
Abstract
Introduction
S
The free-living bacterium Vibrio harveyi is widely used as a model for studying quorum sensing. These bacteria produce three types of AIs that regulate its bioluminescence: the species-specific HSL(AI-1), 6 the “universal AI” (AI-2), 7 and the Vibrio cholerae AI (CAI-1). 6,8,9 Quorum-sensing cascades regulate bacterial properties as bioluminescence, 6 motility in semi-solid medium, 10 and biofilm production. 10 –17
Lasers constitute an emerging treatment modality in medicine. Different lasers interact in a variety of ways with tissues and microorganisms. These interactions depend on different characteristics of the laser such as specific wavelengths, values of energy emitted, and exposure time. The Er:YAG laser contains a medium of Yttrium-Aluminum-Garnet crystal coded with Erbium. The wavelength of the Er:YAG laser is mid-infrared at 2.94 μm, a wavelength with a peak absorption in water molecules, 18 which are abundant in all living organisms.
Persistent bacterial infections and resistance to common antibiotic therapy have emerged as major challenges in the medical field, emphasizing the need to find new methods to address these issues. In the past decade, the introduction of different methods aimed at inhibiting bacterial cross-communication has been suggested as a feasible mean for the prevention and control of emerging bacterial infections. 19–20 Photon energy-mediated techniques may have a bacteriostatic and bactericidal effect on bacteria when coupled with a photosensitizer agent 21,22 or when applied directly to the desired target. 23 –25 However, the effect of direct low-dose laser energy on bacterial quorum sensing is a new avenue of research. This study aims to address this issue.
Materials and Methods
Laser irradiation
Laser irradiation was performed using an Er:YAG laser (Syneron, Yokneam, Israel) at a wavelength of 2940 nm equipped with a 17 mm length and 400 μ diameter sapphire tip. Different irradiation output settings were used, ranging from 0.5 W (50 mJ, 10 Hz) to 4.0 W (100 mJ, 40Hz).Total energy density did not exceed 0.255 J/cm2 for 0.5, 1.5, and 2.5 W and 0.5 J/cm2 for 4 W. Irradiation settings used were assessed as low-dose settings that would not inhibit bacterial growth. The handpiece emitting the beam was fixed at a distance of 4.5 cm from the plate surface and perpendicular to the center of the irradiated wells. Water spray was turned off to avoid dilution of samples, and each sample was irradiated once for 15 sec.
Bacterial strains and growth
V. harveyi wild type (BB120: AI-1
Irradiation of bacteria
After overnight incubation, the bacterial cultures were diluted to an optical density (OD595) of 0.5. Aliquots of 20 μL bacterial suspension were prepared in a 96-well polymerase chain reaction (PCR) plate (Sigma Aldrich, St. Louis, MO) and subjected to 15 sec of laser irradiation, and were later used as a basis for examination of postirradiation effects.
V. harveyi postirradiation growth curves and bioluminescence
Fifteen microliters from the irradiated bacterial suspension was seeded in a 96-well white plate with an optic bottom (Nunc, Denmark) and cultivated with 185 μL of thiamine and riboflavin-supplemented AB medium. Absorbance and bioluminescence of planktonic V. harveyi were recorded every 30 min for 18 h using a plate reader (GENios Tecan, Neotec Scientific Instrumentation Ltd. Camspec, Cambridge, United Kingdom) at a wavelength of 595 nm with 90 sec shaking period performed before each cycle. Relative luminescence values were calculated as the area under the curve of the ratio between luminescence and absorbance. These calculations minimized dissimilarities caused by differences in growth rates. 26,27
V. harveyi postirradiation biofilm generation
V. harveyi biofilm generation was performed as described by Soni et al. 28 Polystyrene 48-well plates (Nunc, Denmark) were seeded with 50 μL of irradiated V. harveyi cultures (OD595∼0.5) supplemented with 450 μL of thiamine and riboflavin AB medium. Incubation was performed at 30°C for 24 h, after which the biofilms were washed twice carefully using 500 μL of saline solution. The biofilm samples were then subjected to DNA quantification as described hereunder.
V. harveyi biofilm biomass quantification
DNA extraction and quantification of samples were performed as described previously. 29 In brief, 160 μL NaOH (0.05 M) (Bio Lab Ltd., Jerusalem, Israel) and 40 μL diethyl pryrocarbonate (DEPC)-treated water (Bio Basic Canada Inc, Markham, Ontario, Canada) were added to each well containing washed biofilm. After 1 h of incubation at 60°C, 18.5 μL of Tris buffer (pH = 7) (Eastman Kodak company, Rochester, NY) was added to each of the samples and stored in −20°C until further use.
