
Abstract
Bacterial canker, caused by Pseudomonas syringae, is a devastating disease of stone fruit trees worldwide. The bacterium has a broad host range and a high capacity for adaptation and dissemination, owing to its high mutation rate and horizontal gene transfer. Traditional control methods based on copper compounds and antibiotics have resulted in the development of resistance in the bacterial population. Thus, alternative approaches are needed, such as phage therapy. This study aimed to characterize the physicochemical and biological properties of novel Pseudomonas syringae pv. syringae (Pss)-specific phages isolated from the soils of northwestern Iran. Seventy-five phage isolates were obtained, and their host range was determined against various bacterial pathogens. Five phages exhibiting the highest lytic activity against Pss and a narrow host range were selected for subsequent analysis. The stability of the selected phages was assessed under different conditions such as ultraviolet irradiation, temperature, pH, NaCl concentration, and chloroform exposure. The selected phages demonstrated significant effectiveness in vivo, exerting substantial suppression on the population of Pss. This reduction was observed for both individual phages and when the phages were utilized as a mixture. The findings indicate that phages have the potential to be used as biocontrol agents in agriculture.
Introduction
Pseudomonas syringae is a gram-negative plant pathogenic bacterium that causes destructive diseases in more than 180 plant species. 1 Bacterial canker in stone fruit trees, caused by P. syringae, is a major global concern. Specifically, three pathovars of P. syringae (P. syringae pv. syringae [Pss], P. syringae pv. morsprunorum race1 [Psm1], and P. syringae pv. morsprunorum race2 [Psm2]) have been identified as the primary causes of bacterial canker in stone fruit trees. 2 Managing this disease is challenging owing to the pathogen’s high mutation rates, horizontal gene transfer, and species-specific variability. 3 Traditional methods such as antibiotics and copper compounds have limitations and have led to bacterial resistance and environmental pollution.4,5 Therefore, alternative strategies, such as using bacteriophage (phage) biocontrol agents, are necessary. Phages are viruses that target specific host bacteria without affecting beneficial bacteria. The use of phages as biocontrol agents in biocontrol has the potential to be used as biocontrol than the use of antibiotics and copper compounds. 6 They are usually highly specific to their bacterial host, without causing damage to benefit bacteria such as antagonist bacteria or microflora. 5 Phages have been studied as biocontrol agents for various plant diseases caused by bacteria such as Dickeya solani, Erwinia amylovora, Pectobacterium carotovorum, Ralstonia solanacearum, and Xanthomonas species.7–12 Several phages have also been isolated and described for different pathovars of P. syringae.13–15 This study aimed to isolate, identify, and characterize phages against Pss from stone fruit orchards in different regions of East Azerbaijan, Iran. The effectiveness of these phages in reducing disease severity was evaluated both in vitro and in vivo, to understand phage potential to be used as biocontrol agents in agricultural settings.
Material and Methods
Bacterial strain and culture media
Pss strain K26 National Center for Biotechnology Information (NCBI, accession no. KY569176), which was previously isolated from stone fruit tree orchards in East Azerbaijan, Iran, was used as the host bacterium for the isolation, infection, and propagation of specific phages. 16 The other bacterial strains that were used in this study for the host range analysis are listed in Table 1. Nutrient broth (NB: glucose 1 g, peptone 15 g, sodium chloride 6 g, yeast extract, 3 g, and water 1000 mL) medium with 0.7% agar was used as the soft top agar overlay and spot assay for the plaque assays. 8 SM buffer (SM: 1M Tris-HCl pH 7.5, 100 mM NaCl, and 10 mM MgSO4) was used for phage suspension and dilution.
