
Abstract
Bacteriophages offer a viable solution to addressing the global issue of bacterial resistance to antimicrobials. Although knowledge of bacteriophages has increased greatly since their discovery in 1915, a significant amount of what is currently known is based on studies conducted in model conditions and aerobic environments. There are a variety of environments in which bacteriophages could be applied to successfully replace or supplement antimicrobials in agriculture, food production, and human medicine where the amount of oxygen is limited. There is a need to use phages in oxygen-limited environments, but few studies have examined the impact oxygen-limited environments have on the ability of phages to kill their hosts. The work that has been done is, however, insightful and will likely stimulate this area that is growing in importance as our need to use phages grows. This review summarizes the studies to date that have reported the characteristics of phages in both oxygen-rich and oxygen-limited environments. We also discuss the importance of considering the ultimate environment a phage will be applied to when designing experiments to isolate and characterize phages for use in phage-based antimicrobial products.
Introduction
With the threat of bacterial resistance to antimicrobials constantly growing in magnitude, interest in using bacteriophages (phages) as alternative therapeutics and treatments has increased. The use of phages as antimicrobials in agriculture, food production, and human medicine requires reliable and effective phage infection in a wide range of environments where, in many cases, oxygen is present at a low concentration, if at all. Examples of oxygen-limited environments range from the bloodstream, wounds, the urinary tract (UT), and the gastrointestinal (GI) system in humans and animals, to vacuum-packed (VP) food products and food stored in modified atmosphere packaging (MAP). Despite this wide range of oxygen-limited environments, the impact of oxygen availability on phage infection is notably understudied and consequently there is a limited understanding of how oxygen availability impacts the ability of phages to infect their hosts.
Differential infection with respect to oxygen availability is mainly an issue for facultative anaerobic bacteria that have adapted so efficiently to survive, and in many cases thrive, in both aerobic and anaerobic environments. The majority of bacteria that comprise the World Health Organization's (WHO) priority list of multidrug resistant pathogens and list of foodborne bacterial pathogens are facultative anaerobes, indicating that these kinds of bacteria are critical targets for alternative treatments in human medicine.1,2
Phages are approved for use as additives to several food products to prevent the growth of foodborne pathogens. 3 Further, there is a significant amount of literature, reviewed in more detail by Abedon et al., Cisek et al., and Dec et al.,4–6 that provides evidence that phages could be used safely and efficiently to treat bacterial disease in animals and humans. However, even with all of the supporting data currently available on developing phage products, more evidence is needed to demonstrate the efficacy of phages, defined as the ability of a phage to reduce the number of targeted bacteria, 7 in vivo. This will contribute greatly to establishing the potential impacts that phages may have on humans or animals before they are more widely approved for therapeutic use, especially in medical practice.5–7
Based on the reduced oxygen availability in the final environments in which phage-based treatments will be applied, such as whole blood and the UT, the scarcity of published studies that examine phage infections in oxygen-deficient environments is surprising. In most phage studies, phages are isolated and their phage–bacterial host interactions are examined based on the optimal lab environment for bacterial growth. These traditional laboratory protocols provide the best possible growth environments for the bacterial host, but the conditions do not always imitate the final environment into which the phage will be introduced and will have to remain effective. This can lead to a situation where the efficient killing of bacteria by phages in vitro has been established, but when tested in vivo against the same host the phages do not perform as well. This scenario was demonstrated by Tsonos et al., 8 who highlighted that conventional in vitro methods of phage isolation, characterization, and selection in the development of a therapeutic phage cocktail do not always lead to efficient phage cocktail activity and successful reduction of bacterial growth in vivo. This study demonstrated the importance of considering the final application of a phage when designing a phage-based product.
Both phage stability and bacterial growth have the potential to impact the efficiency of phage infection. Nutrient availability, pH, temperature, and oxygen availability are widely understood and have been shown in a variety of studies to induce adaptive responses and impact the growth of numerous species of bacteria.9–14 There are also many studies, reviewed in more detail by Jończyk et al., 15 which demonstrate that temperature, nutrients, and pH or combinations of these factors can affect the ability of a phage to successfully infect its host.16–29 Unfortunately, none of these studies investigated the stability of phages in environments where oxygen is limited.
