
Abstract
Alzheimer's disease (AD) is the most common form of dementia, characterized by the presence of two principal hallmarks—amyloid plaques and neurofibrillary tangles. The primary cause of the majority of AD cases is not known. Likewise, the mechanisms underlying the propagation of the pathology from affected tissue to neighboring healthy neurons are largely unknown, but knowledge about them could be helpful to design strategies aimed at halting the progression of the disease. To throw light on the mechanisms of propagation of neuronal damage to healthy tissue, wild-type (WT) hippocampal solid tissue chunks derived from green fluorescent protein (GFP)-positive embryos were grafted into the hippocampus of 6-month-old WT and 3xTg-AD mice, a triple-transgenic mouse model that exhibits both amyloid-beta (Aβ) and tau protein pathology. The histological and morphological alterations of the grafted tissues were assessed 3 months post-transplantation. Tissues grafted in 3xTg-AD hosts, compared to those grafted in WT recipients, presented a significant decrease in neurite outgrowth (35.4%) and dendritic spine density (41.3%), mainly due to a reduction of stubby and thin-shaped spines. Moreover, some cells of the tissue transplanted in 3xTg-AD hosts accumulated intracellular amyloid peptide deposits similar to the cells of the host. Furthermore, the immunohistochemical examination of reactive astrocytes and microglia revealed the presence of more inflammation in the grafted tissues hosted in 3xTg-AD compared to WT recipients. These results show a propagation of neuronal damage to initially healthy embryonic grafts, validating this methodology for future studies on the mechanisms of the progression of AD pathology to surrounding regions.
Introduction
A
For unknown reasons, the pathological lesions start in specific areas of the brain located in the medial temporal lobe, including the hippocampal formation. From this region, the lesions propagate to neighboring areas, but the mechanisms underlying such spreading remain a great challenge for neurological research.
Although a central etiology for AD has not yet been determined, one well-recognized and supported explanation for neuronal degeneration is the amyloid hypothesis. 4,5 This hypothesis states that the accumulation and deposition of Aβ, a proteolytic product of the amyloid precursor protein (APP) in the brain, is the primary cause driving AD pathogenesis and leading to neuronal dysfunction and cell death. Consistent with this view, it has been demonstrated, for instance, that transgenic mice overexpressing both mutant human APP and mutant human tau protein undergo increased formation of tau-positive tangles as compared with mice over-expressing tau alone, whereas the structure and number of their amyloid plaques are essentially unaltered. 6 This indicates that altered APP processing can precede and trigger tau alterations in the pathogenic cascade of AD.
Although the amyloid cascade might be responsible for the initiation in particularly sensitive brain areas of pathological lesions, their diffusion to the rest of the brain could be explained by a different mechanism. Prion-like dissemination is a hypothesis to explain the propagation of the disease. It claims that AD is driven by an infection-like spread of protein aggregates in the brain. 7 Recently, evidence has accumulated showing that oligomers are the only truly toxic forms of Aβ, not only in vitro but also in animal models, giving weight to this hypothesis. Moreover, tiny clusters of Aβ, widely considered the likely triggers of AD, are infectious enough that they can spread into the brain of young and healthy mice starting from affected brain material. 8 Notably, it has been shown that intraperitoneal (i.p.) administration of Aβ-containing extracts induced Aβ aggregation in the vicinity of brain blood vessels similar to the angiopathy observed in AD patients. 9
Grafting techniques are widely used to investigate the neuronal plasticity of the adult nervous system and to study the principles of neuronal development. In contrast, the use of neurotransplantation techniques to study the propagation of pathogenic events in brain diseases such as prion disorders is less well established. 10,11 Solid pieces of embryonic tissue were used in several studies for intrahippocampal grafting to analyze the survival and the various intrinsic and extrinsic factors affecting the post-transplantation development of grafts. 12 –15 Unlike single-cell suspensions, transplantation of solid pieces allows the use of slightly older donor tissue with more differentiated neurons. Additionally, it also preserves the intrinsic organization of the grafted tissue, facilitating the assessment of the specificity of the development of neuronal connections. 16 Therefore, the intracerebral grafting approach in transgenic animal models, which mimic the pathological characteristics of AD, will be a powerful instrument to elucidate the propagation mechanisms of the disease in vivo and to design strategies aimed at halting its progression.
