
Abstract
Effects of stem and progenitor cells or their compounds on recipient cells are investigated intensively today. In spite of this, their ability to interact with native cells and the final targets affected by them, particularly biochemical parameters that characterize cell redox-dependent processes, remain little studied. We have studied how bioregulators of stem and progenitor cells affect these processes in freshly isolated liver after animal pretreatment in vivo. Cytosol of human fetal mesenchymal–mesodermal tissues (8–10 weeks gestation) was administered intravenously; the control group was treated with Hanks' solution. After 4 hr, rats were sacrificed and their livers were isolated. To evaluate liver redox-dependent state, mitochondrial respiratory activity and nitroxyl radical and Alamar Blue™ reduction rates, mitochondrial and cytosolic glycerol kinase and nicotinamide adenine dinucleotide (NADH)-dependent malate dehydrogenase activities were studied. The results obtained demonstrate that bioregulators strongly affect liver redox-dependent processes, increasing mitochondrial respiration in state III and spin probe reduction rate and enhancing Alamar Blue™ reduction by glycolytic and nonglycolytic postmitochondrial enzymes. In addition, mitochondrial glycerol kinase and both isoforms of NADH-dependent malate dehydrogenase were inhibited. These data bring us closer to understanding stem and progenitor cell effects via directed regulation of metabolic redox-dependent processes.
Introduction
Another side of this problem is the widening of commercial propositions to rejuvenate and to revitalize the human organism by various stem or/and progenitor cell preparations, and extracts. These programs often result in improvement of the psycho-physical state of aged recipients. Some of these programs are controversial because they are of doubtful reputation and the mechanisms and pathways responsible for their effects are unknown.
It is well known that developing tissues are characterized by the highest content of stem and progenitor cells at early stages of embryogenesis. Thus, we decided to use fetal tissue cytosol (FTC) of mesenchymal–mesodermal origin consisting of protein and peptide bioregulators for animal pretreatment before stress influence or for support of experimental pathology. It has been shown that FTC administration to animals in vivo for 4 hr before acute toxic hepatitis induction 1 or liver isolation for hypothermic storage 2 led to a significant decrease in organ injury degree and supported liver prooxidant/antioxidant balance during 3 days in the first case and 24 hr in the second one. In models of experimental cirrhosis 3 and chronic alcohol poisoning, 4 FTC was introduced to animals with generated pathology and the observation term was prolonged to 2 weeks. In these cases, FTC efficacy was also shown, especially its capacity to stimulate liver reparative processes and partially restore injured organ functional state. Thus, the community of observed effects was almost independent on experimental model and this fact undoubtedly, closely with universality of targets for FTC action.
In addition, in the model of acute toxic hepatitis in rats, we showed that efficacy of pretreatment strongly depended on the term of FTC administration. It was introduced for 72, 24, or 4 hr before poisoning, and the most prononced and prolonged effect was observed in the last case. 5 Also, the specificity of bioregulator effects was clearly confirmed in the same model using α-fetoprotein and bovine serum albumin (BSA) for comparison. 6
The present study focused on the response of liver cell redox-dependent processes to the pretreatment of animals with FTC. This focus was chosen because support of redox balance is one of the crucial factors of cell survival, even under conditions of drastic degradation of energy-supplying compounds. 7
The aims of our research were to investigate: (1) the liver mitochondria functional state depended on potential and redox balance; (2) the compartmental redox state estimated by Alamar Blue™ (AB) dye reduction; and (3) the activities of glycerol kinase and malate dehydrogenase (MDH) that play important role in cell redox balance and take part in the malate–aspartate and glycerol phosphate shuttles between mitochondrial oxidative phosphorylation and cytosolic glycolysis.
Materials and Methods
Obtaining of stem and progenitor cell bioregulators
FTC from human mesenchymal–mesodermal tissues (8–10 gestation weeks) was used as a source of bioregulators. It was obtained as described previously 1,2 and consisted of only water-soluable compounds. Fetuses were obtained from routine surgical termination of pregnancy in which patients had volunteered to donate fetal tissue under informed consent for therapeutic use and research as dictated by the Ukraine Ministry of Health. The study was also approved by the institutional ethical committee. FTC was standardized by protein content (1±0.2 mg/mL) and was stored by freezing in liquid nitrogen in 1-mL cryovials before use.