Selected Bacteria for the Host Range Assay of Isolated Phages Against Pseudomonas syringae pv. syringae
Isolation of bacteriophages from soil
From 2019 to 2021, soil samples were obtained from five locations in East Azerbaijan, Iran (Tabriz [37.9861N,46.4875E], Azarshahr [37.7629N,45.9557E], Ajabshir [37.4852N,45.9382E], Osku [37.9057N,46.1133E], and Marand [38.4008N,45.7597E]) where stone fruit orchards (almond, apricot, plum, peach, sour cherry, cherry) exhibited symptoms of bacterial canker. Soil samples from a depth of 5–30 cm were collected close to trees infected with bacterial canker disease at each location. A minimum of five samples per location were obtained. The samples were subsequently transported to the laboratory for further analysis. The samples were stored at 4°C until the phage isolation. To release the phages from the soil into the buffer, 20 g of each soil sample was suspended in 40 mL of extraction buffer. The flasks were shaken vigorously for 2–16 h at 200 rpm and room temperature. 17 The suspensions were centrifuged at 8000 rpm for 5 min and filtered through a 0.45 μm syringe filter to remove bacterial debris. To enrich phages, Pss K26 bacteria were streaked onto NB agar plates and incubated overnight at 28°C. The next day, the bacterial culture was diluted to 107–108 CFU/mL (OD600nm) in 100 mL of NB media supplemented with 10 mM MgSO4, 10 mM CaCl2, and 10 mM MgCl2. After incubating for 1–2 h at 28°C with shaking at 120 rpm, 2 mL of soil filtrate was added to the conical flask. The flask was incubated overnight at 28°C with shaking at 120 rpm. The supernatant was obtained by centrifugation (4000 g for 10 min) and filtration (0.45 μm syringe filter). The phages were precipitated by adding 20 μL of ZnCl2 to 1 mL of supernatant and incubating for 5 min at 37°C. The phage pellet was collected by centrifugation (4000 g for 5 min) and resuspended in 100 μL of SM buffer. 17
The phages were purified and isolated by the double-layer agar method with slight modifications. 30 The filtered supernatant (100 μL) obtained after enrichment was mixed with 100 μL of Pss (107 CFU/mL) and 10 μL of 50 mM CaCl2, MgCl2, and MgSO4·7H2O to enhance the phage attachment to the bacterial surface. The mixture was incubated in a water bath at 37°C for 15 min, and then added to 5 mL of soft top agar (NB with 7 g/L agar) and poured on an NB agar plate with 1.5% agar after gentle mixing. The plates were incubated at 28°C for 24–48 h to produce plaques. Each single plaque was collected with a sterile loop and resuspended in 2 mL of sterile SM buffer, shaken for 30 min at 120 rpm to release the phage particles. The suspension was centrifuged at 5000 rpm for 10 min, and the supernatant was filtered through a 0.45 μm syringe filter.
Host range assay
The phage host specificity range was tested using a spot assay test. Table 1 lists the bacterial strains that were utilized. The selected bacteria (Table 1) have diverse ecological roles and impacts on plants, animals, and humans. Some of them are pathogenic and cause diseases in plants or humans, such as Erwinia amylovora, Pectobacterium carotovorum, and Escherichia coli, whereas some of them are beneficial and have commercial or probiotic applications, such as Bacillus subtilis, B. mycoides, and B. licheniformis. Spots (10 μL) of each phage suspension (104 Pfu/mL) (Pfu; plaque forming unit) were applied, and plates were incubated at 28°C for 18–24 h. Clearing phage application zones indicated a positive host range, whereas no lysis zone indicated a negative range. The experiment was repeated twice.18,19
Physicochemical assay
The isolated phages were subjected to various environmental conditions to evaluate their stability such as different pH levels (pH 2, 5, 7, 9, and 12), different temperatures (−20°C, 4°C, 37°C, 45°C, 55°C, 60°C, 70°C, 80°C), and different NaCl concentrations (5%, 10%, and 15%). All the assays were performed for a duration of 1 h.11,18,20
Also, the chloroform stability of the phages was assessed by adding 200 μL of chloroform to 1 mL of phage suspension in SM buffer (103 Pfu/mL) and incubating for 1 h at room temperature. The control sample was treated with SM buffer only. The phage concentrations were determined in Pfu/mL using the double agar layer method. 20 Finally, to assess the impact of ultraviolet (UV) radiation, phage suspensions at a final concentration of 103 Pfu/mL were mixed with SM buffer and poured onto Petri dishes. The suspensions were then subjected to UV light exposure using a UV lamp (55W UV) emitting wavelengths of 290–320 nm for durations of 0, 5, and 15 min. Subsequently, samples were collected, and phage concentrations were determined using the double agar layer method. 20
All stability tests were performed in triplicate, and the phage concentrations were calculated accordingly.