Oxygen limitation has been shown to alter gene expression in bacteria, which, in some cases, results in changes in metabolic pathways, the expression of bacterial cell surface proteins, or can lead the bacterial cell to enter a dormant state.30–34 Completion of the lytic phage cycle requires the successful interaction between a phage and its host in several different ways, including adsorption of phage virions to host cell surface receptors and replication of the phage genome by the host cell. Adaptive responses in the bacterial host induced by limited oxygen availability could impede the ability of the phage to infect the host cell under such conditions, reducing the efficiency with which it can lyse its host or even preventing the successful completion of the lytic cycle entirely. 35
A more in-depth review of the phage infection mechanisms that could theoretically be impacted by changes in the bacterial host induced by oxygen limitation was recently conducted by Hernández and Vives. 34 In this review, they highlight the lack of information available on phage infection in anoxic conditions for both obligate and facultative anaerobes. They also theorize, based on current literature, as to which physical and metabolic changes in facultative anaerobic hosts induced by oxygen limitation have the potential to impact phage infection.
The specific conditions associated with the environment that a phage will be applied to and need to work in should be considered when isolating and characterizing phages for the development of products and treatments with specific end-point applications. This process should include characterization under different levels of oxygen availability, as such environments can have a significant impact on the behavior of the bacterial host and thus, the ability of the phage to infect it. In addition, standardized methods should be implemented to quantify phage virulence under different conditions. This review presents a summary of the limited data that are currently available on the effect of oxygen limitation on phage infection, and in consequence the limited understanding of this topic. It also highlights the fact that no single methods or metrics are currently used to quantify phage virulence.