DNA quantification was performed by implementing quantitative PCR with primers for 16Sr RNA using ABI prism instrument (Applied Biosystems Prism 7300, Foster City, CA). Samples were quantified according to the specific standard curve. Total genomic DNA was extracted from an overnight culture of V. harveyi BB120 using GenElute Bacterial Genomic DNA kit (Sigma Aldrich, St. Louis, MO). Extracted genomic DNA for the standard curve analysis was stored at −20°C.
V. harveyi bacterial motility assay
Motility assays were performed on soft agar plates as described previously with some modifications. 30 Cultures of V. harveyi after overnight incubation (OD595∼0.5) were irradiated at four output powers (0.5, 1.5, 2.5, and 4 W) and inoculated immediately on plates containing 0.2% agar AB medium. After an incubation period of 18 h at 30°C, motility halos were recorded and measured using Image J software (National Institute of Health, New York).
Statistical analysis
All experiments were conducted in triplicates. Descriptive statistics were calculated using a one-way analysis of variance with a two-way significance (p < 0.05) (IBM SPSS, version 21, Chicago, IL).
Results
V. harveyi postirradiation growth
Growth of planktonic bacteria was monitored by OD measurement every 30 min for 18 h. Recorded growth patterns of V. harveyi inoculums are presented in Fig. 1. Growth curves of treated bacteria demonstrated a pattern similar to the untreated controls. For all study groups, the exponential phase was evident from 3.5 up to 12 h, and the stationary phase reached an average OD of 0.74 at 18 h, for all groups with no significant differences (p > 0.05).
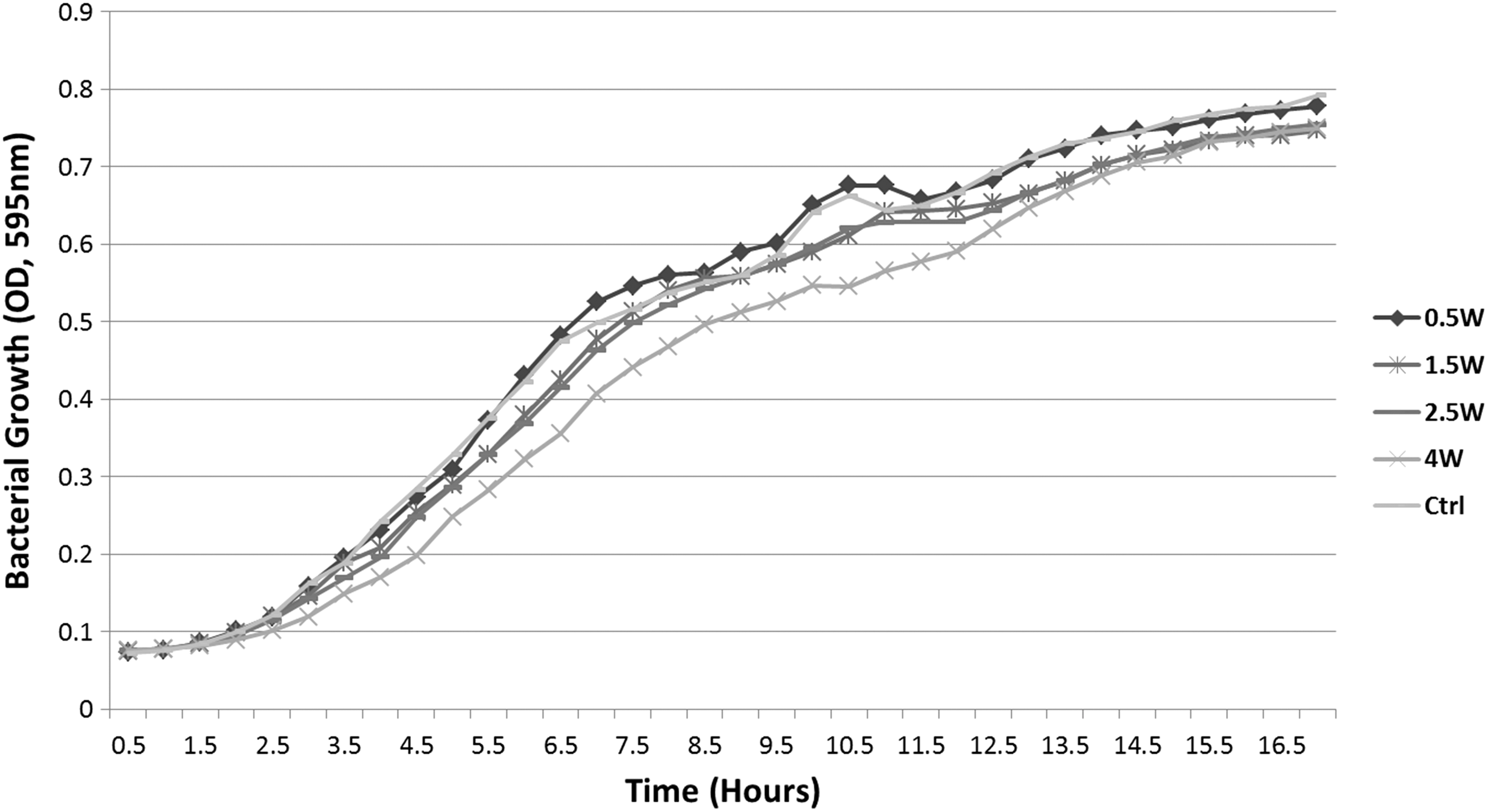
Optical density measurements of bacterial growth kinetics recorded every 30 min for 18 h. Graphs represent irradiated V. harveyi study groups (0.5, 1.5, 2.5, and 4 W).