Bioassays
The influence of phages on the growth of Pss was assessed by the killing curve assay. Pss (OD600nm = 0.1) in NB culture was mixed with each phage dilution (105 to 107 Pfu/mL) in SM buffer. The samples included BC (bacterial control), PC (phage control), individual phages, and phage cocktails (a combination of five phages). These five phages were selected based on their plaque size, clarity, and physicochemical stability. To draw the killing curve, the growth rate of the bacterial population in each treatment was determined by measuring the optical density at 600 nm (OD600nm) of the culture at different time points (0, 2, 4, 6, 8, 10, 12, 14, 18, 24, and 48 h) using a spectrophotometer. The culture was incubated at 28°C and shaken at 200 rpm. The experiment was replicated three times, and each treatment had four replications.6,19
For the greenhouse experiment, a randomized design with five treatments and five replicates was used. The temperature of the glasshouse was set at 28°C with a 16/8-h light/dark cycle and 50%RH. The treatments were five single phages, a phage cocktail, a control with phage buffer, dH2O, and a positive control with Pss inoculation. Cherry twigs of a native cultivar (Shabestar) were grown in pots and inoculated with Pss and phages following the method of Hulin et al.
3
Each twig had five wounds with different treatments. The positive control had Pss without phage spray, whereas the negative control had phage buffer or dH2O without Pss. The pots were covered with plastic wrap, incubated in the dark for the first 24 h, and then placed under greenhouse conditions.19,21 The symptoms of the disease on the branch surface were examined, and the disease severity percentage was assessed according to the following formula:
Results
Isolation of bacteriophage from soil samples
This study aimed to isolate specific phages against Pss, the causal agent of stone fruit bacterial canker. Soil samples were randomly collected from around stone fruit trees showing symptoms of the disease and enriched for phage presence. A total of 56 soil samples were analyzed for the presence of phage using the double layer agar method (Fig. 1). The results showed that phage plaques were observed only in the soil samples from Tabriz, Marand, and Azarshahr regions, whereas no phages were identified in the soil samples collected from the Osku and Ajabshir regions. A total of 75 phage isolates were obtained and preserved at a temperature of 4°C. These plaques were segregated into the following four distinct groups based on their size and clarity: large (8–10 mm), medium (5–7 mm), small (2–4 mm), and very small (<2 mm) (Table 2).
The plaque formation of phage isolates from the soils of northwestern Iran using the soft agar overlay technique. The clear zones indicate the lysis of Pseudomonas syringae pv. syringae by the phages.
Phenotypic Characteristics of Five Isolated Phages from the Soils of Northwestern Iran Against Pseudomonas syringae pv. syringae
Bacteriophage host range
The results of the host range assay showed a differential ability to infect and lyse pathogenic bacteria and beneficial bacteria. However, among 75 phage isolates, 5 phages—despite having high lethality on Pss—were unable to infect beneficial bacteria such as P. reinke, B. mycoides, B. simplex, B. licheniformis strain MA-76, B. safensis, B. subtilis strain RN40, B. siamensis strain CBxLGR21, B. tequilensis strain N4-1, and other pathogenic bacteria. These five phages, named MAH-15, AZA-17, AZG-19, AZB-20, and AZZ-21, were selected based on the host specificity range and further characterized (Table 2).