Literature Selection Method
To identify studies that specifically mention findings relating to the effects of oxygen limitation on the characteristics of phages and the infection of their hosts, a search using Biopython's 36 Entrez module with specific search terms was carried out. First, all of the articles that have been published up to and including August 13th, 2020 with the word “phage” or “bacteriophage” in the title or abstract were extracted from PubMed, resulting in a collection of 81,986 publications. Thereafter, this collection of articles was further filtered with any of the following terms: “hypoxic,” “normoxic,” “aerobic,” “anaerobic,” “anaerobiosis,” or “oxygen-limiting.” This search returned 473 articles that were then manually assessed for their relevance to the topic of this review. Articles were selected for inclusion in this analysis if the study investigated phage/host interactions or phage characteristics under both aerobic and oxygen-limited conditions.
General Overview of Selected Literature
In total, 16 articles35,37–49 were selected and are discussed in more detail in this section, but a specific summary of the findings for each study is shown in Table 1 and a visual summary of the overall findings is presented in Figure 1. The majority of phages studied across all of the articles reviewed here infect Escherichia coli, and interestingly for most of these phages there was no difference observed in their virulence or infection characteristics between aerobic and anaerobic environments. In the majority of the selected articles, the phages are described as strictly lytic.37,39–41,44–48,50 In the remaining studies, the life cycles of the phages were either described as temperate 35 or were not mentioned at all, which tended to occur in the older studies.38,42,43,49,51
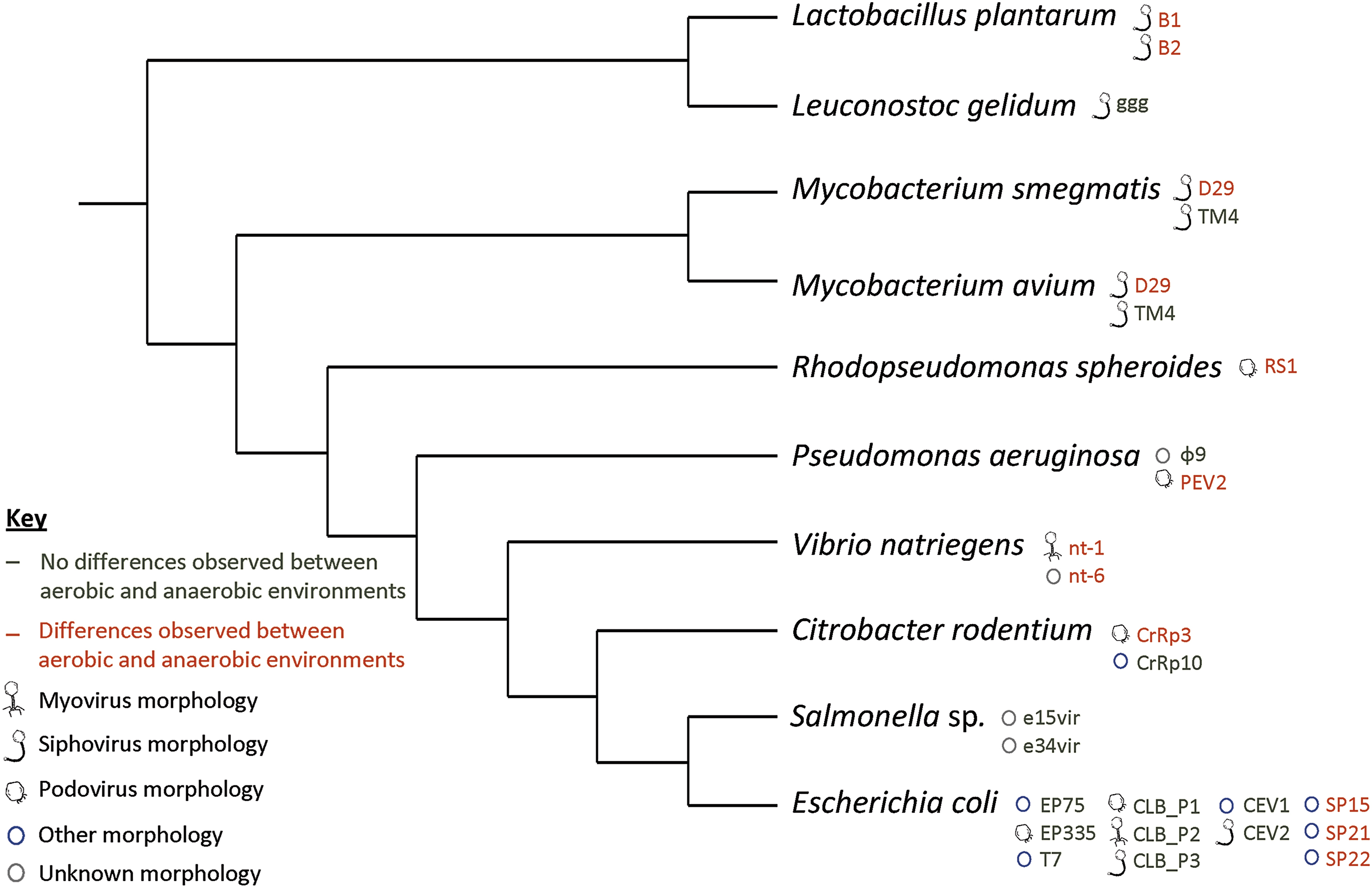
An unscaled phylogenetic tree of the bacteria used in the studies based on the rpoB gene encoding the β subunit of bacterial RNA polymerase. Bacterial species are labeled alongside the phages that have been included in this review, as they have been examined to determine the impact of oxygen limitation on phage infection. The parsimony tree was inferred from a clustalw 60 protein alignment by using SeaView. 61