V. harveyi biofilm biomass quantification
To determine whether growth patterns were affected in biofilm form, quantitative PCR was employed on biofilm samples grown for 24 h from irradiated samples (Fig. 2). No significant differences were observed between study groups and untreated controls, with an average of 15.07 ng/μL of bacterial DNA (p > 0.05).
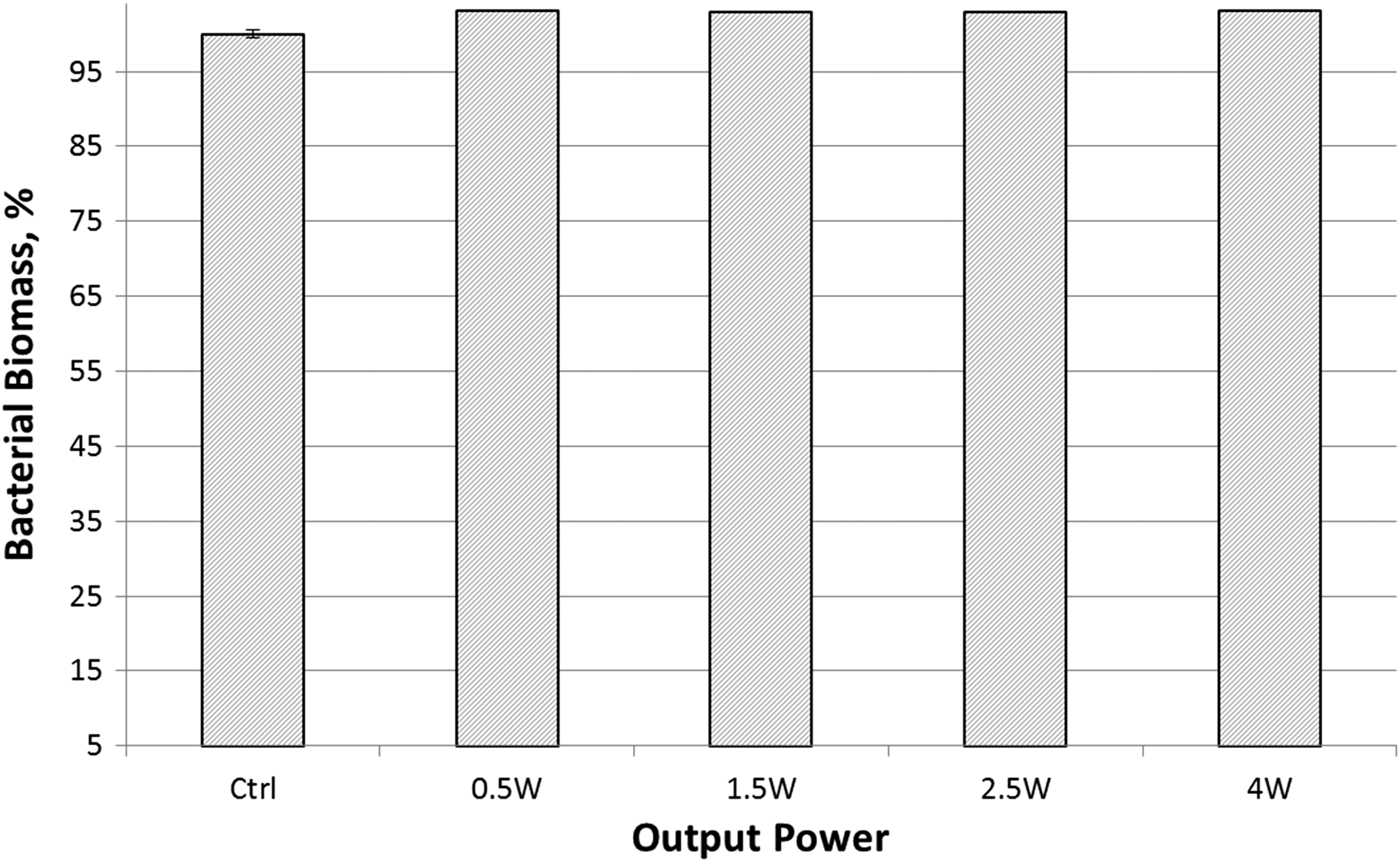
qPCR quantification of bacterial biomass from biofilms incubated for 24 h and grown from samples previously exposed to laser irradiation at 0.5, 1.5, 2.5, and 4 W compared with nonirradiated controls. qPCR, quantitative polymerase chain reaction.