Experimental model
Female white rats weighing 200–250 grams (n=14) were used in our experiments. Animals were maintained under standard conditions with free access to food and water. Rats were housed and treated as dictated by the guidelines of the Ukrainian Academy of Sciences and The Local Institute Committee on Ethics and Welfare. All surgical interventions in animals, including decapitation, were performed under anesthesia induced by ether inhalation.
There were two experimental groups (n=7 in each): (1) Sham-pretreated group (Hanks' solution injection) and (2) FTC-pretreated group. FTC was injected into the rat femoral vein in a dose of 0.3 mL/100 grams of weight. The control group received an equal volume of Hanks' solution. After 4 hr, the animals were anesthetized and cold liver flushing in situ was performed with physiologic solution. The organs were isolated and samples of liver tissue were homogenized.
Mitochondria isolation
Mitochondria were isolated after tissue homogenization, standard differential centrifugation in sucrose medium, and resuspension in the isolation medium. 8 Livers were homogenized in ice-cold isolation medium (250 mM sucrose, 10 mM Tris-HCl, 1 mM EDTA, pH 7.4). Final protein concentrations of the mitochondrial suspensions were 40–50 mg/mL.
Mitochondrial respiration assays
Respiratory parameters were studied by Chance using Clark-type electrodes in a thermostatically controlled chamber (1 mL) at 26°C with Rank Brothers Ltd. model 20 oxygen monitor (Bottisham, Cambridge, UK) connected to a personal computer. Samples for respiratory measurement contained 2.0–2.5 mg of protein and the assay medium consisted of 200 mM mannitol, 50 mM sucrose, 1 mM EDTA, 10 mM potassium dihydrogen phosphate (KH2PO4), and 30 mM Tris-HCl buffer (pH 7.4) as described previously. 8 The respiratory substrates were 5 mM malate, 5 mM glutamate, and 8 mM succinate. Oxidative phosphorylation was initiated by addition of 250 μM adenosine diphosphate (ADP) to activate respiration state III (V3), while resting respiration state IV (V4) was measured after ADP depletion. The calculation of the oxygen consumption rate was performed according to Estabrook. 9 From the ratios of oxygen consumption in state III and state IV (V3/V4), the respiratory control index (RCI) was calculated.
Electron paramagnetic resonance spectroscopy of nitroxyl radical reduction by mitochondria
Mitochondrial redox activity was studied by the electron paramagnetic resonance (EPR) method using a water-soluble spin probe TEMPONE (2,2,6,6-tetramethyl-4-oxopiperidine-1-oxyl, Aldrich, USA). The spectra of EPR were recorded with a Bruker ER 100D spectrometer (Germany) with standard thermal attachment. The sweep of the magnetic field was 100 Gs, response time was 0.5 sec, and sweep time was 100 sec. Glass capillaries with an inner diameter of 500 nm and a 0.1-mL volume were used. The control experiments showed that under the conditions described the spectra were not distorted due to overmodulation or inertial effects.
For standardization of experimental conditions, the EPR signal of the standard crystal with chrome inclusions was recorded at the same time with the EPR probe signal. As a kinetic parameter of spin probe reduction, the relative time changes of mean field component amplitude of probe EPR spectra (ho) were used.
AB reduction by liver postmitochondrial fractions
AB is a redox indicator that changes its color from blue to pink during reduction. AB has the ability to fluoresce, and fluorescence intensity is directly proportional to activity of redox enzymes. 10
In our experiments, liver postmitochondrial fractions (PMF) obtained during mitochondria isolation were used. An amount of 0.1 mL of 10% AB was added to 1 mL of incubation medium. Monoiodoacetate (MIA) at a final concentration 10 mM was used as a glycolysis inhibitor. Then 0.2 mL of PMF was added to each probe in a 96-well plate (Corning, Canada), and samples were incubated for 5 and 10 min with constant mixing. The parallel samples MIA and AB were placed in wells containing just incubation medium. AB reduction degrees were determined fluorimetrically using a spectrofluorimeter (Tecan GENios, Tecan Inc., Australia) with an excitation wavelength of 550 nm and an emission of 590 nm. Data were processed with the software XFLUOR4 v.4.50. The results were presented as the difference between fluorescence of experimental and bare probes and were expressed as standard fluorescence unit (SFU).