Summary of Studies and Their Findings Related to the Effect of Oxygen Availability on Phage Infection a Organized by Publication Year from Oldest to Most Recent
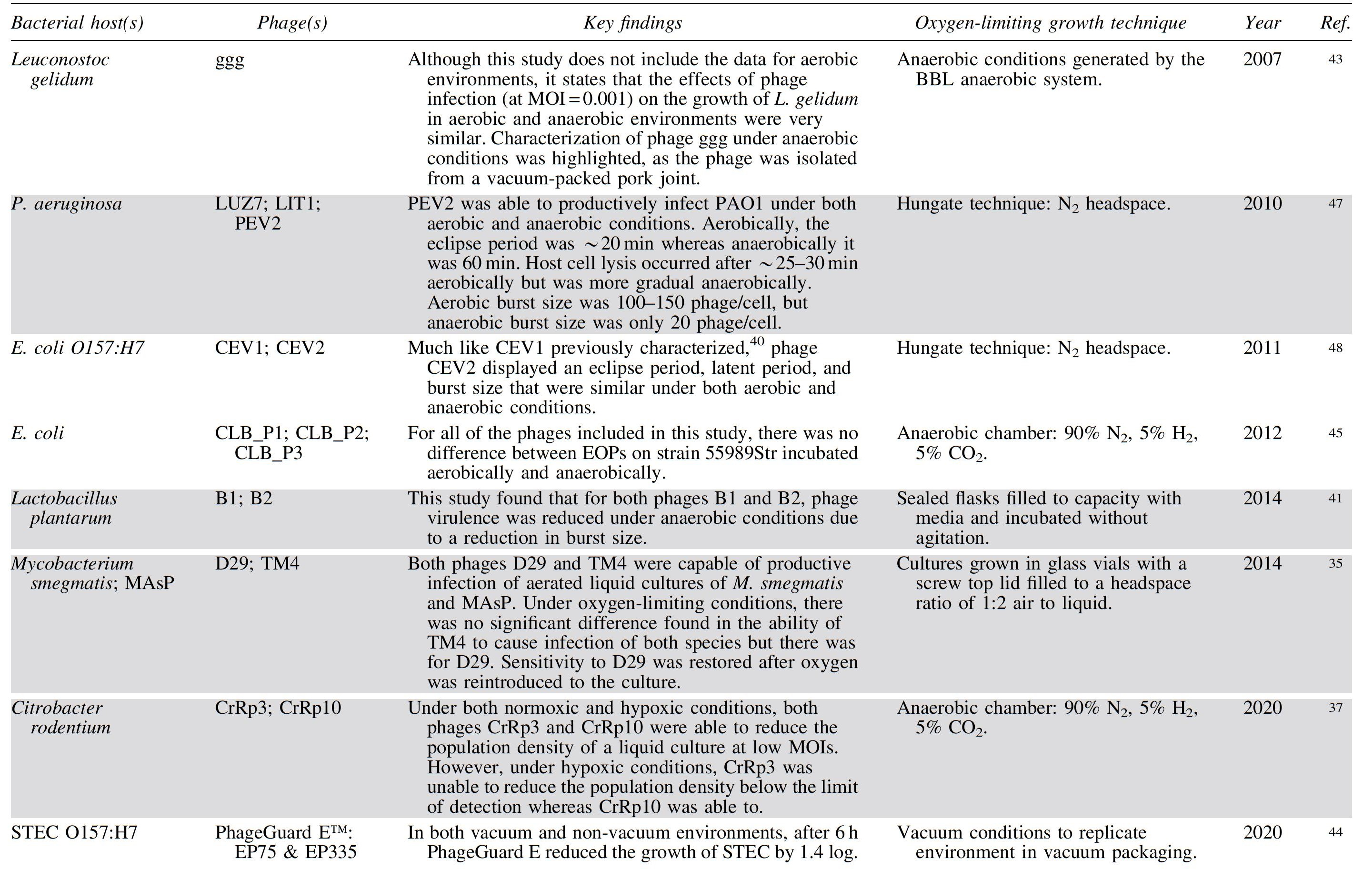
Studies in which differences in phage infection were observed between aerobic and anaerobic conditions are highlighted in gray; studies where no differences were observed are in white.
EOP, efficiency of plating; MAsP, Mycobacterium avium subspecies paratuberculosis; MOI, multiplicity of infection; STEC, shiga toxin-producing E. coli.
The definitions of anaerobic conditions vary between the studies, with some authors using standard anaerobic conditions maintained within anaerobic chambers whereas others use oxygen limiting growth techniques, varying culture surface areas, or sealed bottles with nitrogen headspace to create environments with lower oxygen availability. The method used to generate anaerobic conditions in each study is detailed in Table 1.
In general, these studies use standard characteristics to compare phage infection under different oxygen concentrations such as burst size, latent period, efficiency of plating (EOP), and adsorption rate, all of which are widely accepted as measures of phage efficiency. In some cases, the difference in phage infection was demonstrated by plaque size 38 or the ability of phage to reduce the optical density (OD) of a liquid culture.37,50 Although there are clearly methodological differences within the studies in terms of measurements used for efficiency of phage infection and virulence and classification of anaerobic environments, all of these studies present findings that demonstrate the effect of oxygen availability on phage infection.
The studies are split into two main groups: studies demonstrating that oxygen availability alters phage infection characteristics and studies where oxygen availability was not found to affect phage infection characteristics. Within those groups the studies are, in general, grouped together based on the method by which the phage was characterized, or its virulence measured, for example, burst size or EOP. In some cases, studies have been grouped based on the application of the phages investigated.