V. harveyi bioluminescence and quorum sensing
Although growth curves of bacteria were not significantly affected by the output of laser irradiation (Figs. 1, 2), bioluminescence kinetics of irradiated bacteria showed significantly increased values after irradiation with higher output powers (Fig. 3). As a means of quantitatively assessing quorum sensing of incubated inoculums, the area under the curve was calculated for each group and divided by corresponding OD recordings (Fig. 4). For the 0.5 W irradiated group, a nonsignificant difference was recorded compared with untreated controls (p > 0.05), yet statistically significant differences were recorded for the 1.5, 2.5, and 4 W groups, with increased bioluminescence levels of 22.6% ± 3.31%, 3.52% ± 8.47%, and 66.9% ± 2.00%, respectively.
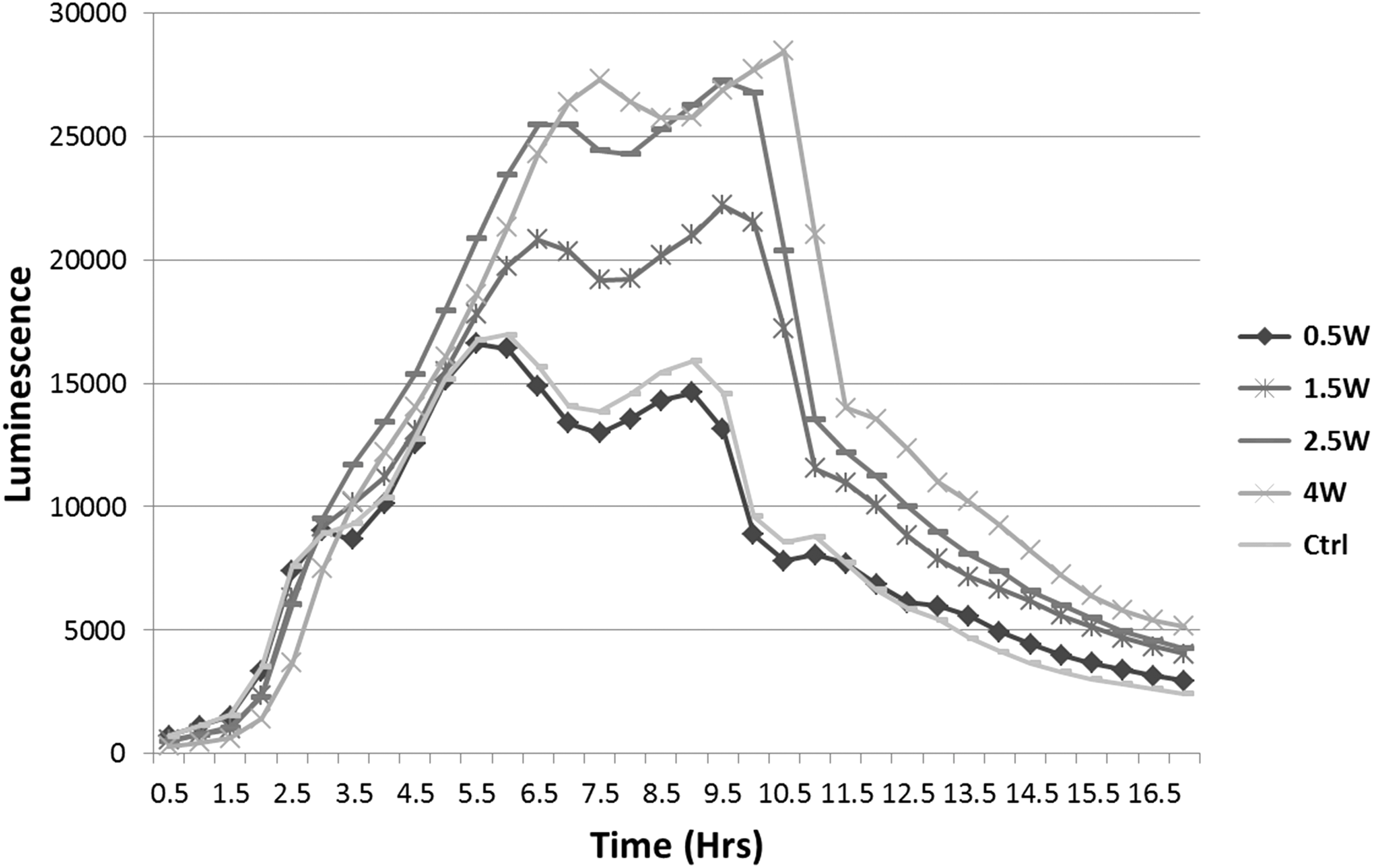
Kinetics of irradiated V. harveyi wild-type BB120 cultures during 18 h incubation. Bioluminescence values of incubated cultures increased as the output power setting was higher.