Determining glycerol kinase and malate dehydrogenase activity
Enzyme activities were studied using 20% (wt/vol) homogenate prepared in 10 mM Tris-HCl buffer containing 0.3 M sucrose and 5 mM EDTA (pH 7.4). Glycerol kinase (EC 2.7.1.30) and nicotinamide adenine dinucleotide (NADH)-dependent-malate dehydrogenase (MDH; EC 1.1.1.83) activities were measured using a Cary-50 spectrophotometer (Australia) in a thermostatically controlled (37°C) cell with continuous mixing by NADH depletion in coupled reactions. Briefly, assay conditions were: wavelength, 340 nm; for glycerol kinase—0.05 M KH2PO4/dipotassium phosphate (K2HPO4; pH 7.4), 20 mM glycerol, 20 mM adenosine triphosphate (ATP), 20 mM MgSO4 11 ; for MDH—0.1 M KH2PO4/K2HPO4 (pH 7.4), 0.2% Triton X-100, 0.15 mM NADH, and 0.33 mM oxaloacetate. 12
Enzyme activities were expressed as nmol NADH/mg of protein/min.
Protein assay
Protein concentration in homogenates was determined by the biuret method with BSA as the standard. Assays were performed at 540 nm.
Statistical analysis
Results are expressed as mean±standard error of the mean (SEM) (M±m). Statistical data manipulations were performed by analysis of variance (ANOVA), and post hoc analyses done using the Student t-test by means of the GraphPrizm 4.0 statistical program package; differences were accepted significant at P<0.05.
Results
Liver mitochondrial respiratory parameters after rat FTC pretreatment
Respiratory activity of isolated mitochondria is a very sensitive indicator of organ state, which changes after administration of biologically active substances, hormones, 13 and toxic agents. 14 As shown at Table 1, mitochondrial respiratory activities in the control group during malate+glutamate or succinate oxidation were characterized by low rate of oxygen consumption in the steady state (V4) and by a high RCI that appropriates to intact mitochondria.
nmol O2/mg of protein/min (M±m; n=7–9).
p<0.05 as compared with control group.
FTC, Fetal tissue cytosol; V4, respiration state IV; V3, respiration state III; RCI, respiratory control index.
After FTC administration, oxygen consumption rates in state III at succinate oxidation were significantly higher than in control group but were insufficient for RCI increase. In the case of NAD-dependent malate+glutamate addition, both V4 and V3 did not differ from the control group (Table 1).
EPR spectroscopy of nitroxyl radical reduction by mitochondria after rat FTC pretreatment
It was shown earlier that nitroxyl radical TEMPONE could be applied for estimation of carrier relative activities in the mitochondrial respiratory chain. 15 The mean of half-reduction potential for this spin probe amounted about+48 mV, implying mitochondrial complex III (with redox potential is approximately+40 mV) as the donor of protons and electrons, although the question regarding which of the complex III subunits are able to reduce TEMPONE remains open. It was just shown that the EPR signal was increased when mitochondria were inhibited by antimycin A and potassium cyanide (KCN) showing evidence of participation of Fe-S centers in electron transport and TEMPONE protonation. 15
As shown in Fig. 1, FTC pretreatment led to a significant increase of the spin probe reduction rate. This fact indicates that bioregulators are able to promote the speed of endogenous substrate oxidation and electron transport in the succinate oxidase unit of the respiratory chain.