Studies Demonstrating Oxygen Availability Alters Phage Infection Characteristics
For nine of the studies included in this review there were differences in phage infection characteristics under aerobic conditions compared with anaerobic conditions.35,37,39,41,42,47,49–51
Earliest studies
It was established as early as 1925 by Gregory Shwartzman that oxygen availability impacts the efficiency of phage infection. 51 In this study, Shwartzman used test tubes with different diameters to investigate the effect of oxygen availability on the ability of the phage to propagate on their hosts. They concluded that “partial anaerobiosis,” introduced by limiting the surface area of the culture, pertains to the successful regeneration of the phage added at a very low concentration. 51 After the investigation of “partial anaerobiosis” on the lytic capabilities of phages, Shwartzman used the same host/phage combination to study the effect of oxygen on the interactions between the phage and bacteria. 49 Here, Shwartzman observed that a reduction in the efficiency of phage infection (measured by the turbidity of plaques) was detected aerobically at pH 7.6 and anaerobically pH 7.049 and concluded that oxygen availability influences the behavior of the bacteria and, as such, the ability of the phage to infect it. Both of these studies, published relatively soon after the discovery of phages, provide early indications that the availability of oxygen influences the ability of phages to successfully infect their hosts. 51
Quantification of phage virulence with burst size and latency period
It was not until, almost 50 years later, in 1974 that the next study to investigate the effects of oxygen limitation on phage infection was published. This is likely due to the loss of interest in phage research and then a resurgence in its popularity. This later study, conducted by Abelovich and Kaplan, was driven by studies,52,53 which showed that light and oxygen availability impacted the growth of Rhodopseudomonas spheroides. Based on this, they investigated the effect of oxygen availability on phage infection characteristics for phage RS1, an R. spheroides phage. 42 They found that at low multiplicity of infection (MOI) there was reduced adsorption and ability of the phage to infect its host under anaerobic growth conditions; however, this ability was restored when the MOI was increased to 10.
They also found that the latent period was 120 min aerobically and extended to 150 min anaerobically but that under both conditions the burst size remained the same. Abelovich and Kaplan concluded that, collectively, their results suggest that anaerobic growth of the bacterial host influences the ability of the phage to adsorb to and infect the host cell, but once successful infection has occurred, the anaerobic growth conditions have no impact on the ability of the phage to replicate. 42
Interestingly, from the articles we reviewed, the study by Abelovich and Kaplan 42 was the only phage-system studied where the latent period was extended under anaerobic conditions, but the burst size remained the same. A study published by Zachary in 1978 on two Vibrio natriegens phages, nt-1 and nt-6, showed that when infecting the same host strain, both nt-1 and nt-6 had longer latent periods when grown under anaerobic conditions, 39 much like what was seen for phage RS1. 42 However, in contrast to the findings of Abelovich and Kaplan for phage RS1, 42 nt-1 had a larger burst size when grown anaerobically, whereas nt-6 had a smaller burst size under anaerobic conditions. 39
Other studies that observed a reduction in the burst size for different phages when grown under anaerobic conditions include: Silva and Sauvageau, 41 where they showed that anaerobic growth reduces the burst size for both Lactobacillus plantarum phages B1 and B2; and Ceyssens et al., 47 where Pseudomonas aeruginosa phage PEV2 was shown to have a longer latency period and smaller burst size under anaerobic growth conditions.
Quantification of phage virulence using OD
A recent study conducted by Mizuno et al. 37 looked at the effect of aerobic and anaerobic growth conditions on the ability of phages CrRp3 and CrRp10 to infect Citrobacter rodentium. They showed that both phages were able to productively infect their host during aerobic and anaerobic growth, but under anaerobic conditions and at low MOIs CrRp3 was unable to reduce the OD as low as it could under aerobic conditions. They also showed that resistance to CrRp3 occurred under both growth conditions but that CrRp10 was more resilient to the development of resistance in its host as no regrowth was observed in any cultures infected with CrRp10 under either condition for the duration of the experiment.
A similar technique of evaluating phage infection under anaerobic and aerobic conditions was used in an investigation into the use of phages to control E. coli O157:H7 growth in the GI tract of mice conducted by Tanji et al. 50 They found that in vitro, the three phages included in the study; SP15, SP21, and SP22, were both independently and in combination as a cocktail able to prevent the growth of E. coli O157:H7 more efficiently when grown aerobically, represented by a larger reduction in OD, than when they were grown under anaerobic conditions. They also found that under anaerobic conditions there was a larger regrowth of bacteria after the addition of a cocktail of all three phages than the regrowth that was observed under aerobic conditions. This indicates that the development phage resistance occurred sooner and more aggressively during anaerobic growth.
