
Abstract
Prion diseases are fatal neurodegenerative diseases that occur either spontaneously or genetically or are caused by infection. Spontaneously occuring prion diseases are age related. The infectious agents, called prions, are proteinaceous infectious particles, composed mainly of the host-encoded prion protein (PrP) in a misfolded, insoluble, and aggregated isoform. Advanced glycation end products (AGEs) are well known to contribute to protein misfolding, insolubility, and aggregation. Thus, we studied if AGE-modification could influence PrP aggregation. We analyzed PrP preparations immunochemically to determine if they contain AGE-modified PrP. We also studied the influence of AGE modifications on the PrP aggregation process in vitro.
Introduction
Advanced glycation of proteins is a nonenzymatic reaction cascade between reducing sugars and reactive amino groups of proteins. This reaction, called the Maillard reaction, occurs in vivo in an age-related manner. The Maillard reaction leads to several modifications in proteins such as adducts or crosslinks, which are called advanced glycation end products (AGEs). 5 AGEs accumulate in long existing proteins within the lifespan of an organism. Furthermore, they are suspected to play an important role in several neurodegenerative diseases like Alzheimer or Parkinson disease. 6,7 At the molecular level, AGE modifications have been shown to induce protein insolubility, higher resistance against proteolytic digestion, and cytotoxicity. 5,8,9 Recently, they have been demonstrated to induce formation of amyloid fibrils. 10
We investigated whether the formation of AGEs could contribute to the misfolding and aggregation of the prion protein. In a first step, we analyzed if infectious prions or noninfectious PrPC preparations contain AGE modifications. Additionally, we applied a method to induce formation of AGEs in vitro to PrP and examined if AGE modification has an effect on PrP amyloid formation.
Materials and Methods
Prion protein preparations
Infective PrP27–30 was prepared from infected Syrian hamster brains according to Prusiner et al. 11 PrPC from healthy Syrian hamster brains was prepared according to Pan et al. 12 CHO-PrPC was prepared from Chinese hamster ovary cell lines as described in Elfrink et al. 13 Recombinant Syrian hamster PrP comprising the full-length recPrP(29–231) and amino-terminally truncated recPrP(90–231) were prepared as described in Elfrink et al. 13
In vitro sodium dodecyl sulfate conversion system
RecPrP was subjected to a sodium dodecyl sulfate (SDS)-dependent in vitro conversion assay, as reported previously. 14 All experiments were performed in 10 mM NaPi, 250 mM NaCl, 0.01% SDS, pH 7.2 (conversion buffer), at 37°C. RecPrP concentration concentrations are indicated in results.
In vitro-generated AGE modification
AGE modifications were achieved according to Thornalley et al. 15 by incubating 125 ng/μL of PrP in 1 mM glyoxal, 1 mM methylglyoxal, and 200 mM NaPi for 2 days at 45°C. Samples were directly analyzed for AGE-specific autofluorescence (λex = 370 nm, λem = 440 nm). Additionally, AGE fluorescence was measured with the fluorescence parameters: λex = 445 nm, λex = 460–650 nm in a Tecan Sapphire plate reader (Tecan Group, Maennedorf, Switzerland).
Immunodetection of PrP and AGE-modified PrP
After boiling in loading buffer, samples were applied to a dot blot and immunostained using either PrP-specific antibodies (pAb A416 or 3F417) or AGE-specific antibodies (PEN-12 = anti-Pentodisin and H12 = anti-Pyrralin; GERBU Biotechnik GmbH, Gaiberg, Germany; 4G9 = anti- N-carboxymethyllysin [CML] 18 ). Immunostaining of blotted samples was performed according to Appel et al. 2 Rotiblock (Roth, Karlsruhe, Germany) was used as blocking reagent.
To examine solubility of recPrP after incubation in conversion buffer, samples were centrifuged at 100,000 × g for 1 h. Supernatant (s) and pellet (p) fractions were applied to a dot blot and immunostained using a PrP-specific antibody (pAbA4).
Thioflavin T assay
ThioflavinT assays using 8 ng/μL recPrP(29–231) were carried out as described in Panza et al. 19 Spectra were recorded from 460 nm to 650 nm with fixed λex of 455 nm.