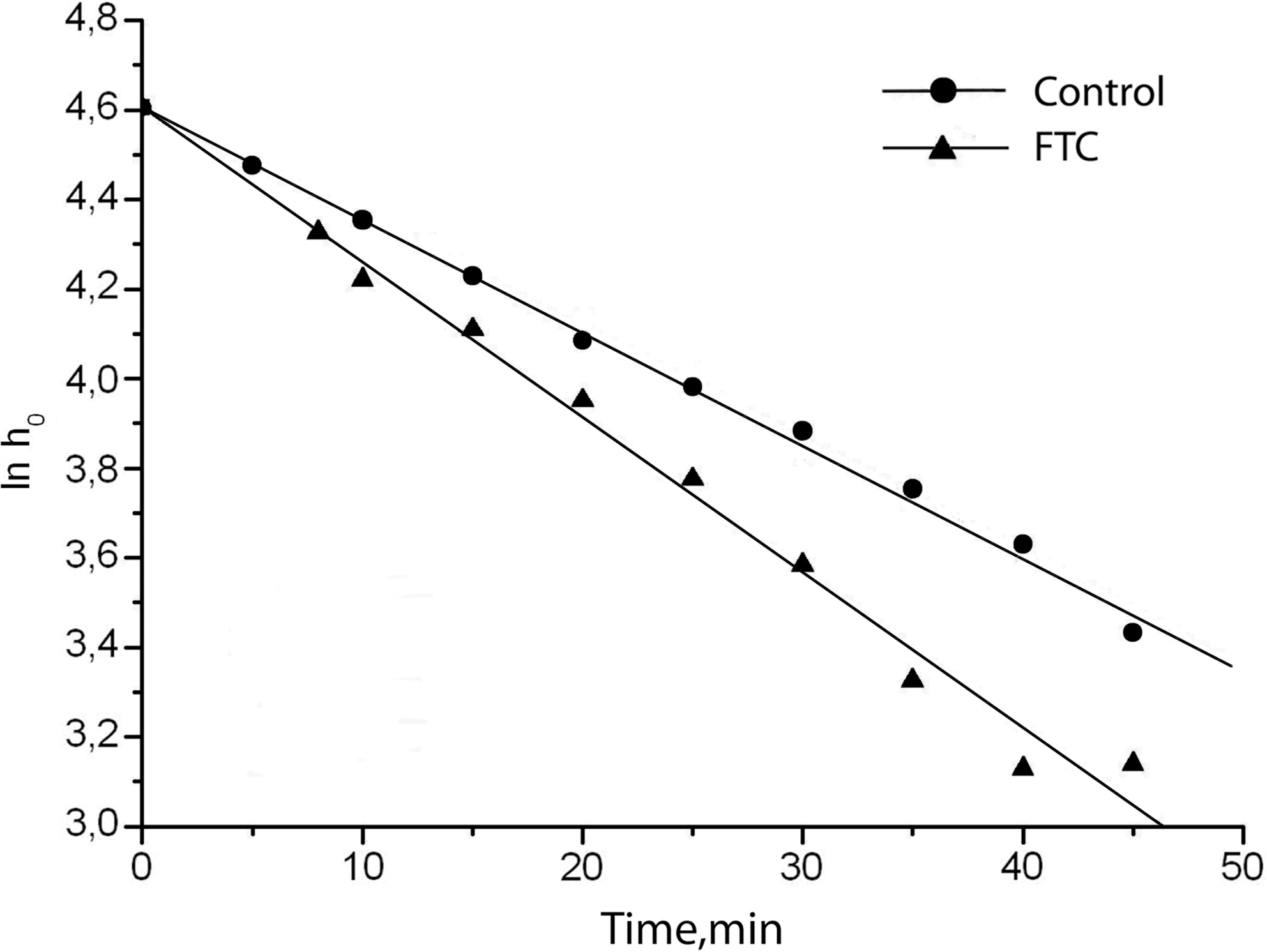
Rate of spin probe TEMPONE reduction by liver mitochondria after rat fetal tissue cytosol (FTC) pretreatment.
AB reduction in liver postmitochondrial fractions after rat FTC pretreatment
According to existing data, the degree of AB reduction is directly proportional to redox enzyme activities. 16 It was shown earlier that the main contributor to AB reduction is the mitochondrial respiratory chain, and the application of its inhibitors resulted in significantly decreasing the ability of AB to be reduced by mitochondria as well as by whole cells. 17
Because the mitochondrial functional state was already studied by commonly used methods of polarography and EPR-spectroscopy, we decided to focus our attention on the redox state of nonmitochondrial liver cell components such as cytosolic, microsomal, and other enzymes. Glycolysis as source of macroergs was of particular interest for us. For the study of the contribution of glycolysis, its inhibitor MIA, which blocked the metabolic pathway at the glyceraldehyde 3-phosphate dehydrogenase stage, 18 was used.
As shown on Fig. 2, after 5 min of incubation with AB in the control group, 75% of total fluorescence was related to glycolytic component. FTC pretreatment led to an increase of glycolysis intensity 2.2 times that of the control and in the nonglycolytic compound 3.8 times that of the control. The component ratio was changed also; in the case of the control, it was 3:1 and after FTC application it was 1.86:1.