The objective of the present study was to investigate the propagation of AD-related lesions to healthy solid embryonic hippocampal tissues derived from green fluorescent protein (GFP)-positive embryos, 3 months after their transplantation into the hippocampi of either adult wild-type (WT) or 3xTg-AD mice, a triple-transgenic mouse model of AD that exhibits both Aβ and tau pathology.
Materials and Methods
All animal experimental procedures reported here were approved by the Bioethical Committee of the University of Turin (January 23, 2013) and by the Italian Ministry of Health (Experimental Protocol Communication of January 28, 2013).
Animals
Six-month-old male homozygous 3xTg-AD (n = 11) and their WT SVBL6 littermates (n = 7), were used as recipients in this study to investigate the role of soluble Aβ in the propagation of AD. The 3xTg-AD mice, obtained from Professor LaFerla's laboratory, 17 over-express the familial AD mutations PS1M146V, APPSwe, and mutated Tau (TauP301L). In these mice, at 6 months of age, only soluble Aβ forms are expressed.
Donor embryonic hippocampal tissues were obtained from healthy transgenic mice overexpressing ubiquitously enhanced GFP (eGFP) under the control of the β-actin promoter. 18 Pregnant females at 14–15 days of embryonic age used as donors of the embryos, were deeply anesthetized by i.p. administration of a mixture of ketamine (200 mg/kg; Ketavet, Bayer, Leverkusen, Germany) supplemented by xylazine (50 mg/kg; Rompun, Bayer) in saline. The embryos were then removed by cesarean section. The day following insemination was designated as E0.
Transplantation experiment
The embryos with the highest eGFP expression were detected and selected by the Illumatool Bright Light System LT-9500 (Illumatool Lighting System, Light Tools Research, Encinitas, CA) and collected in a basic medium (saline solution with 0.6% glucose) for the dissection procedure. Hippocampal tissue pieces were dissected from the selected litter and dissociated mechanically into small blocks (∼1 mm). These blocks of hippocampal tissues were then pooled in a new petri dish with cold basic medium (Fig. 1A).