Physicochemical assay
The pH stability profile of the five studied phages exhibited the highest levels of stability at pH 7.0 and 9.0 while displaying diminished stability at pH 5.0 (77% reduction). Furthermore, at pH 12.0, a decrease of 35% in phage titers was observed. Notably, all phages were rendered inactive at pH 2.0 (Fig. 2a). Regarding temperature stability, the range spanned from −20°C to 65°C, with an optimal stability observed between 4°C and 25°C. At temperatures of 37°C, 45°C, 55°C, and 60°C, phage viability was diminished by 30%. However, complete inactivation occurred at 70°C and 80°C (92% reduction). The disparity between the control temperature of 25°C and the temperatures of 70°C and 80°C was statistically significant (Fig. 2b). The UV irradiation stability of the five phages was notably low, with a 71% reduction after 5 min and complete inactivation after 15 min (Fig. 2c). The stability of the phages under NaCl exposure varied, with a 55% reduction at a 10% NaCl concentration and a 78% reduction at a 15% NaCl concentration. Significantly different results were observed between the control phage (without NaCl) and the phage samples exposed to NaCl (Fig. 2d) (one-way analysis of variance, Dankan, p > 0.05). Interestingly, the stability of the five selected phages was unaffected by chloroform treatment. No significant difference was observed between the control phages (without chloroform) and the treated phages (data not shown).