Overall, the study showed that when combined, the three phages were effective at reducing growth of E. coli O157:H7 in the GI tract of mice. However, despite in vitro studies indicating a more rapid development of resistance under anaerobic conditions, this study did not test the susceptibility of E. coli O157:H7 cells that were recovered from the mouse gut to the phage cocktail. Having such data in future would be really useful, as they would demonstrate whether the development of bacterial resistance to phages occurs as rapidly, if at all, in vivo and would provide more insight into the suitability of certain phages for therapeutic use.
A specific study of mycobacteriophages and oxygen limitation
A study of mycobacteriophages by Swift et al. 35 used less conventional methods to demonstrate the effects of oxygen limitation on the ability of phages to infect their host. In these mycobacterial studies, a phage amplification assay (PAA) can be used as a faster alternative to normal viable cell counts for quantifying mycobacterial cells. 54 The aim of their study 35 was to investigate how different growth states of mycobacteria affected the ability of certain phages to infect the host cells. Subsequently, they demonstrated the consequential effect this had on the sensitivity of the PAA and thus the reliability of this method of tuberculosis diagnosis. Their study demonstrated the effect that growing the host in an oxygen-limiting environment has on the ability of different phages to infect its host in comparison to cells grown in aerated conditions. Under anaerobic conditions, the sensitivity of the PAA was significantly lower for both Mycobacterium smegmatis and Mycobacterium avium subspecies paratuberculosis (MAsP) when using phage D29 and the sensitivity was restored on reintroduction of oxygen to the growth environment. Their results indicate that D29 is unable to efficiently infect bacteria in a dormant state induced by oxygen limitation.
Our current knowledge and understanding of phage biology leads us to hypothesize that the resulting changes in certain characteristics of phage infection are specific to each phage and the relationship it has with each of its host strains, and collectively these studies support this idea. Bacterial adaptation to different atmospheric growth conditions can affect components of the bacteria,9,30–33 such as its cell wall and DNA replication, which can be utilized by the phage for adsorption, infection, replication, and, ultimately, the completion of its infection cycle. Alterations to these components, which arise due to environmental stresses such as reduced oxygen availability, can cause variation in the infection characteristics of different phages as a result of changes to the bacterial components that phage infection mechanisms exploit.
Studies Where Oxygen Availability Was Not Found to Affect Phage Infection Characteristics
For seven of the studies included in this review, there were no differences observed between the infection characteristics of phages under aerobic conditions compared with anaerobic conditions.38,40,43–46,48
Quantification of phage virulence with EOP
When investigating the anaerobic transfer of antibiotic resistance in P. aeruginosa, Graves and Riggs 40 demonstrated using EOP values that for P. aeruginosa phage, ϕ9, there were no significant differences in the ability of ϕ9 to infect three different host strains under aerobic and anaerobic conditions. A study by McConnell and Wright 38 demonstrated using several Salmonella and E. coli phages that growing the bacterial host under anaerobic conditions resulted in alterations to plaque morphology. They proposed that increasing the generation time of the bacteria by growing them under anaerobic conditions results in the increased size of plaques. 38 Despite the change in plaque morphology, they showed that there were no changes in the EOP values for phage/host combinations grown anaerobically or aerobically. 38
In a different study, Maura et al. 45 found no differences between the EOP of phages CLB_P1, CLB_P2, and CLB_P3 on E. coli host strain 55989Str when they were grown aerobically or anaerobically. Interestingly, contrary to the findings from in vitro experiments, in vivo experiments studying the effect of a single dose of phages administered orally to mice colonized with 55989Str showed that the phages were able to replicate, but they were unable to reduce the growth of 55989Str in the gut. 45 This suggests that in this case there are other contributing factors that affect phage infection in the murine GI tract.