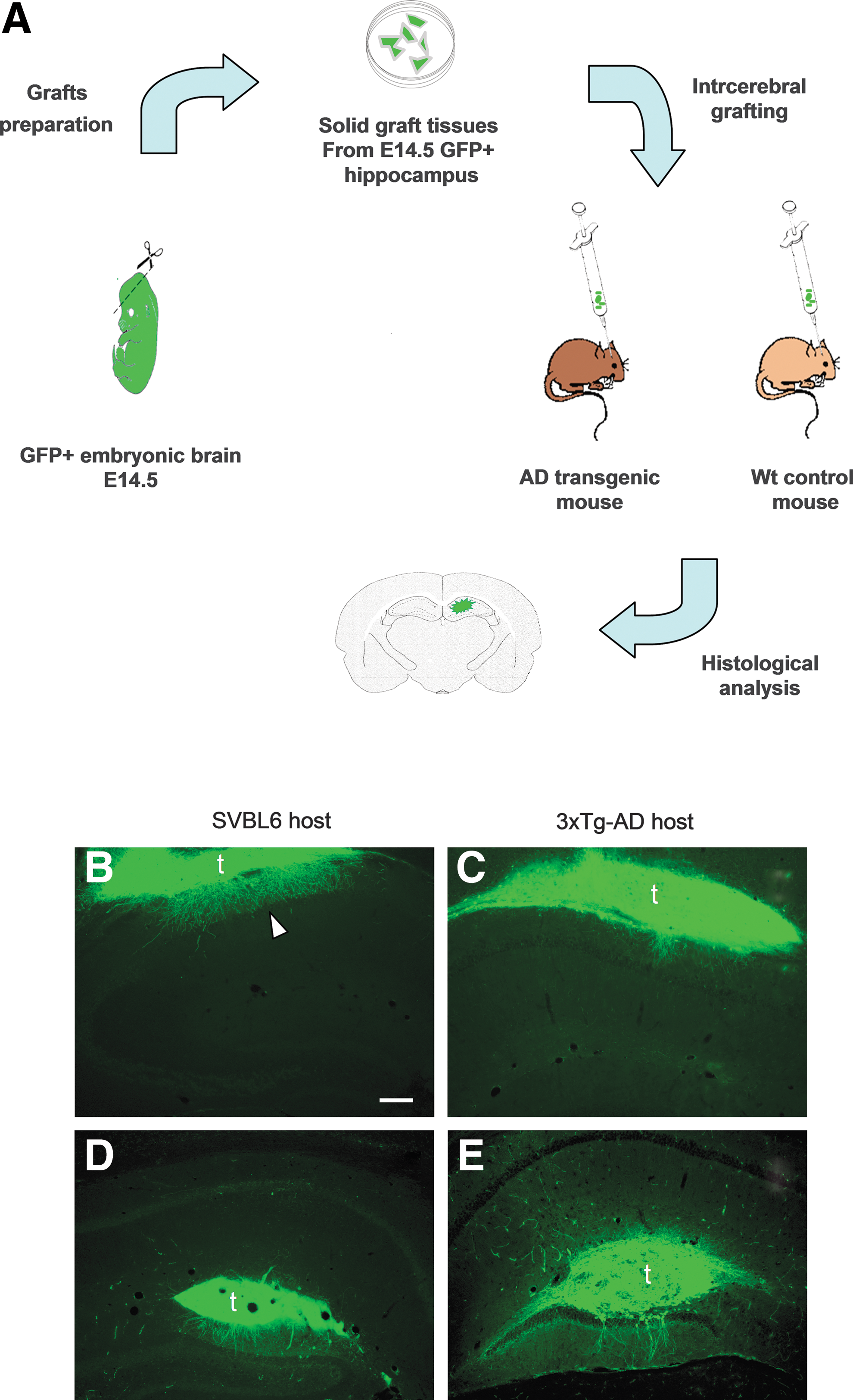
Transplantation procedure and morphological appearance of wild-type (WT) fetal hippocampal tissue following transplantation into the hippocampus of adult 3xTg-AD and SVBL6 mice. (
The host mice were anesthetized by i.p. administration of a combination of ketamine (100 mg/kg; Ketavet, Bayer, Leverkusen, Germany) supplemented by xylazine (5 mg/kg; Rompun; Bayer) in saline. The mice were then placed on a stereotaxic apparatus; the skull was exposed and bilateral holes were made by a drill. Using a 10-μL Hamilton syringe or a 100-μL mechanical pipette connected to the glass capillary, less than 10 μL of medium containing a block of embryonic hippocampal tissue was injected into the hippocampus of either 3xTg-AD or SVBL6 hosts at the following coordinates with respect to the bregma—anterior–posterior (AP), −2.5 mm; lateral (L), ±2.0 mm; dorsal–ventral (DV), −2.5 mm. After the injection, the needle was left in place for 2 min to avoid graft withdrawal before being slowly moved up. The skin was sutured and the animals were returned to their cages.
The recipient animals were sacrificed 3 months post-transplantation. This time point was applied to avoid additional features that could appear later in these transgenic animal models, in particular tau pathology.
Histological procedures
All grafted mice were deeply anesthetized with the ketamine/xylazine solution and perfused transcardially with 100 mL of 0.12 M phosphate buffer (pH 7.2) containing 4% paraformaldehyde (PFA) fixative. The brains were removed immediately and stored overnight in the same fixative at 4°C and finally transferred to a cryoprotective solution of 30% sucrose in 0.12 M phosphate buffer until they sank. Coronal serial sections of 25-μm thickness were cut with a cryostat throughout the brain and collected free-floating in phosphate-buffered saline (PBS).