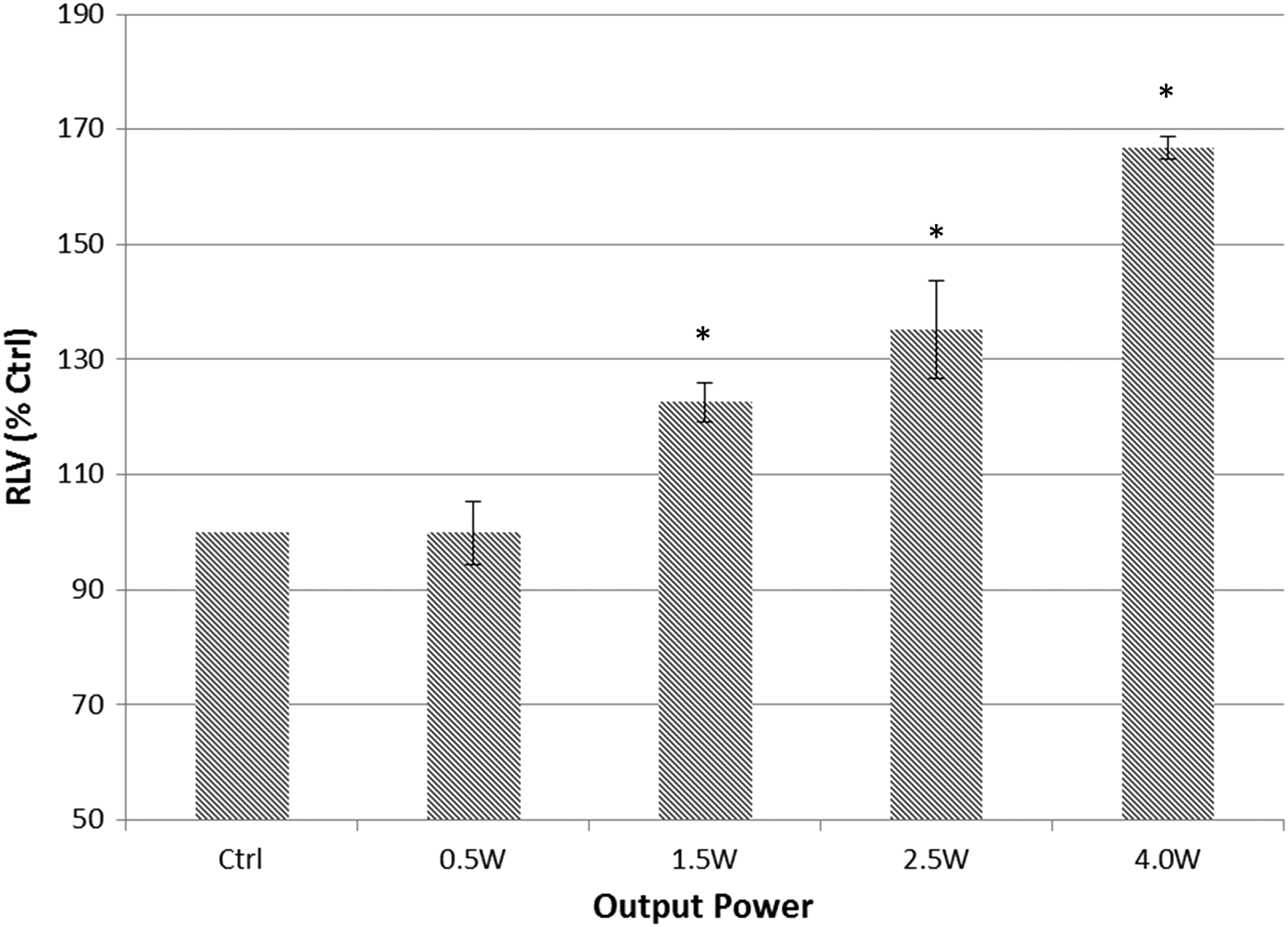
Comparison of RLVs for each group. RLV curves were calculated by dividing each bioluminescence reading by the absorbance values to normalize the luminescence value of each sample to its cell density and to avoid dissimilarities caused by differences in growth rates by absorbance values. Asterisks mark indicates significant difference to the respective controls (t-test, p value <0.05). RLVs, relative luminescence values.