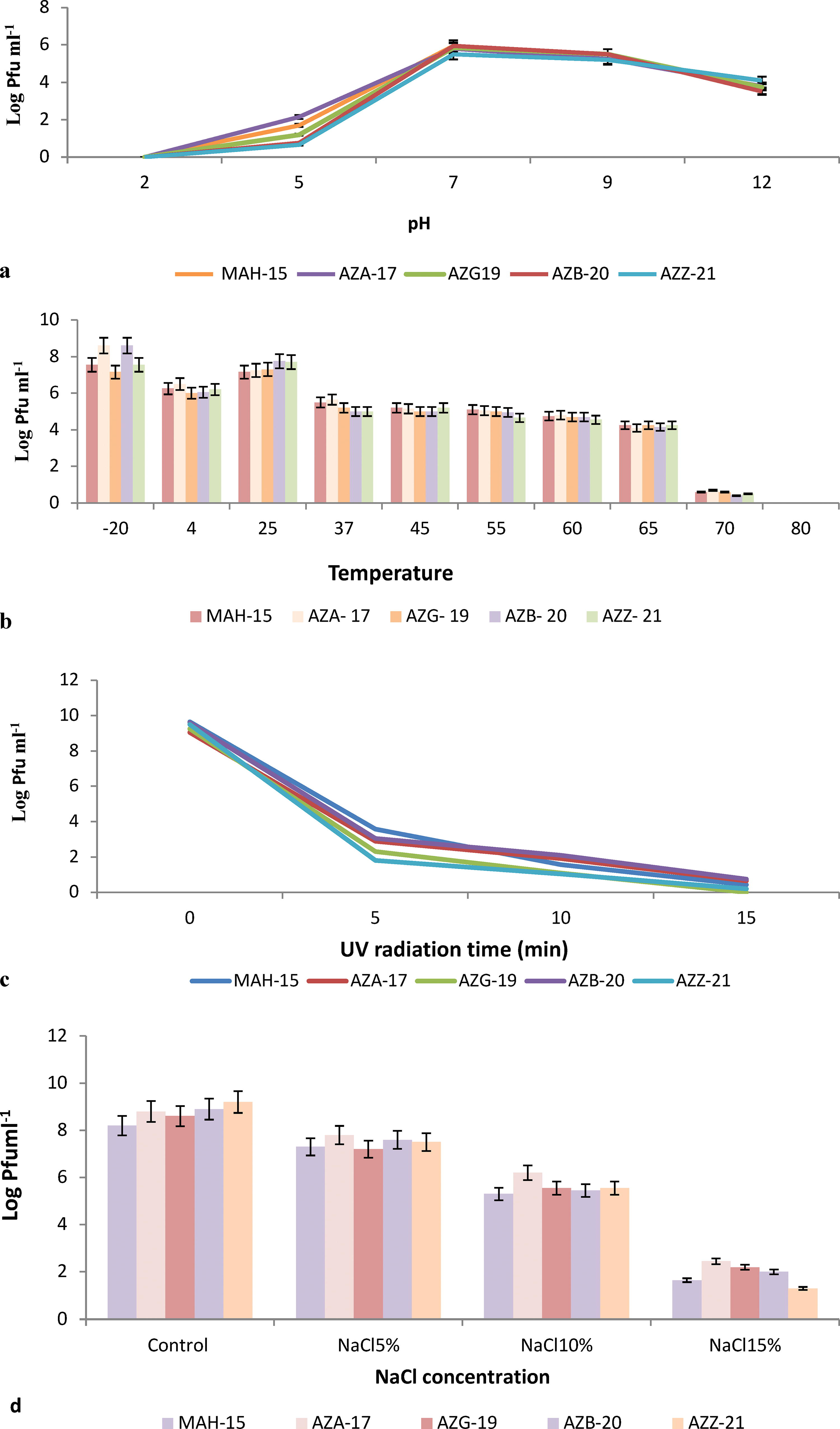
Results of physicochemical assays on five isolated phages from soils of northwestern Iran in in vitro conditions:
The killing curve and glasshouse assays
The evaluation of phage virulence was conducted through the formulation of killing curves using individual phages and a phage cocktail during infections against Pss (Fig. 3). As demonstrated, all phages exhibited the capacity to diminish the Pss population, whereas certain phages (e.g., AZB-19) and the phage cocktail displayed greater efficacy compared with others. Notably, no bacterial resistance to phages was observed after an incubation period of approximately 48 h. To assess the in vivo efficacy of the five phages in reducing Pss damage, cherry twigs were subjected to greenhouse conditions and subsequently examined after 30 days (Fig. 4). Remarkably, both individual phages and the phage cocktail demonstrated significant reductions in Pss damage (p < 0.05). The phage cocktail particularly achieved the highest reduction rate (85%) among all treatments. Following the application of the cocktail, phage AZB-20 exhibited the highest reduction rate (82%), followed by AZG-17 (71%), MAH-15 (68%), AZA-19 (65%), and AZZ-21 (31%) (Fig. 5).
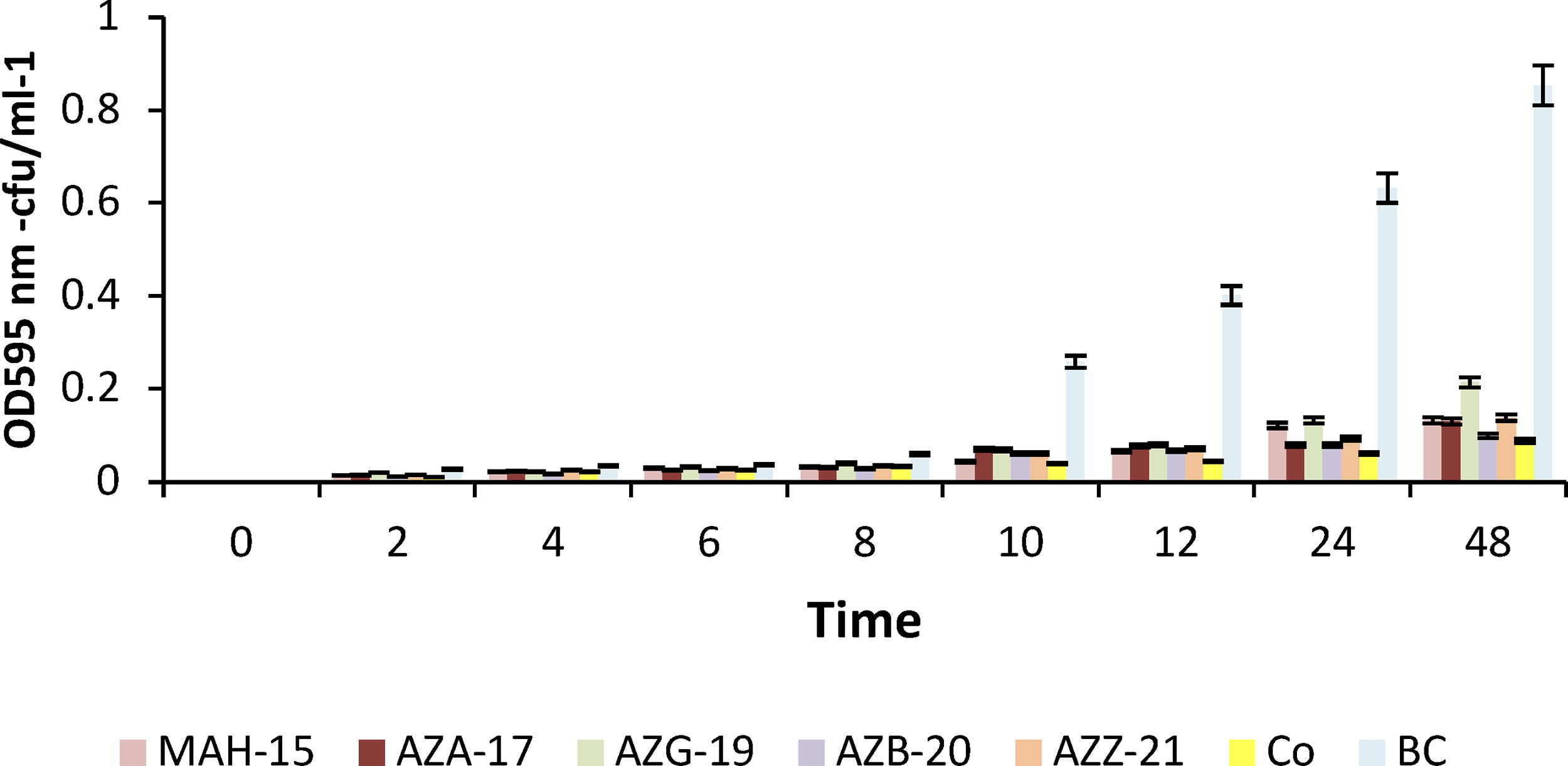
The effect of individual and cocktail phage treatments on the growth curve of Pss isolated from the soils of northwestern Iran. The phage treatments included five individual phages and a cocktail of all five phages (PC). The control group (C) received no phage treatment.