Free-floating sections were washed in PBS three times before application of the primary antibody. Primary antibodies were incubated overnight at room temperature in PBS containing 0.25% Triton X-100 and 1.5% serum made in the species of the fluoresceinated secondary antibody. 4′,6-Diamidino-2-phenylindole (DAPI) (Fluka, Buchs, Switzerland) was used to counterstain cell nuclei in some reactions. Finally, the stained sections were mounted on microscope slides with Tris-glycerol supplemented with 10% Mowiol (Calbiochem, LaJolla, CA) and coverslipped.
Primary antibodies and dilution factors were as follows: Mouse monoclonal antibody 6E10 specific to hAPP/Aβ (1:300 Signet Pathology Systems, Inc., Dedham, MA), rabbit anti-glial fibrillary acidic protein (GFAP) for GFAP expressed by the astrocytes (1:1000; Dakopatts, Glostrup, Denmark), and rabbit anti-Iba1 for ionized calcium-binding adaptor molecule 1 expressed by microglia/macrophages (1:1000; Wako Chemicals, Richmond, VA). In addition, we also used goat anti-GFP (1:1000; Abcam) and rabbit anti-GFP (1:1000; Molecular Probes) to enhance the intrinsic fluorescence of transplanted neurites and their spines during the acquisition of microscope images at high magnification.
Image analysis and quantification
Immunofluorescence-treated sections were examined with a Zeiss Axiophot microscope (Karl Zeiss, Oberkochen, Germany) equipped with a Nikon DS-5M digital camera (Nikon Italy, Florence, Italy). Selected sections were examined with a Leica TCS SP5 confocal microscope (Leica Microsystems, Milan, Italy). Neurites were imaged at high magnification using a 63× oil immersion lens (z-distance between two serial images, 0.5 μm) and an additional electronic zoom factor so that the morphology of spines could be determined and subsequently quantified. For the spine examination, confocal stacks were analyzed with NeuronStudio
19
–21
(
The total number of GFAP-positive neurons and Iba1-positive neurons in the transplanted tissues was counted by manual semi-quantitative assessment using ImageJ software. Confocal stack images were used to this purpose. The area of cell counting was measured and defined for all stacks, and the density of positive cells (cell/μm2) was calculated. Finally, the neuronal density was converted to the number of neurons per mm2. Neurolucida software (MicroBrightField Inc., Colchester, VT) or Imaris 7.4.2 software (Bitplane AG, Zurich, Switzerland) was used to analyze total length, number, and branching of neurites. Data were statistically evaluated by means of one-way analysis of variance (ANOVA). In all instances, p < 0.05 was considered as statistically significant. Data in the graphs are represented as mean ± standard error of the mean (SEM).
Results
Graft survival and distribution
Three months following transplantation into the hippocampus of 3xTg-AD and SVBL6 mice, graft bulks appeared physically well integrated into the surrounding host. The majority of grafted tissues were detected in the dentate gyrus of the host hippocampus (n = 7), whereas four were in the oriens layer and extended into the corpus callosum (Fig. 1B–E). The cells of the graft islets were counterstained with DAPI to visualize cell nuclei and examine apoptosis (Fig. 2). The presence of intact nuclei without signs of chromatin condensation confirmed the graft survival without ongoing apoptosis. Furthermore, all cells of the tissues transplanted in the hippocampus and their efferent fibers showed GFP expression, indicating again good cellular survival.
