
Abstract
Introduction
L
Different wavelengths have been tested and showed different effects. 17 –19 The blue light (wavelength 400–500 nm) has been reported to treat several diseases, principally for hyperproliferative skin disorders such as acne, 1 –3 psoriasis, 5 and keratosis. 20 LED at 415 ± 15 nm can contribute to the treatment of keloids and other fibrotic skin diseases because it inhibits dermal fibroblast proliferation. 21 Moreover, LED at 453 nm modulates differentiation and proliferation of human skin cells. 22
Once the effect of LED on the skin was demonstrated, it is hard to believe that it has no effect on dendritic cells (DCs) since the skin is one of its major sites of work. DCs play an essential role in developing the immunity because they consist of a group of highly specialized white blood cells that capture, process, and present antigens. 23,24 Further, DCs have been linked to several skin pathologies, such as autoimmune diseases and tolerance process. 25 –28
In summary, despite this probable involvement, previous data showing the effects of LED on DCs, important cells of the innate response, are still incipient. Thus, the aim of this study was to evaluate the effects of LED at 460 ± 20 nm on murine DCs.
Methods
Animals
Wild-type mice, C57BL/6, between 6 and 8 weeks old were used. The animals were bred and maintained, as determined by the Ethics Committee on Animal Use (CEUA), in experimental animal facilities of the Federal University of Triângulo Mineiro/UFTM, Uberaba-MG, Brazil.
Reagents
Ultrapure lipopolysaccharide (LPS) from Escherichia coli 0111: B4 was acquired from Invivogen (San Diego, CA). Recombinant murine granulocyte macrophage colony-stimulating factor (GM-CSF) was acquired from PeproTech (Rocky Hill, NJ). To determine the doses of both molecules, the recommendations of the manufacturer were followed. Also, concentration–response studies were performed. Antibodies and cytokine kits (OptEIA™ ELISA) were acquired from BD Biosciences (San Jose, CA) and/or eBioscience (San Diego, CA).
Bone marrow dendritic cells
Bone marrow-derived dendritic cells (BMDCs) were obtained, as described previously by Oliveira et al.,
29
with some modifications. Briefly, mice tibia and femurs were used to get bone marrow cells. These cells were prepared at 2.0 × 105 cells/mL and cultured in 10 mL RPMI-1640 (GE Healthcare, Uppsala, Sweden) supplemented with 10% v/v inactivated fetal bovine serum (Gibco, Grand Island, NY), 25 mM
LED irradiation on BMDCs
LED device (Radii-cal—SDI Limited, Victoria, Australia), emitting 460 ± 20 nm wavelength with an irradiance of 275 mW/cm2 was used (Table 1). The intensity of light emitted by the apparatus was evaluated by a radiometer Demetron (Kerr/Sybron Dental). For this, the active tip of the photopolymerizer was placed vertically on the central part of the photosensitive surface of the radiometer at a mean distance of 5 mm. Three measurements were taken to obtain the mean of intensity of light. To evaluate the influence of LED on DCs, BMDCs at a concentration of 2 × 105 cells per well (48-well cell culture plates) in a volume of 200 μL of Roswell Park Memorial Institute medium (RPMI) 10% were stimulated with different exposure times (12, 30, or 60 sec), giving a fluence of 3.3, 8.2, or 16.5 J/cm2, respectively. After 1 h, 200 μL of RPMI 10% with or without LPS (final concentration 100 ng/mL) was added. Cells without irradiation, which were cultured in the same medium, with or without LPS, were prepared for the control. After 18 h, the molecules MHC-II, CD40, and CD86 were analyzed by flow cytometry. In addition, the levels of cytokines IL-12p40, IL-6, IL-10, and TNF-α, in the supernatants, were measured by ELISA.
Flow cytometry analysis
The cultured cells were analyzed according to Oliveira et al. 29 by flow cytometry. The monoclonal antibodies used in this step were as follows: anti-CD11b, anti-CD11c, anti-CD86, anti-MHC-II, or anti-CD40 labeled with PECy7, FITC, antigen-presenting cell (APC), or PE. FACSCalibur flow cytometer (BD Immunocytometry Systems) was used for acquisition and the FlowJo software (Tree Star, Inc., Ashland, OR) and CellQuest 5.1 software (BD Biosciences) for analyses.