The biocontrol efficacy of different phage suspensions on cherry seedlings infected with Pseudomonas syringae pv. syringae. The phage suspensions included five individual phages and a cocktail of all five phages. The positive control group (C) received no phage treatment. The disease severity was measured by the percentage of necrotic leaves.
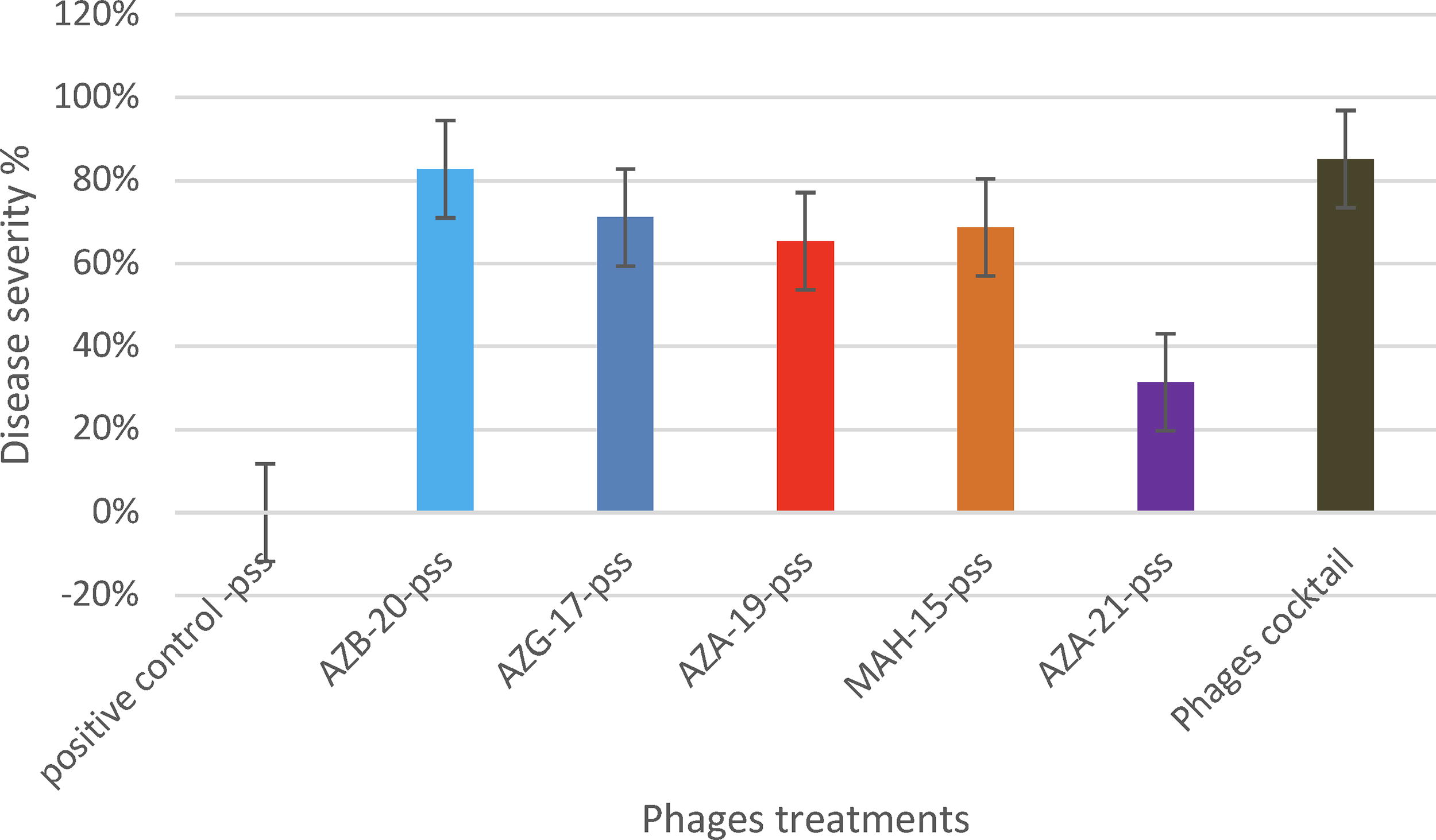
The in vivo evaluation of the biocontrol potential of five phage isolates from the soils of northwestern Iran against Pseudomonas syringae pv. syringae. The positive control group (C) received no phage treatment.
Discussion
In this research, we explored the use of five phages, specifically targeting Pss host bacteria, as potential curing agents for bacterial canker in stone fruit. Our findings pointed out that for a phage to be considered effective in biocontrol, having a broad host range for a phage is not a good character; phages with a narrow host range are more desired as they cannot affect other bacteria. With regard to the host range assay, these five isolated phages exhibited a limited spectrum. Specifically, the phages inhibited only the Pss K26 host bacteria and were uniquely tailored to Pss host bacteria, without exerting any adverse effect on other pathogenic or antagonistic bacteria. In addition, these phages elicited the formation of clear plaques, thereby suggesting a lytic infection cycle, which is notable for reducing the risk of horizontal gene transfer of lysogenic phage. 22 Using the killing curve assay, it was observed that these phages could effectively stifle the growth of the host Pss bacterial population. Consistently, higher concentrations of phage correlated with greater effectiveness in the reduction of optical density, thus suggesting that a titer of 106 Pfu/mL would result in optimal efficacy in decreasing the bacterial presence and resistance development. It should be noted, however, that high concentrations of phages can act as selection pressure on the bacterial population, thereby potentially promoting resistance evolution. 19 Consequently, the use of a phage cocktail turned out to be a more effective approach, yielding lower resistance development. 17 Although the use of individual phages did demonstrate their impact on hindering the growth of host bacteria, the evaluation of lytic dynamics curves proved to be an efficient method to gauge their killing capabilities. This methodology would assist in selecting phages with varying patterns to compose a phage cocktail, comprising an array of phages that could potentially affect maximum target bacterial strains. 23 Lastly, the effectiveness of the phage cocktail against host bacterial growth was assessed. The results underscored that the phage cocktail was more effective in restraining Pss K26 bacteria than individual phages. Notably, the growth of Pss host bacteria was rendered undetectable in NB cultures after 24 h.
