulating the pathology, which could be due to the cell specificity of genes and miRNAs. In this context, machine learning approaches may be used to deliver improved methods and algorithms for the identification of preferential mRNAs based on the impact of other noncoding RNAs (Thomas et al., 2024). Moreover, the availability of longitudinal datasets across liver pathologies could be useful in deriving robust mechanisms of miRNA regulation and its consequences in the future.
Conclusions
We identified the differentially regulated miRNAs in HBV-related pathologies in humans, ranging from HBV infection, fibrosis, cirrhosis, ACLF, and HCC. It is noteworthy that only some of the reported miRNAs may be involved in the pathologies induced by HBV, whereas others may experience changes over time or have their effects cancelled out by complex cellular regulations. Thus, the actual effect of these miRNAs, which could indeed reflect in the HBV infection advancing into fibrosis, cirrhosis, and HCC, needs to be investigated in detail. To address this, data from GEO and TCGA were used to compile a comprehensive resource on functional miRNAs and their effects on respective targets, aiming to advance our understanding of the infection mechanism.
Through bioinformatics analysis, five mRNAs, namely, PIGC, KNTC1, CSTF2, SLC412A, and RAB17, regulated unidirectionally in HCC patient data were found to be potentially targeted and modulated by five unidirectionally expressed hDEmiRs, hsa-miR-98-5p, hsa-miR-193b-3p, hsa-miR-142-5p, hsa-miR-522-5p, and hsa-miR-370-3p, respectively. However, the role of these miRNA–mRNA interactions, which was similar across different datasets, has not been validated in HBV-induced pathologies, including HCC. Hereby, we present the first of its kind compendium of hDEmiRs in HBV-related liver pathologies and analyzed their functional role in HBV-induced HCC cases calling for further validation studies that could aid in diagnosis and treatment of HBV pathologies.
Footnotes
Acknowledgments
We thank the Yenepoya (Deemed to be University) for providing the necessary facility to the Centre for Integrative Omics Data Science (CIODS), to carry out this study.
Authors’ Contributions
K.R.: Writing—original draft (lead), formal analysis and data curation (lead), writing—reviewing and editing (equal). R.V.: Writing—reviewing and editing (equal). R.R.D.: Data acquisition and software (lead). R.R.: Methodology (lead) and writing—reviewing and editing (equal). N.R.: Conceptualization (lead), writing—original draft (supporting), and writing—reviewing and editing (equal).
Author Disclosure Statement
The authors declare that there are no conflicting financial interests.
Funding Information
No funding was received for this study.
Abbreviations used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.