Cytokine assays
The supernatant level of IL-10, IL-12p40, IL-6, and TNF-α was measured by specific commercial kits, BD OptEIA (BD Biosciences), by ELISA-type immunoassay “sandwich.” To measure the cytokines IL-12p40 and IL-6, the samples were diluted 20 and 10 times, respectively. Results were expressed in pg/mL and analyzed by linear regression with the aid of StatView program.
Statistical analysis
GraphPad Prism 5.0 (GraphPad Software, San Diego, CA) was used to perform the statistical analysis of the experimental data. All variables were tested for normal distribution (Kolmogorov–Smirnov test) and variance (Bartlett's test). Then, analysis of varience (ANOVA) with Tukey's post-test was used. The results are expressed as mean and standard deviation. A p value less than 0.05 was considered significant.
Results
Effect of LED on cytokine production
LED increases (p < 0.05) IL-12p40 and IL-6 production in both BMDCs (Fig. 1) and LPS-matured BMDCs (Fig. 2). Figure 1A shows the increase of IL-12p40 on BMDCs, which was approximately three times when irradiated with 3.3 and 8.2 J/cm2 compared with the BMDCs without LED. In Fig. 2A, the increase of IL-12p40 on LPS-stimulated BMDCs was two times more when irradiated with 3.3, 8.2, and 16.5 J/cm2 compared with LPS-stimulated BMDCs without LED. Regarding the cytokine production of IL-6 on BMDCs, as shown in Fig. 1C, the increase was approximately five times with 3.3 J/cm2, six times with 8.2 J/cm2, and four times with 16.5 J/cm2 compared with the BMDCs without LED. On LPS-matured BMDCs, the IL-6 production increased approximately three times, with all the fluences used (Fig. 2C). In all cases, the increase was not time dependent (Figs. 1 and 2). The levels of the cytokines TNF-α (Figs. 1B and 2B) and IL-10 (Figs. 1D and 2D) were unchanged in any of the experiments done.
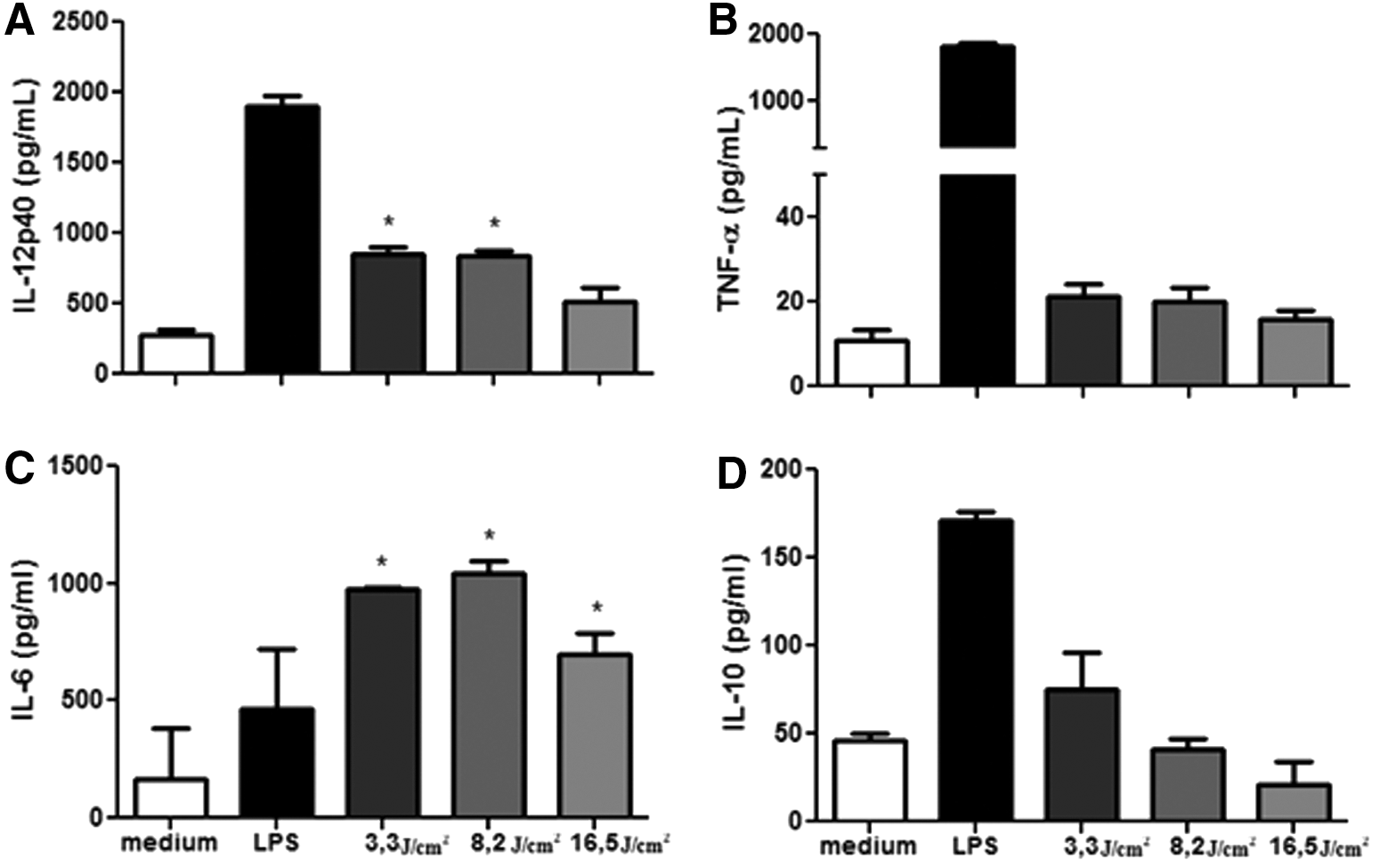
Effect of LED on cytokine production of BMDCs. Bone marrow cells derived from C57BL/6 mice were collected and cultured for 7 days in the presence of GM-CSF to allow differentiation into BMDCs. BMDCs were then treated with fluencies of 3.3, 8.2, or 16.5 J/cm2. After 18 h, the culture supernatant was collected and analyzed to detect IL-12 p40
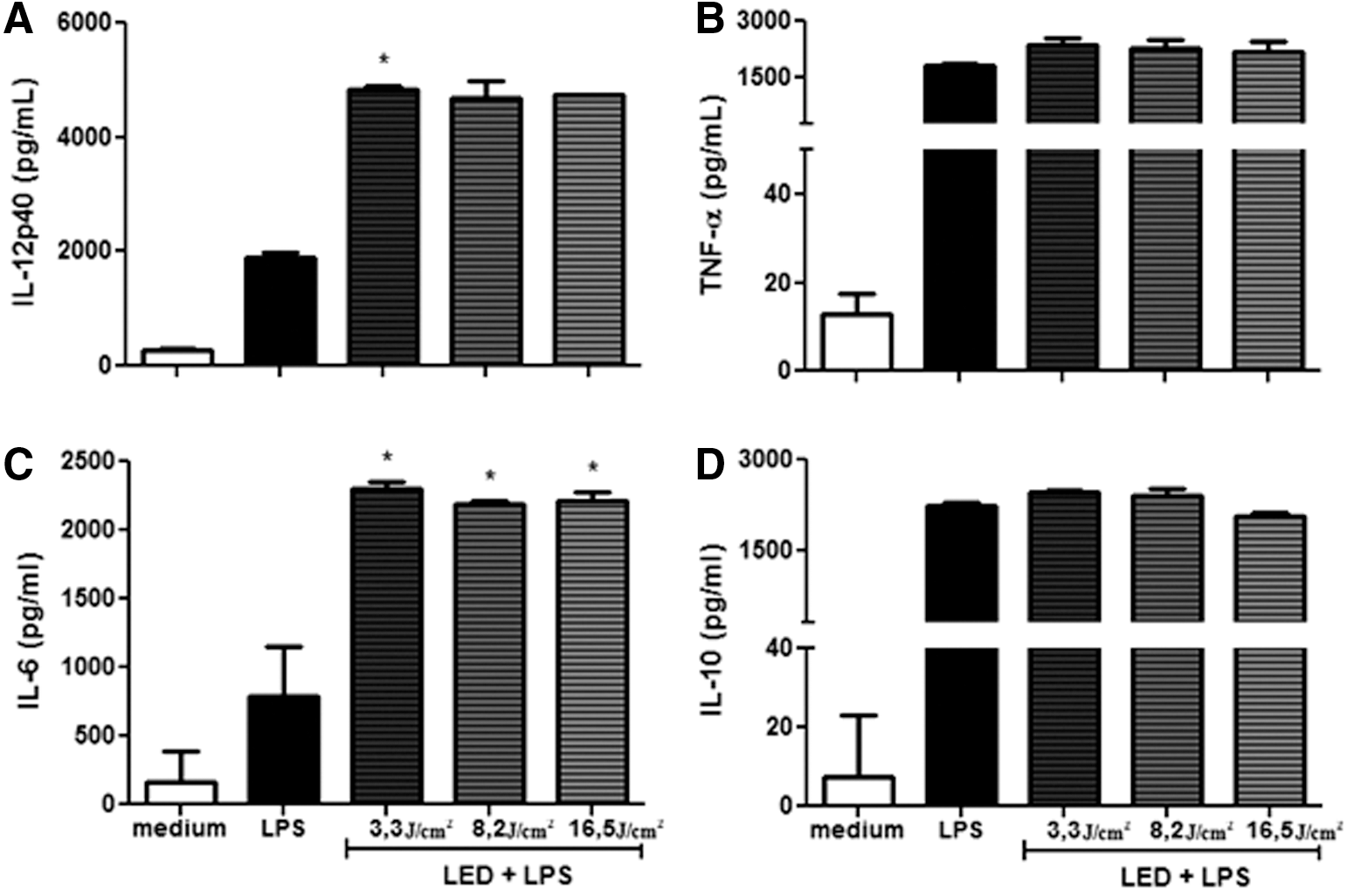
Effect of LED on cytokine production on BMDCs stimulated with LPS. Bone marrow cells derived from C57BL/6 mice were collected and cultured for 7 days in the presence of GM-CSF to allow differentiation into BMDCs. BMDCs were then treated with fluencies of 3.3, 8.2, or 16.5 J/cm2. One hour later, LPS (100 ng/mL) was added. After 18 h, the culture supernatant was collected and analyzed to detect IL-12 p40
Effect of LED on surface molecules
The expression of MHC-II molecule was significantly inhibited (p < 0.05) on BMDCs, as shown in Fig. 3A and B. The inhibition was ∼28% with 3.3 J/cm2 and 14% with 8.2 J/cm2, compared with the BMDCs without LED. Besides that, the expression of the CD86 molecule was significantly increased (p < 0.05) on BMDCs (Fig. 3C, D), compared with the BMDCs without LED, but not in LPS-matured BMDCs (Fig. 4C, D). In other words, only the BMDCs without LPS had its surface molecules altered by LED (Fig. 3A/B and C/D). The increase of CD86 molecule was about 20% on all three different fluences used (Fig. 3C, D). The expression of CD40 was not altered in any of the experiments (Figs. 3E/F and 4E/F).