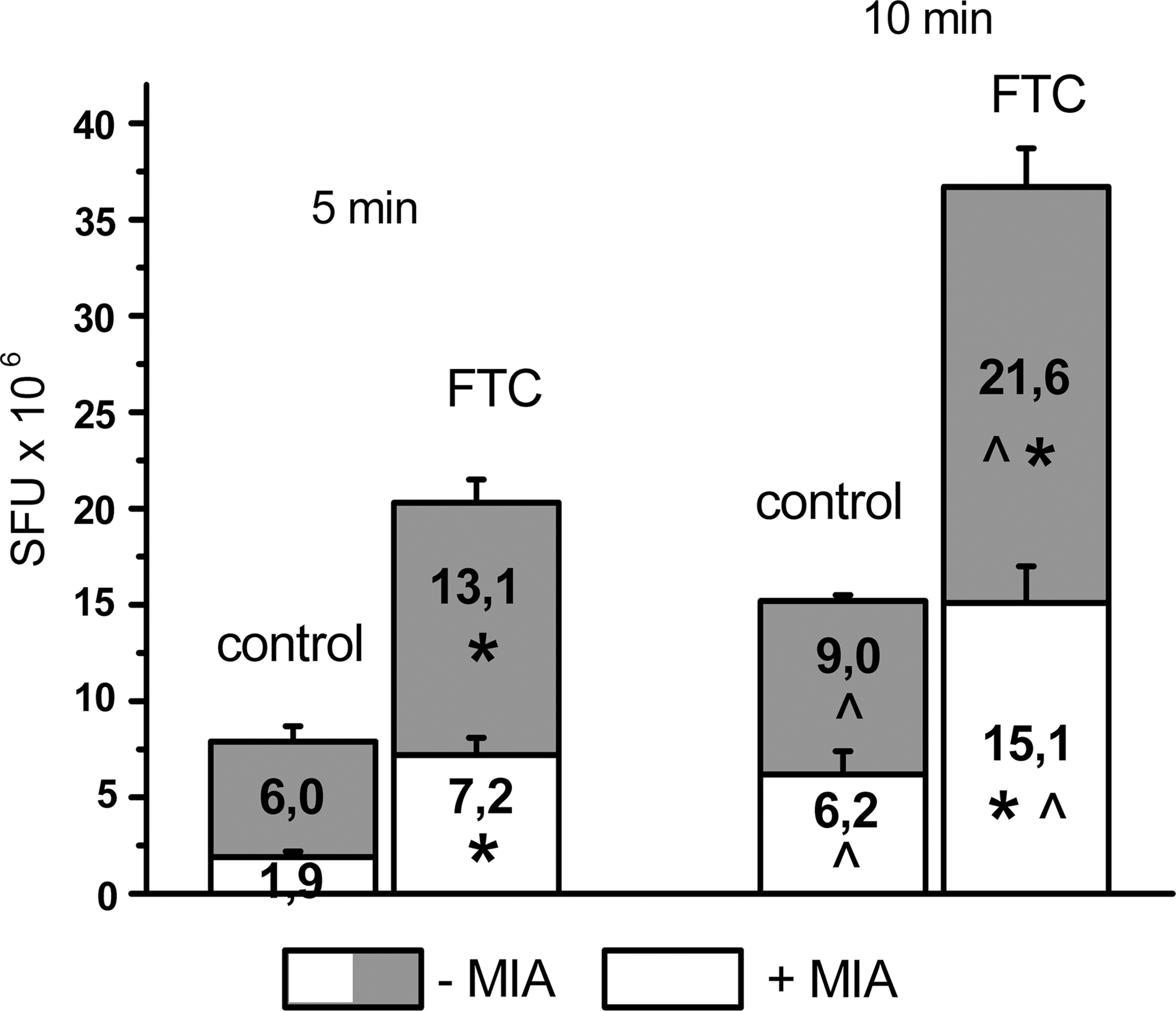
Fetal tissue cytosol (FTC) pretreatment effect on Alamar Blue™ (AB) reduction by liver postmitochondrial fractions (PMF) after 5 and 10 min of incubation in the presence or absence of glycolysis inhibitor monoiodoacetate (MIA). (*) p<0.05 as compared to control group; (^) p<0.05 as compared to 5 min of incubation. SFU, Standard fluorescence unit.