It is crucial to study the stability of phages under different conditions, such as temperature, soil pH, and UV radiation. One of the most pertinent factors determining phage stability is attributed to pH, 24 as has been observed to influence the attachment, infectivity, and the intracellular replication and amplification processes of phages. 25 Extremes of high and low pH levels are disruptive to the lysozyme enzyme and capsid proteins present in phages, thus impeding their ability to attach to receptor sites on the host cell. 25 In the research undertaken, the chosen phages exhibited a similar degree of tolerance to fluctuating pH levels, corroborating the findings of preceding studies. Furthermore, temperature is a crucial variable essential to phage viability specifically in the phyllosphere, 21 an environment notably challenging for phage survival, with numerous phages reported to rapidly degrade under various adversarial conditions such as daylight hours. 21 The results from this study identify that the selected five phages displayed survival capabilities across a temperature range from 4°C up to 65°C. The maximum survival rate of phage particles was observed at −20°C, thereby indicating that freezing may be an optimal storage method for these phages. A complete depletion of phage presence was detected at temperatures of both 70°C and 80°C. Conversely, Pss host bacteria exhibit growth within the temperature bounds of 5°C and 30°C, suggesting that these five phages possess the ability to lyse Pss bacteria at relevant temperatures. 21 UV radiation has been identified as a particularly destructive environmental factor contributing to the loss of phage particles. 26 UV is likely to damage nucleic acids as well as capsid proteins. 27 This issue can potentially be mitigated by releasing the phages at high titers toward the end of the day or at night when radiation levels are at their lowest. 28 Alternative solutions could include administering the phages as a spray of concentrated phages or a cocktail of phage particles, or with carrier bacteria trapped within micro-or nanocarriers. 29
This investigation also sought to assess the efficacy of the five phages in reducing the Pss host bacterial population on a model system of cherry twigs under greenhouse conditions. The application of both individual phages and phage cocktail on the surface of the cherry twigs illustrated that all five phages were capable of reducing the Pss host population by more than 60% for individual phages and exceeding 85% in the case of the phage cocktail. It can therefore be extrapolated that using other methods such as carrier bacteria may facilitate phage replication on the phyllosphere. 30 In summation, the persistence of phages in the phyllosphere could potentially be enhanced through the use of protective formulations, carrier bacterial strains, and strategic applications of phage in the evening and just before sunrise in the early morning.4,21
In summary, the results showed that the isolated phages that have a high specificity for Pss bacteria were isolated from stone fruit trees. Thus, they have the potential for use as biocontrol agents against this pathogen. Further molecular characterization of the five phages may provide useful information for the effective control of Pss. Our results highlight the promising potential of these phages for the biocontrol of Pss contamination in stone fruit orchards.
Footnotes
Acknowledgment
We thank the Plant Bacteriology Laboratory of the University of Tabriz for providing laboratory equipment for this research.
Authors’ Contributions
Z.Z.A. designed and executed the laboratory and field experiments. S.A.M. and H.B.B. provided guidance and supervision throughout the study. D.K. performed the data analysis and interpretation. R.K. drafted, revised, and finalized the article and acted as project leader. All the authors read and approved the article for submission.
Author Confirmation
Z.Z.A., R.K., S.A.M., and H.B.B. are from the University of Tabriz (Tabriz, Iran), and D.K. is from the University of Zanjan (Zanjan, Iran), all where education and research are the primary functions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The author(s) received no financial support for the research, authorship, and/or publication of this article.