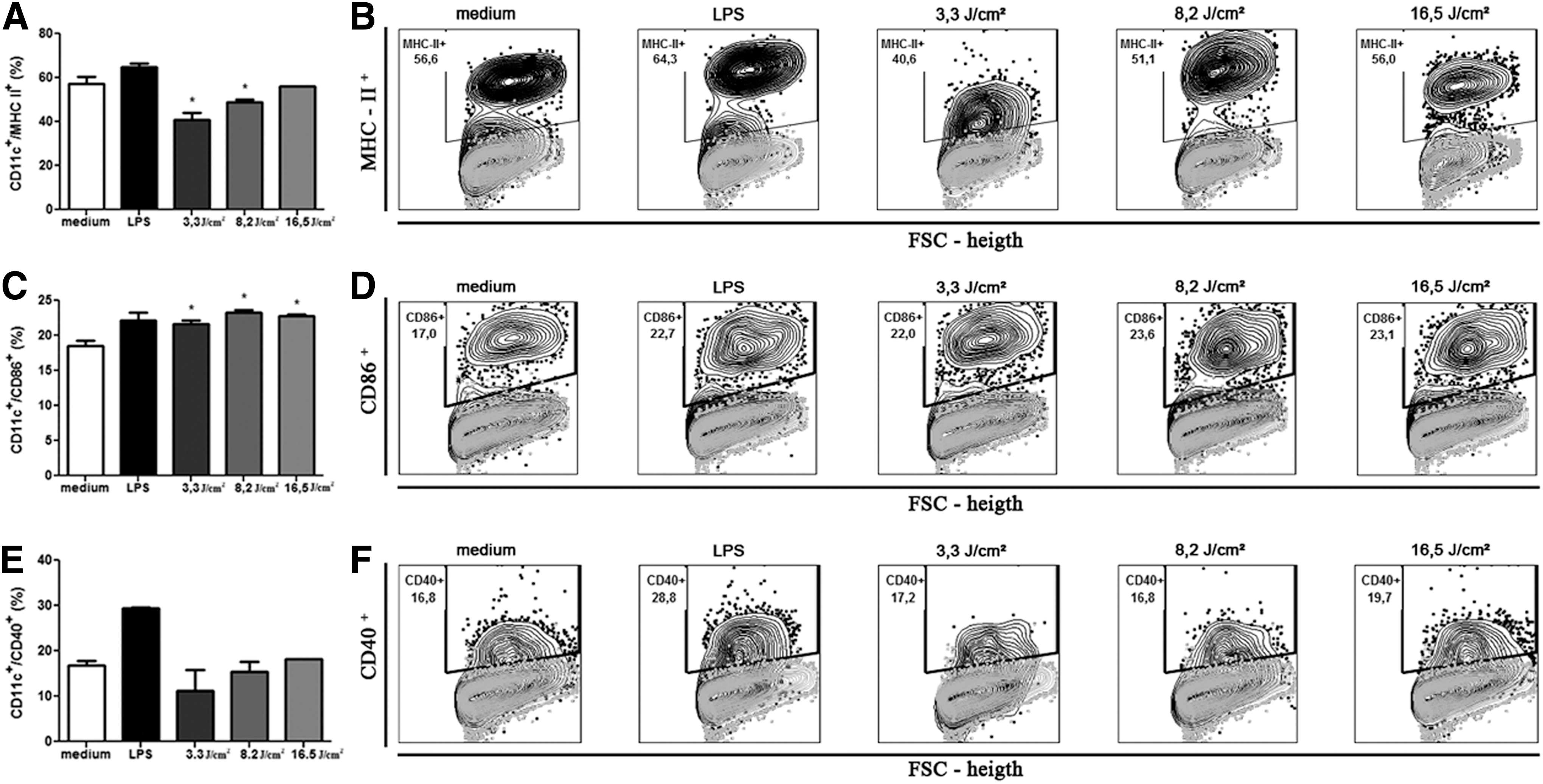
Effect of LED on the expression of MHC II, CD40, and CD86 on BMDCs. Bone marrow cells derived from C57BL/6 mice were collected and cultured for 7 days in the presence of GM-CSF to allow differentiation into BMDCs. BMDCs were then treated with fluencies of 3.3, 8.2, or 16.5 J/cm2. After 18-h, culture cells were collected and evaluated for the expression of surface molecules CD11c+/MHC II+
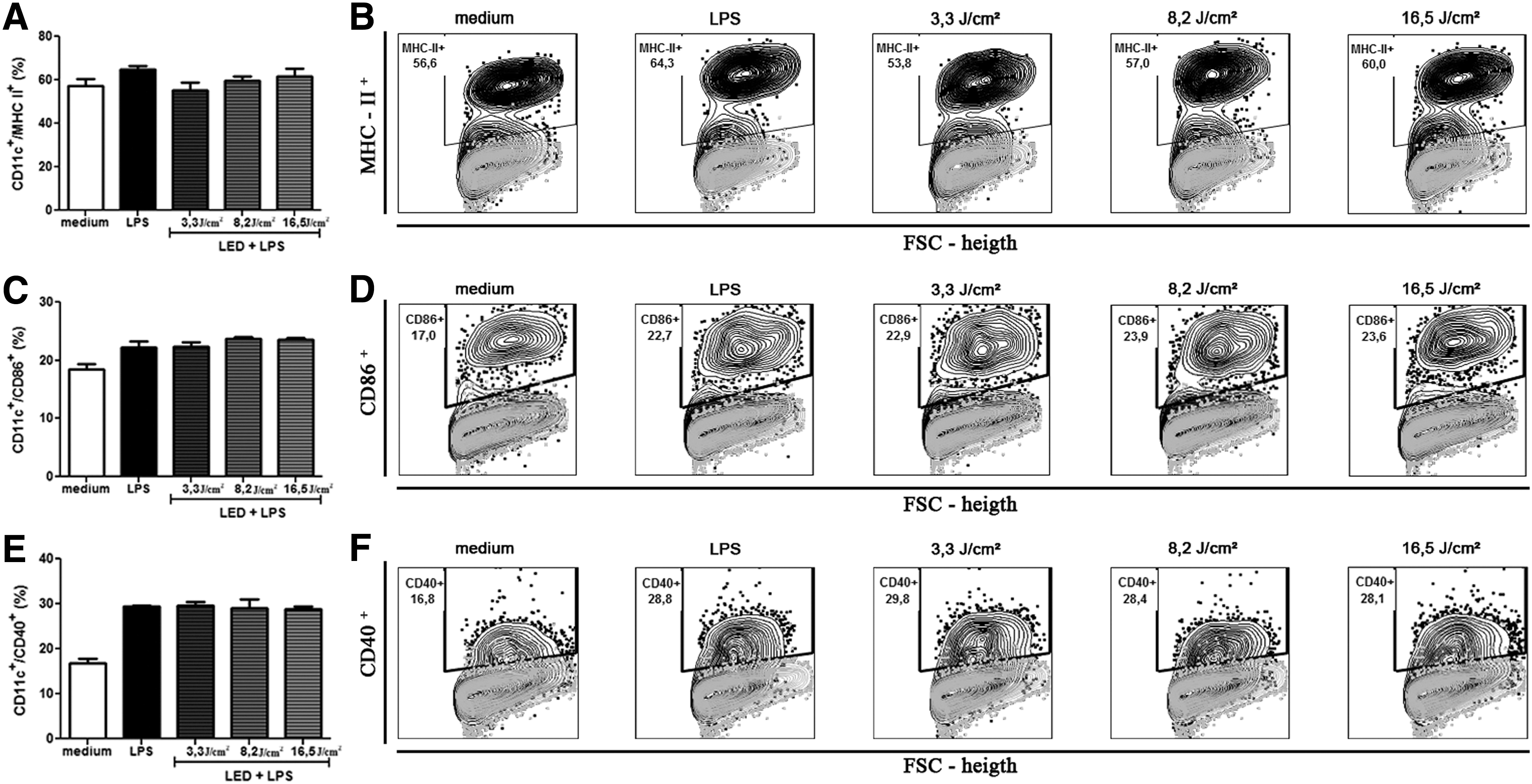
Effect of LED on the expression of MHC II, CD40, and CD86 on BMDCs stimulated with LPS. Bone marrow cells derived from C57BL/6 mice were collected and cultured for 7 days in the presence of GM-CSF to allow differentiation into BMDCs. BMDCs were then treated with fluencies of 3.3, 8.2, or 16.5 J/cm2. One hour later, LPS (100 ng/mL) was added. After 18 h, culture cells were collected and evaluated for the expression of surface molecules CD11c+/MHC II+
Discussion
The present study demonstrated that LED at 460 ± 20 nm modulates immature and mature DCs. These effects are interesting because we, for the first time, reveal that LED modulates the biology of these cells. These results improve the knowledge about the LED effect, since so far most of the studies only demonstrated the anti-inflammatory effects of LED. 30 –32 In our view, the findings shown herein indicate that an increase in the levels of inflammatory cytokines and a higher expression of costimulatory molecules on DCs would be important for the generation of a stronger and more efficient immune response.