After 10 min of incubation, some differences in AB reduction dynamics were observed that looked like “lag-phase” of the interaction between indicator and redox systems. In the control group as well as in the FTC-pretreated group, both components were increased, although in last case the degree of AB reduction was significantly higher than in the control. In the sham-treated group, the total fluorescence intensity was raised 1.9 times; in the glycolytic component, it was raised 1.5 times and in the nonglycolytic one 3.3 times (Fig. 2). The compound ratio was about 1.45:1. In the FTC group, the total intensity was increased 1.8 times, glycolysis activity 1.6 times, and the nonglycolytic component 2.1 times; the ratio was 1.43:1.
Thus, data obtained after 10 min of incubation with AB showed comparable contribution of redox processes to the total liver cell activity; however, in the case of FTC pretreatment, their rate was significantly accelerated. This fact may be confirmed by calculation of AB reduction conventional rates between 5 and 10 min of incubation, which for the control group were: Total, 1.46; glycolytic component, 0.6; nonglycolytic, 0.86 SFU·106/min. For the FTC-pretreated group, these indices were: Total, 3.28; glycolytic,1.7; nonglycolytic, 1.58 SFU·106/min.
Consequently, activation of both glycolytic and nonglycolytic redox systems by FTC and the increase of the succinate reduction rate (Table 1) suggest that bioregulators of stem and progenitor cells have the strong potential to affect activities of enzymes involved in transport of reduced equivalent from cytosol to mitochondria. Cytosolic and mitochondrial glycerol kinase and NADH-dependent MDH were selected in our research.
Liver glycerol kinase and malate dehydrogenase activity
As shown at Fig. 3A, more than 90% of glycerol kinase activity is attributed to the mitochondrial pool. This fact is comparable with the data of McCabe, which showed enzyme localization on the external surface of mitochondrial outer membranes. 11 In the control group, liver homogenate glycerol kinase activity was 10.10±0.88 and cytosolic was 1.37±0.11 nmol NADH/mg of protein per min. After FTC pretreatment, enzyme activity in homogenate was in 1.8 times (p<0.05) lower than in the control group; no differences were observed in PMF.
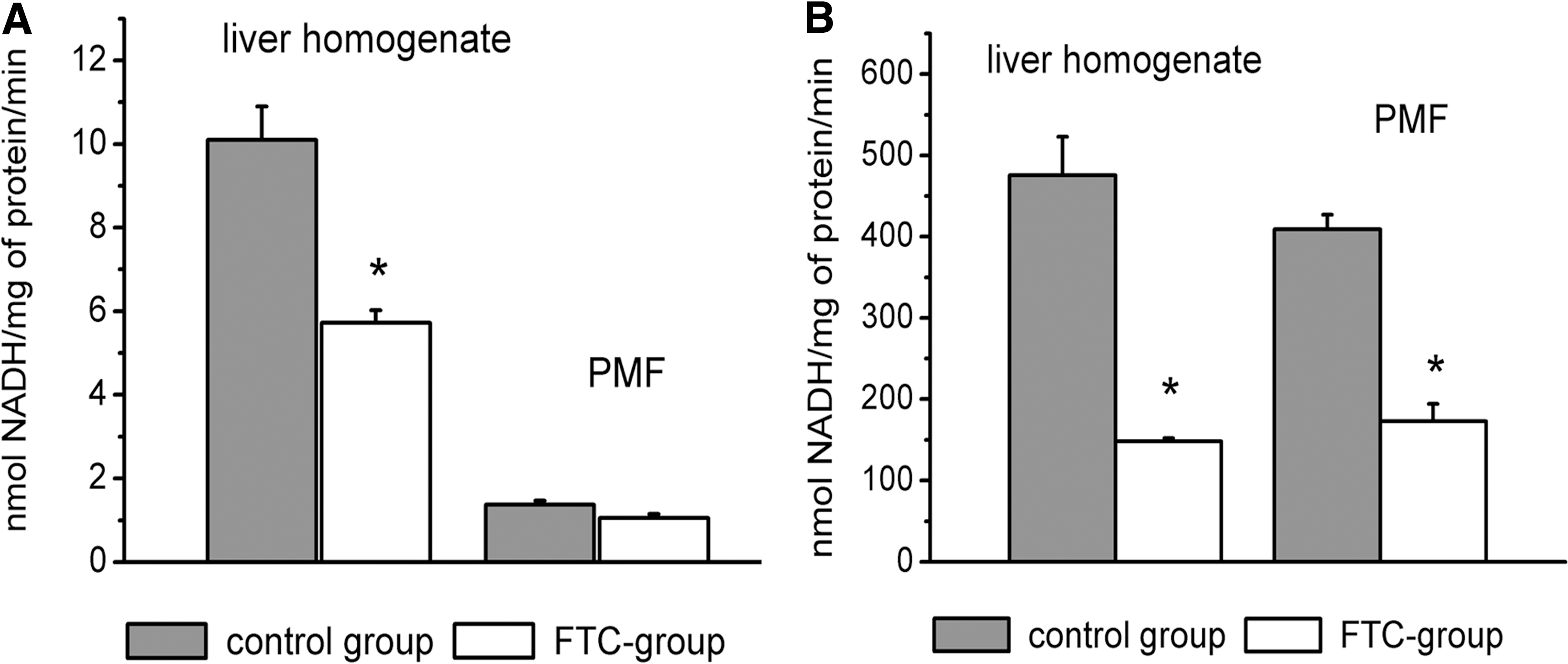
Glycerol kinase (
In comparison, NADH-dependent MDH was maximally presented by the cytosolic isoform: 475.8±23.5 nmol NADH/mg of protein per min in liver homogenates and 409.2±17.8 in PMF. FTC pretreatment led to decrease of MDH activity at 4 times in homogenates and 2.4 in PMF (Fig. 3B).
Discussion