Quantification of phage virulence with burst size and latency period
Two studies conducted by Raya et al.46,48 showed that for E. coli O157:H7 phages CEV146 and CEV248 there was no difference observed in the eclipse period, latent period, and burst size between cultures grown aerobically and anaerobically. They also showed that the application of CEV1 alone 46 and the combination of both CEV1 and CEV248 to the sheep gut significantly reduced the levels of E. coli O157:H7 throughout the GI tract.46,48
Characterization of phages for food safety applications
A study by Greer et al. 43 showed that there were no differences between host–phage interactions for phage ggg and its Leuconostoc gelidum host aerobically and anaerobically. The publication does not include the specific data for aerobic growth, as the phage was isolated from an anaerobic environment. Therefore, characterization of the phage was conducted largely under anaerobic conditions, but it is noted that there was no difference in phage infection in broth cultures under anaerobic and aerobic conditions. 43
The most recent study included in this review was conducted by Shebs et al., 44 who investigated the ability of PhageGuard E™ to reduce the growth of shiga toxin-producing E. coli (STEC) on the surface of beef packaged under vacuum and non-vacuum conditions. Although this study does not specifically characterize any of the phages that comprise the PhageGuard E cocktail, it does show that the cocktail is equally effective at reducing the growth of STEC on the surface of beef that has been packaged under a vacuum or using oxygen-permeable packaging.
Discussion
Facultative anaerobes have the metabolic versatility to successfully grow under both aerobic and anaerobic conditions. The flexible metabolic nature of these bacteria demonstrates how adaptable they are to changes in atmospheric environments and leads one to consider whether phages adapt in a similar way under environmental stresses and how this might affect downstream industrial and therapeutic applications of phages. It is interesting to note that the studies summarized earlier did not all come to the same conclusion as to how oxygen-limitation impacts phage infection. Of the total 16 studies, 9 studies demonstrated that there was a change in the characteristics of phage infection in cultures grown anaerobically compared with those grown aerobically,35,37,39,41,42,47,49–51 whereas 6 studies found no differences,40,43–46,48 and the remaining study found variation in plaque morphology but no difference in EOP. 38
Overall, this emphasizes the specific nature of the relationship between phages and their hosts. Further, some of the studies we reviewed investigated the same species of bacteria with different phages but present opposite findings, that is, one study found no difference in phage infection between aerobic and anaerobic conditions, whereas the other did.40,44–51 This shows how important it is to thoroughly characterize phages that are intended for therapeutic or industrial use. To ensure an optimal outcome, phages should be characterized in conditions as close as possible to the environment they will ultimately be deployed in.
This need for more “physiologically true” characterization may have contributed to the lack of success in two recent clinical trials for phage therapy (PT).55,56 Although these trials were exploratory, complex and much was learned from them, it is true to say that in neither case was evidence published to show in vitro characterization of the phages in oxygen-relevant conditions. Our analysis of data reviewed here leads us to suggest that such characterization may have improved the outcome of the trials. It does seem logical that phages to be used in PT cocktails should be studied extensively in environments that reflect the oxygen availability of the environment where they are needed to be effective.
Measuring phage efficacy
The fact that there is no standard measure to quantify phage virulence makes fully understanding how it is impacted by oxygen availability challenging. The studies summarized here have used different methods to measure the differences in efficiency of infection for their phages. Although it is pertinent to describe the different stages of phage infection (e.g., eclipse period, latent period, burst size) when characterizing the way a phage infects its preferred host, if the purpose of phage discovery is for a practical application it is arguably more important to consider the infection cycle in its entirety by using quantitative measures such as EOP 57 and the recently described virulence index. 58
Although these techniques do not provide specific information on all of the individual aspects of the phage infection cycle, the experiments can be repeated under a variety of different conditions to provide a quantitative value representing the efficacy of phage infection that can be used to compare the activity of the same phage in different environments or the efficacy of different phages infecting the same host. This is key to demonstrating how phages may respond to changes in important environmental parameters and how the entire phage infection cycle is affected.
Applying phage ecology to phage isolation practices
When considering the growing interest in the use of phages for a wide variety of practical applications, a deeper understanding of phage ecology and adaptability may really augment our ability to effectively exploit them. Thinking more broadly about bacterial and phage ecology, in natural environments phages have to respond to and infect bacteria in a range of physiological states and environments if they are to survive. Knowing more about how phages behave in response to changes presented by their bacterial hosts and their environment could, therefore, improve the process of selecting phages.
A method of categorizing phages in a way that considers their behavior in different environments has recently been proposed by Clokie et al. 59 Here, they describe the application of the ecological CSR framework, originally used to categorize plants, to categorize phages into their “functional types.” 59 They hypothesize that the temporal transcription phases of phages correlate with phages being competitors (C), having the ability to survive in stressful environments (S) or the ability to withstand disturbance (R). Their proposed categorization method provides key information about how a particular phage may respond to certain environmental pressures.