DCs are the most important APCs of the lymphoid and nonlymphoid tissues. In other words, they are cells extensively distributed, in constant migration, and being found in nonlymphoid peripheral tissues, such as skin, allowing exercising their supervisory role. 28,33 After activation, occurs the maturation of these cells, a process that leads to the production of many cytokines such as IL-6, IL-12, and TNF-α and increases expression of costimulatory molecules, such as MHCII, CD86, CD80, and CD40. 34
In this study, we showed that LED at 460 ± 20 nm, in both immature and mature cells, activated by the presence of LPS, was able to stimulate the secretion of IL-12 and IL-6, cytokines widely known for their proinflammatory activities. It is reported that ultraviolet light (300–400 nm) might increase the expression of some inflammatory cytokines, 35 –37 but with LED at longer wavelengths (600–850 nm), anti-inflammatory effects have been showed. 30 –32 Therefore, depending on the wavelength of light, immune effects can be different. Anti-inflammatory effects were observed when a bigger wavelength of light was used for long or frequent periods of time. 38
At the 392.5 ± 1 nm wavelength, the use of LED increased the expression of TNF-α and IL-1. 18 Moreover, LED at 710 nm slightly induces the transcription of IL-1β and IL-6. Besides that, the population of murine CD4+ T cells increases. 30 These results suggest that this proinflammatory effect can increase DC maturation. Thus, the matured DC will be able to present the antigens with more efficiency. 39 Therefore, the wavelength of light, frequency, and exposure time will be decisive for the direction of results. 19,30
Once the process of maturation starts, DCs migrate to the peripheral lymphoid tissues where they presents antigens to naive CD4+ T lymphocytes, resulting in linking between innate and adaptive immunity. 40 In the initial stages of Th differentiation, the cytokines secreted by DCs play a major role, with the cytokine IL-12 being one the most important for the generation of Th1 cells. 41
The costimulatory molecules that increased on DCs after activation are central for the activation of T lymphocytes, and in the absence of costimulatory signals required for this activation, the process is impaired and the T cells may become anergic. 42 Here we showed that on immature BMDCs, the LED is able to downmodulate the cellular expression of MHC-II, increases CD86, and does not alter CD40, but in LPS-matured BMDCs no effect was observed. Thus, the LED would not affect the antigenic presentation by already activated DCs. There are no works associating LED and these markers in the literature.
Conclusions
In conclusion, the results demonstrate that LED stimulates cytokine production and expression of costimulatory molecules on BMDCs, suggesting a proinflammatory role in the tested conditions, which probably increased DC maturation, turning them into more efficient antigen-presenting cells. In summary, these results may help to improve knowledge about the clinical application of LED and its relevance to the treatment of several diseases.
Footnotes
Acknowledgments
The Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), and PAPE-UNIUBE supported this work.
Authors' Contribution
T.M.C.-C. contributed to the experimental procedure, planning the study, statistical analysis, and editing the article; M.T.M. contributed to the experimental procedure, planning the study, and editing the article; M.V.d.S. participated in the design of the study, cytometric analysis, design of figures, and editing the article; G.C.M.B.T. participated in the standardization of the study and editing the article; V.R. took part in the design of the study and editing the article; C.J.F.O. conceived the study, participated in its design and coordination, and editing the article; G.T. conceived the study, participated in its design and coordination, and editing the article.
Author Disclosure Statement
No competing financial interests exist.