Stem and progenitor cells of fetal origin contain a small number of mitochondria with poorly developed cristae. Their content increases and structure is perfected in close correlation with the degree of cell commitment. 19 On the basis of this principle, poor mitochondrial genesis is the most important feature of fetal cells and is associated with their acquired resistance to hypoxia and high activity of anaerobic glycolysis, depending on the conditions of prenatal development. These properties are probably common and universal for all stem cells. It was shown that mesenchymal stromal cells were resistant to ischemic conditioning and survived for 3 days in the presence of the mitochondrial oxidative phosphorylation uncoupler 2,4-dinitrophenol, maintaining the capacity to follow differentiation. 20 Undoubtedly, regulatory mechanisms supporting such functional states should exist. In our opinion, the main participants of these pathways are stage-specific, biologically active substances. Screening of noncellular extracts of mesenchymal stromal cells from various sources detected a wide spectrum of regulatory molecules of protein and peptide origin, such as growth factors (of hepatocytes and fibroblasts, epidermal, insulin-like, transforming (α andβ), neurotrophic), cytokines (tumor necrosis factor-β, interleukin-1α (IL-1α, −6, −7, −8, −9), chemokines and other substances. 21,22 We separated five peptide fractions (800–3,000 Da) and 15 protein fractions (molecular weight 9–110 kDa) from FTC by gel chromatography and gel electrophoresis. 23 α-Fetoprotein content in FTC was extraordinarily higher than in extracts from fetal liver or neural cells of the same gestation term. 6 Recently, we detected a high level of insulin-like growth factor-1 (IGF-1) in our preparation (unpublished data).
Our conception is based on the view that just a “physiologic cocktail” is capable of providing the integral effects affecting all metabolic pathways in target cells and provides considerable support to adult cells during stress. This concept is confirmed by our earlier results 1 and agrees with the opinions of other authors. 22 The possible explanation of this support is activation of adaptation mechanisms in mature cells that are usually inherent to undifferentiated ones.
Independent of the type of stress, the activated cascade of response reactions is rather universal, with some peculiarities regarding their priority in each specific case. These reactions refer to oxidative stress and disturbances of energetic balance, cytokine profile, and membrane state. All of these reactions are characteristic of the experimental models of acute and chronic liver failure that we investigated earlier. 1 –4 As a result of FTC administration, we observed that: The decrease of serum markers of injury (in all models) indicated the enhancement of cell cytoplasmic membrane stability; the improvement of mitochondrial functional state (in the models of experimental cirrhosis and liver long-term hypothermic storage); the prevention of ATP level and glycolysis activity fall (in short-term liver storage); and the normalization or improvement of organ pro-oxidant/antioxidant balance (in all models). In the cases of experimental cirrhosis and chronic alcohol poisoning, the stimulation of reparative processes following by decrease of inflammatory reaction was observed.