Although this is an early proposition for a method by which to categorize phages, the logic presented in this work is supported by several studies, reviewed here, that have characterized phages in multiple environmental conditions. It is helpful to present further information from the CSR analysis with a specific focus on oxygen status, as the framework does seem to make sense with the few studies on phages that do have available transcriptional datasets, supporting the idea that knowing more about phage ecology can inform our ability to utilize phages. These phages are discussed later in terms of how their CSR classification is supported by three studies we have reviewed here. Only three phages are discussed here, as they are the only phages for which the transcriptional data exist that is needed to categorize them by this method.
Mycobacterium phage D29—in their analysis, Clokie et al. categorized D29 as a “C” phage, suggesting that it is a phage that thrives when its host is growing in abundance under stable conditions. 59 As discussed earlier, D29 was shown to be unable to infect M. smegmatis and MAsP under anaerobic conditions when the bacteria was dormant, but could infect both species once aerobic conditions were restored and the bacteria was actively growing again. 35 This supports the “C” phage categorization, as it is a phage that tends to thrive in optimal conditions.
Pseudomonas phage PEV2—this phage was categorized as a “CS” phage, indicating that it is a phage that combines competitor traits along with the ability to thrive despite being in a stressful environment. 59 This is consistent with our observations in the literature where Ceyssens et al. showed that it can productively infect its host bacteria under both aerobic and anaerobic conditions. 47
E. coli phage T7—this is a well-characterized phage and was categorized as an “SR” phage, designating it as a phage that can cope with both stress and disturbances in its environment, for example, changes in oxygen concentration. 59 This categorization is supported by the study by McConnell and Wright, 38 discussed earlier, which showed that growth in anaerobic conditions altered the plaque morphology for this phage but had no impact on the EOP value, suggesting that under anaerobic conditions T7 retained the ability infect its host efficiently. 38
Going forward, we may identify that phages that are particularly good at coping with anaerobic conditions may dominate in specific categories of this framework. It is clearly an ecological trait that is incredibly useful for many practical applications, and that we will hopefully understand better in the future. When searching the literature, we did not find examples of studies where the focus of phage isolation and host range determination was explicitly linked from the outset to the physiologically relevant needs of a facultative anaerobe as the host. It would be of significant interest to investigate whether applying different oxygen concentrations during the phage isolation process using the same environmental sample and the same host resulted in the isolation of different categories of phages under each condition. Perhaps by growing bacteria under optimal conditions we miss out on isolating phages that have adapted to efficiently infect their host when its growth is hindered, and these could be the phages that are best suited to the final environment a phage may be applied to.
Concluding Remarks
The literature reviewed here demonstrates that changes in the availability of oxygen can have a significant impact on characteristics of the phage infection cycle and the ability of phages to productively infect their host. This supports the suggestion that phage researchers must be cognizant of the ultimate application of the final phage product from phage isolation through to characterization of host/phage interactions throughout every stage of experimentation.
This review also highlights the lack of a standard method that is routinely implemented in the characterization of phages to measure phage virulence. A standard approach that demonstrates the characteristics of phages across the duration of their life cycle, such as that previously suggested by Storms et al.,58 would facilitate the comparison of phages in different conditions and streamline phage discovery. Further, we suggest more forethought when designing the phage discovery process to target specific categories of phages that are most likely to perform well under certain conditions. Including studies of the phage-host dynamics under conditions where oxygen is limited is straightforward and can be done by using equipment as simple and inexpensive as a candle jar.
This review has highlighted the impact that oxygen availability can have on phages and their relationship with their hosts. Based on what is already known about phages, we suggest that there is a treasure trove of phages out there waiting to be found; however, the key to isolating the ones we need may lie in more nuanced and “real-world” isolation and characterization procedures.
Footnotes
Authors' Contributions
F.E.H. and M.R.J.C. conceived of the idea for this review. T.S.P. conducted the literature search, and F.E.H. reviewed the findings. F.E.H. prepared the original draft. T.S.P. created the figures with editing by F.E.H. F.E.H., T.S.P. and M.R.J.C. reviewed and edited the drafts. All co-authors have reviewed and approved the article before submission. This article has been submitted solely to this journal and is not published, in press, or submitted elsewhere.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Biotechnology and Biological Sciences Research Council (BBSRC) and University of Leicester funded Midlands Integrative Biosciences Training Partnership (MIBTP) [MIBTP2: BB/M01116X/1] PhD studentship awarded to F.E.H.