Polyacrylamide gel electrophoresis and silver staining
SDS/polyacrylamide gel electrophoresis (PAGE) (12%) and silver staining were carried out as described in Birkmann et al. 20
Results
To investigate if preparations of PrP contain AGE-modified PrP, we used AGE-specific antibodies for immunochemical detection. We analyzed four preparations of noninfective PrP: recPrP(29–231) and recPrP(90–231), both lacking posttranslational modifications; CHO-PrPC and natural PrPC prepared from healthy hamster brain, both containing posttranslational modifications (glycosylations and glycosylphosphatidylinositol [GPI] anchor). These PrP preparations were compared to highly infectious PrP27–30 prepared from brain tissue of hamster afflicted with scrapie (Fig. 1A). In vitro AGE-modified bovine serum albumin (BSA) and keyhole limpet hemocyanin (KLH) were used as AGE-positive controls. The upper blot of Fig. 1A shows PrP-positive signaling as derived by anti-PrP antibody 3F4. As expected, both non-PrP controls (Fig. 1F, G) did not show any signal, whereas all PrP containing samples (Fig. 1A–E) showed PrP-specific signals. AGE-positive signals were derived by three different anti-AGE antibodies: CML was detected by 4G9, pentosidine by PEN-12, and pyrralin by H-12. CML signaling was observed in all samples of different strength. The lowest level of CML signaling was detected in PrP27–30. Pentosidin and pyrralin were detectable mainly in recPrP(90–231) samples. All results were reproduced by silver-stained PAGE followed by western blot analysis, proving that AGE- and PrP-positive signaling occurred in the same bands (data not shown).
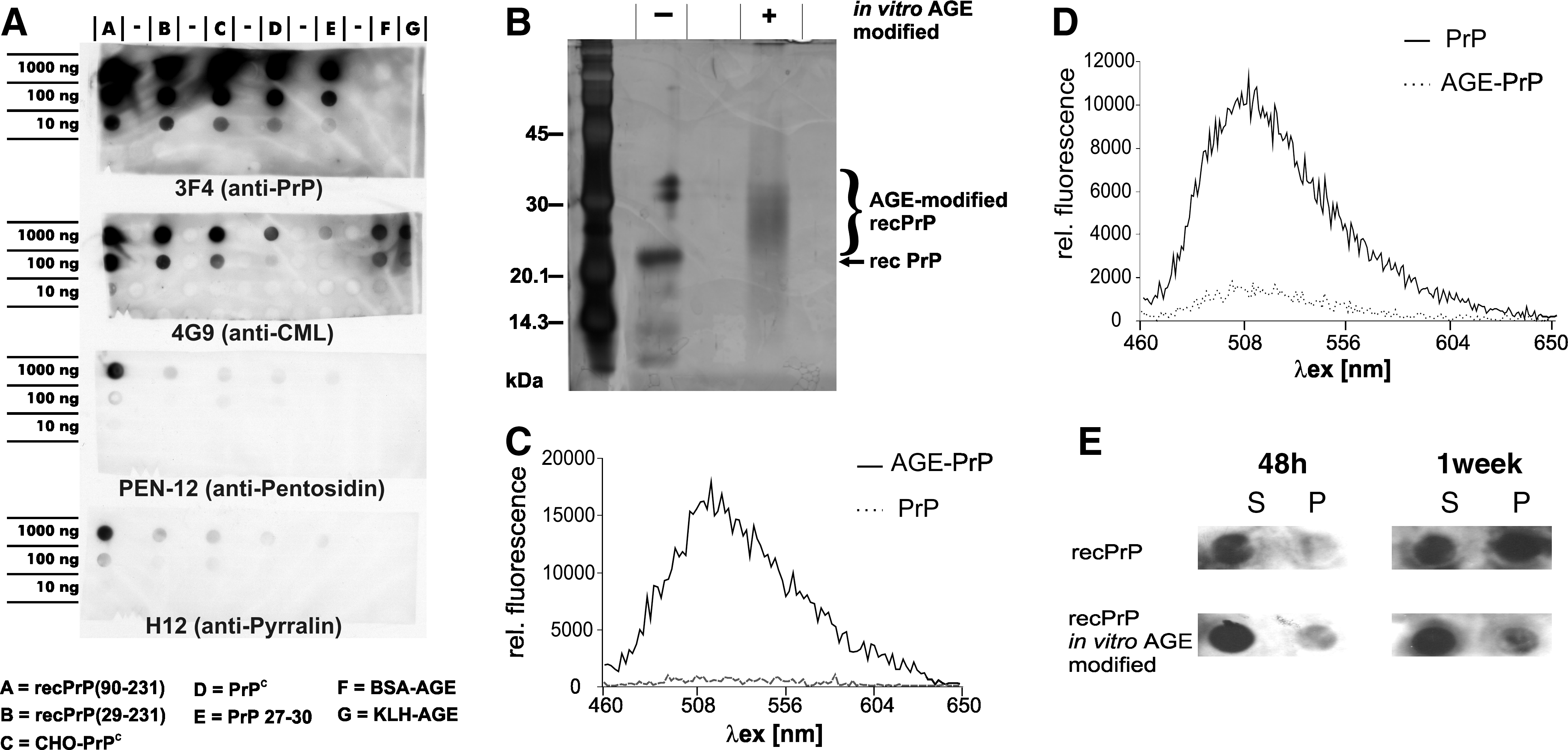
(
To analyze if AGE-modification of PrP has an effect toward its aggregation characteristics, we adopted a method for in vitro AGE modification 15 and applied it to recPrP(29–231). In vitro modification of AGE-PrP was verified by silver-stained PAGE. The increase in molecular weight caused by in vitro AGE modification of PrP could clearly been shown. Additionally, the in vitro AGE modification of PrP was also confirmed by AGE-specific autofluorescence (data not shown). AGE modification could even be observed by fluorescence of λex = 445 nm and λex = 460–650 nm (Fig. 1C).
The influence of AGE-modification to PrP aggregation was studied applying a PrP-aggregation assay. 