V. harveyi motility assays
Modified agar plates containing AB+ medium were used to assess bacterial motility in colonies formed after irradiation. Figure 5 shows images of agar plates incubated with bacteria irradiated at different output powers. Figure 6 provides a computerized analysis of motility halos of experimental groups relative to the untreated controls. The bacteria's ability to expand colonies was significantly inhibited as the level of the laser power output was increased. Motility halos of colonies decreased by 21.7% ± 7.4% for the 0.5 W irradiated group, 37.5% ± 8.0% for the 1.5 W group, 24.4% ± 8.0% for the 2.5 W group, and by 65.9% ± 8.4% for the 4.0 W group.
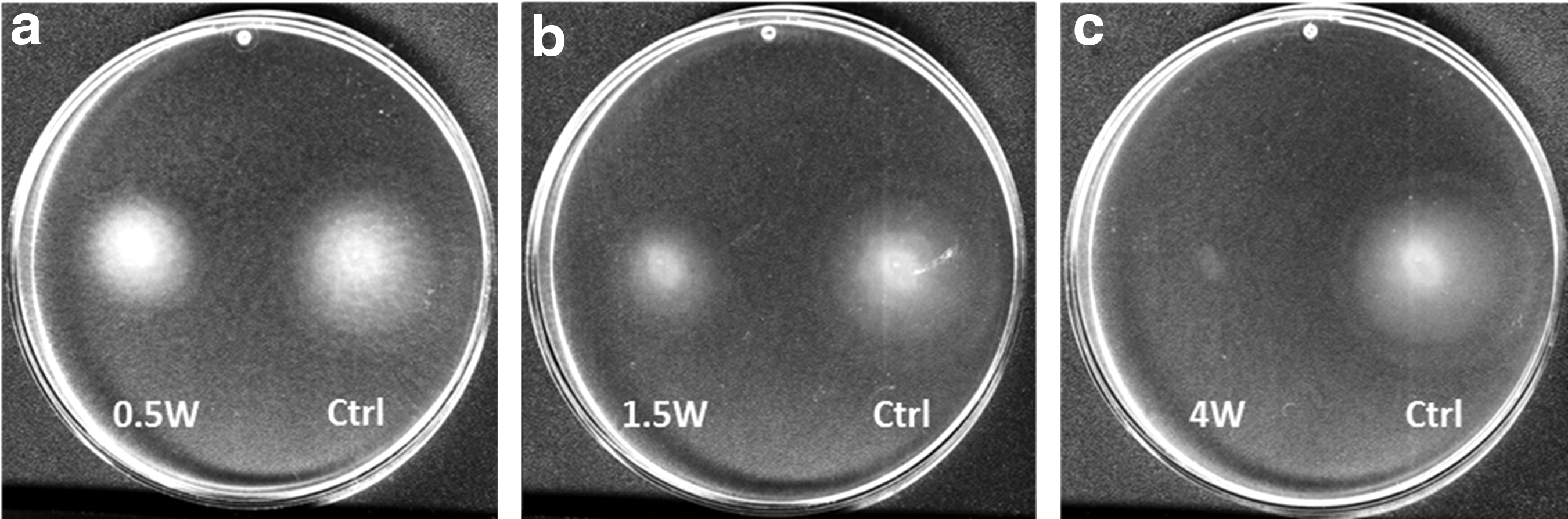
Scans of agar plates inoculated with bacteria after laser irradiation at 0.5 W
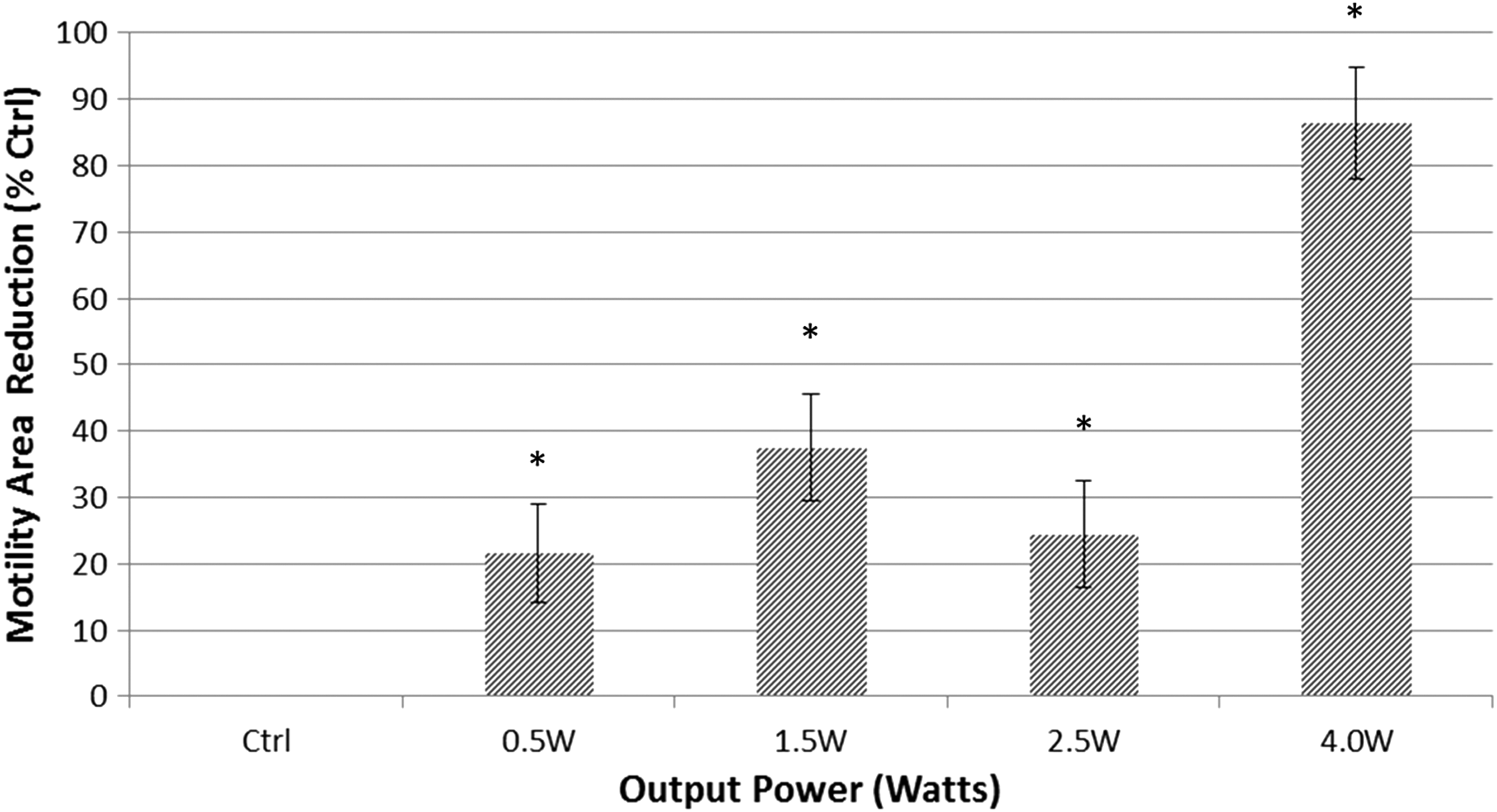
Comparison of motility halos reduction by irradiated bacteria for 15 sec at 0.5, 1.5, 2.5, and 4.0 W to untreated controls. Halos area analysis was performed using ImageJ software. Asterisks mark represents significant difference to the respective controls (t-test, p value <0.05).
Discussion
Synthesis, secretion, and binding of extracellular signal molecules, such as AIs facilitate bacterial cross-communication also known as quorum sensing. 1 The bacterial response to the AIs leads to alterations in bacterial gene regulation, which leads to changes in phenotypic properties. Inhibition of quorum sensing is important for controlling bacterial properties. Current research on inhibition of bacterial quorum sensing focuses mainly on finding sensor blockers, AIs analogues, and affecting secretion and degradation of AIs. 19,31,32 Since the quorum-sensing cascade is an important regulatory system in bacteria, additional means for preventing this cross-talk should be explored. The effect of photon-based energy irradiation on quorum sensing has yet to be investigated. Photon energy, at specific wavelengths, can effectively penetrate into deep layers of bacterial biofilm. Harnessing this energy may offer a substantial advantage when compared with standard techniques. Therefore, we sought to investigate the effect of low-dose Er:YAG laser energy on V. harveyi, which is widely used as a model for bacterial cross-communication.