The present results in some way disprove existing opinion that one of the main peculiarities of stem and progenitor cell action is their ability to detect pathological foci and to affect the injured area without an influence on healthy areas. Our data demonstrate that stage-specific, biologically active substances are capable of successfully interacting with native cells, changing their properties by redirection of metabolic pathways. These changes result in more efficient cell adaptation to stress.
Impairment of Complex I is a first stage of mitochondrial oxidation disturbance. These alterations consist of disturbances in electron transport between NADH and coenzyme Q and cause a sharp decrease in total oxidative phosphorylation output. 24,25 In spite of a drop in the NAD+-dependent oxidation rate, the intracellular macroerg level at an early stage of stress decreases insignificantly, whereas alternative compensatory metabolic pathways are activated. For example, succinate oxidase is a powerful source of reduced equivalent in the respiratory chain and, in some cases, succinate oxidation is capable of monopolizing the respiratory chain. 26 Therefore, the increase of succinate oxidation rate after FTC administration may be a result of increased activity of redox enzymes, which provide accumulation of endogenous substrates in mitochondria. This suggestion is consistent with the EPR-spectroscopy data. The possible explanation of the high rate of spin probe TEMPONE reduction after animal pretreatment with FTC is the presence of endogenous substrates like succinate, aspartate, and fatty acids that can be oxidized by mitochondria.
The intensity of metabolic pathways after FTC pretreatment was studied using the redox indicator AB. At present, the key role in AB reduction is assigned to mitochondria. 10,17 Also, cytosolic and microsomal enzymes can take part in this reaction. 27,28 Our results show that MIA inhibited AB reduction in PMF more than in three times. This indicates that glycolysis is one target of FTC. Most likely, this effect is mediated by growth factors and cytokines that are capable of affecting this pathway. It was shown, for example, that epidermal and insulin-like growth factors modulate the activities of the key glycolytic enzymes-phosphofructokinase and piruvate kinase by reversible phosphorylation. 29
Animal pretreatment with FTC activates not only glycolytic but also nonglycolytic redox enzymes in cytoplasm. Taking into account the enhancement of succinate oxidase activity, we studied the compounds of cytosol-mitochondria shuttles—MDH and glycerol kinase. The choice of the latter enzyme is based on close correlation between its and glycerol-3-phosphate dehydrogenase's (GPDH) activities. 30
It is well known that cytoplasmic NADH is not able to penetrate the inner mitochondrial membrane; therefore, it is transported by substrate shuttle mechanisms. Thus, during glycerol phosphate shunt function in the cytoplasm, dihydroxyacetone phosphate is transformed to glycerol-3-phosphate, which easily penetrates the mitochondrial membrane, where it is oxidized to dihydroxyacetone phosphate again. Reduced flavoprotein passes electrons to coenzyme Q and further to the respiratory chain; dihydroxyacetone phosphate moves from mitochondria to the cytoplasm and becomes able to interact again with NADH. The malate–aspartate shuttle consists of malate and cytosolic and mitochondrial isoforms of MDH. The functional role of this system lies in the fact that the mitochondrial membrane is impermeable to oxaloacetate, and by means of the shuttle α-ketoglutarate and aspartate are transported to the cytosol.
It was shown that animal FTC pretreatment led to a significant decrease of MDH and glycerol kinase activities, supporting our suggestion about endogenous substrate accumulation by mitochondria and their switching over to succinate oxidase flavin adenine dinucleotide (FAD)-dependent pathway of oxidation combined with inhibited NADH-transporting shuttle activities.
The data obtained indicate that stage-specific bioregulators significantly alter liver redox-dependent processes and cause changes in cell functional state. In particular, the mechanisms of immediate adaptation to hypoxia are activated, and this fact is supported by enhanced succinate-linked oxidation in mitochondria. This process is accompanied by activation of glycolysis and other redox-dependent reactions beyond mitochondria. Our results detail the mechanisms of cell mobilization by biologically active substances of fetal origin, one of which is the directed regulation of metabolic redox-dependent processes. The results obtained bring us closer to understanding how stem and progenitor cells produce their effect during stress. This data may serve as background for practical application of stem and progenitor cell preparations in programs of revitalization and rejuvenation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.