14 Within this assay, incubation of PrP under balanced SDS and NaCl conditions leads to formation of amyloid PrP fibrils. AGE-PrP and untreated PrP were incubated for 5 weeks under amyloid formation conditions. After 5 weeks, unmodified PrP displayed an amyloid-specific increase in ThT-fluorescence; in contrast, AGE-PrP showed no increase in ThT-fluorescence (Fig. 1D). Additionally, the solubility of PrP was studied during the early incubation period after 48 h and after 1 week. The soluble and insoluble fractions were analyzed using a PrP-specific antibody (pAbA4). Untreated PrP displayed higher insolubility than AGE-PrP (Fig. 1E). However, under conditions leading to amorphous aggregates, formation of larger insoluble aggregates were observed with AGE-modified recPrP (data not shown).
Discussion
AGEs are suspected to play an important role in several neurodegenerative diseases. 6,7 They can induce protein aggregation and cytotoxicity. 8 With respect to prion diseases, we analyzed prions and preparations of noninfective cellular PrP according to their PrP-AGE content (Fig. 1A). All analyzed PrP preparations were immunopositive for AGEs. Recombinant PrP preparations presented the strongest AGE-positive signals. Therefore, AGEs should be considered when using recombinant PrP, especially if cytotoxicity of in vitro–generated PrP aggregates should be tested. Infectious PrP27–30 exhibited the lowest AGE-positive signals. One could expect that prion aggregates contain a higher AGE amount due to their long molecular lifespan compared to the fast-turnover cellular PrP. Therefore, the different distribution of AGEs in PrP27–30 and PrPC might represent a different history of exposition to carbonyl stress during the individual biology of the molecules. A low AGE content in PrP27–30 can provide hints that the conversion takes place early after PrP expression and is followed by protection against further AGE modification within prion aggregates, which has to be established in the future.
We observed that in vitro AGE-modification of PrP led to an inhibition of PrP fibril formation. Others have reported that the Maillard reaction using glucose as a reducing sugar strongly decreased the infectivity of prions. 21 Combined, these results lead to the conclusion that the Maillard reaction could play a preventive role either in the phase of pathological prion protein aggregation or after the formation of infective prions.
Footnotes
Acknowledgments
The authors are indebted to Stanley B. Prusiner (University of California, San Francisco, San Francisco, CA) for kind gifts of prion materials, Roche Diagnostics GmbH, Penzberg for kind gifts of 4G9 antibody and KLH-AGE, and Jörg Tatzelt (LMU, Munich, Germany) for kind gifts of A4 antibody. Stimulating discussions with Dr. Rosemarie Kientsch-Engel, Roche Diagnostics GmbH, Penzberg, are highly acknowledged. This work was supported by the EU-Network of Excellence "NeuroPrion" (FOOD-CT-2004-506579).
Author Disclosure Statement
No competing financial interests exist.