Photon-based energy has been shown to affect bacteria in different ways. 33,34 Blue light has been shown to have a sustained postirradiation effect on Streptococcus mutans when grown into new biofilm, with increased growth after irradiation on the one hand, and impaired viability and virulence factors on the other. 35 Confocal laser scanning microscopy of S. mutans biofilms grown on tooth surfaces and subjected to high power CO2 irradiation demonstrated an increased proportion of dead bacteria at the margins of the irradiated area, whereas at the center of the focal point the bacteria evaporated. 36
Low-level laser therapy may affect bacterial characteristics with a minimal effect on growth and viability 37,38 or inhibit bacterial viability at higher doses. 39 These effects also depend on the environmental conditions and physiological state of the bacteria. 37,38 In addition, the Er:YAG laser has been shown to have therapeutic effects at low-energy levels. 39,40
When using low output settings for up to 15 sec with a power density of no more than 1.0 J/cm2, growth of irradiated inoculums did not substantially differ from untreated controls in planktonic state, reaching an OD of at least 0.73 at the stationary phase. This ensured that the changes we found in the bacterial characteristics after laser treatments were not because of differences in growth patterns of the irradiated bacteria.
Bioluminescence of growing bacterial batches was recorded in conjugation with OD measurements. Bioluminescence of the bacteria has been used as a means of measuring the secretion and distribution of AIs into the bacterial environment and as an indicator of cell-to-cell communication, especially when using V. harveyi as the tested bacteria. 41,42
A positive dose response correlation was found between the laser energy levels and the measured increase in quorum sensing, which ranged from 22.6% to 66.9% for irradiated groups (at 1.5, 2.5, and 4 W). Different stress levels induced by photon energy may cause bacteria to increase intercellular quorum sensing, as means of establishing community-coordinated protection against the laser irradiation. Although population-based AI secretion was increased, cellular mass in biofilm form was not affected by the low-dose irradiation. These findings suggest that although the cascades in charge of intercellular secretion and binding were upregulated, other intracellular processes were inhibited to the point that the cellular mass in the biofilm was similar after 24 h of incubation.
V. harveyi is known to produce biofilms at high population density. Biofilm production is regulated through genes regulated by quorum sensing. 11,14,16,17 When biofilm grown from irradiated samples was analyzed, only negligible differences were recorded between experimental groups and untreated controls, measured as DNA levels that indicate cellular mass. 43 Subjecting bacteria to very low levels of laser energy may have negligible and even a positive effect on bacterial growth through biostimulation, 38,39,44 and not necessarily a bactericidal effect.
The three main quorum-sensing cascades that V. harveyi utilizes contribute to biofilm gene regulation and expression. 8,45 All three signal cascades eventually rely on dephosphorization of a regulatory component—the LuxO protein. Once in its phosphatase form, LuxO is unable to transcribe regulatory RNAs that inhibit production of the master regulator protein LuxR, thus activating gene expression. 46 Waters and Bassler 41 showed that although all three signal cascades eventually lead to LuxR production and gene expression, different promoters have different affinities for the regulatory protein. Each cascade (HAI-1, AI-2, and CAI-1) has a different effect on LuxR accumulation, meaning that the effect of each of the three signals on gene activation is different. In the current research, the Er:YAG laser irradiation may have affected these cascades in different ways, either by inhibiting a “weak” cascade (CAI-1) that may have a minimal effect on bacterial growth or by inhibiting a “strong” cascade (HAI-1) and in the same time upregulating other cascades. The data presented here emphasize that irradiated bacteria were still able to grow and develop populated biofilms.
Motility is considered as one of V. harveyi primary virulence factors. The bacterium has a dual flagellar system combining a single polar flagellum for swimming in aqueous environment and a set of lateral flagella utilized for swarming on more viscous surfaces. 47 A general reduction in swimming across all irradiated colonies was recorded, from a 21.7% reduction for 0.5 W irradiation and up to 65.9% for 4.0 W irradiation. Motility and biofilm production are positively regulated by quorum sensing and by more than one signaling pathway, including the AI-2 and the HAI-1 signaling cascades. 30,48 As stated before, the effect the laser had on bacterial metabolism may have hampered the communication signals that possess different transduction capability, leading to the different phenotypes observed in our results. 41 For that reason, it is possible that although several components affected by laser irradiation could have a crucial role in expression of flagellar genes, it may have a limited effect on the growth of bacteria in biofilm and planktonic states, causing the different phenotypes observed in our results.
In light of current evidence, we suggest that the Er:YAG laser had a direct effect on behavioral adaptation of V. harveyi. Further research should focus on analyzing gene expression and biofilm characteristics of the irradiated bacteria. The findings of our research may add a crucial insight onto the field of manipulating bacterial communication, with notable relevance for control of infectious diseases using laser irradiation.
Footnotes
Acknowledgments
This work was part of Dr. Idan Redenski's M.Sc. thesis. This study was supported, in part, by the Cabakoff foundation.
Author Disclosure Statement
No competing financial interests exist.