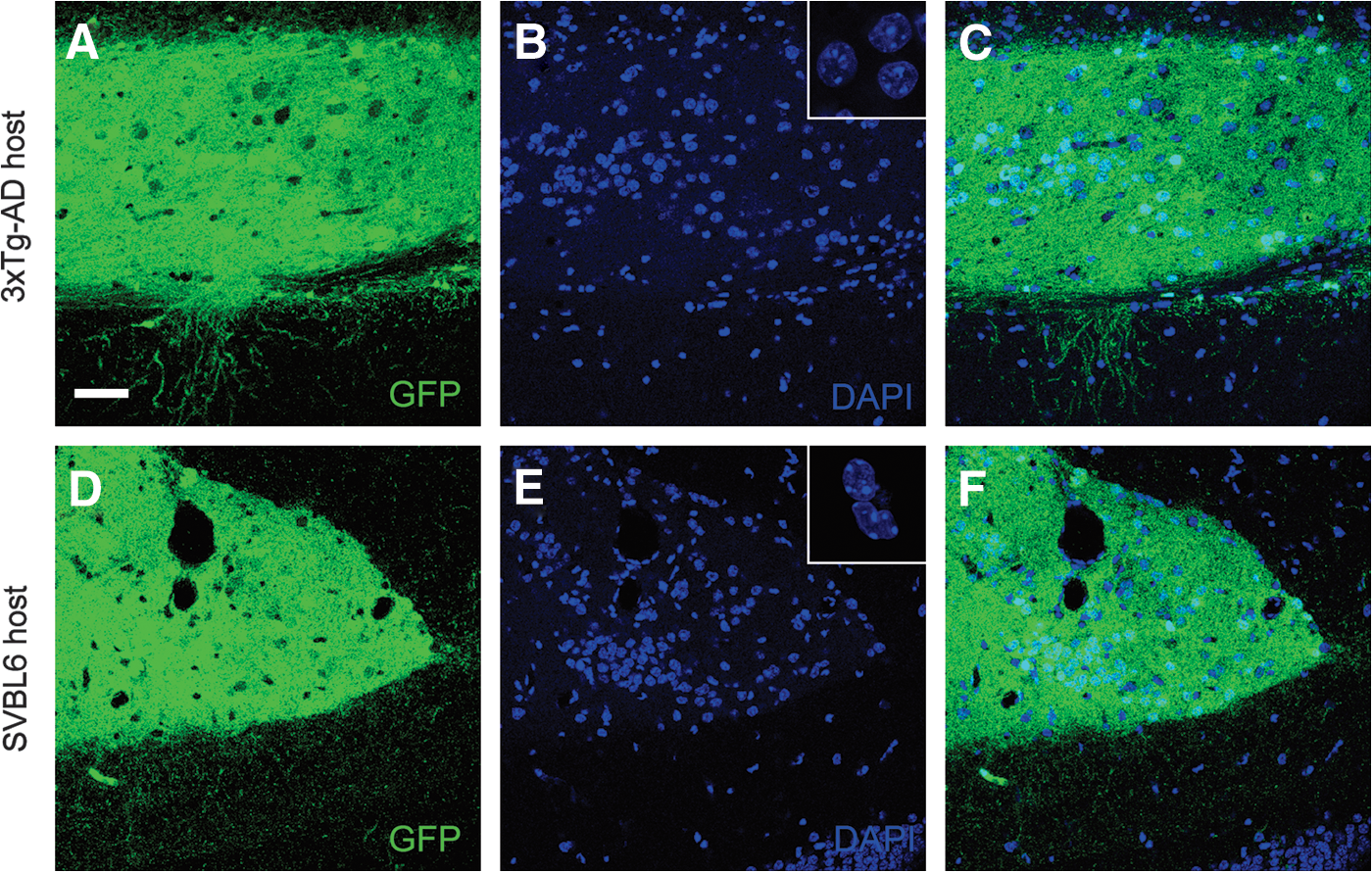
Representative 4′,6-diamidino-2-phenylindole (DAPI)-stained nuclei of coronal brain sections at 3 months post-transplantation. Hippocampal graft tissue in 3xTg-AD host (
Amyloid deposition in the grafted tissues
As a preliminary step of our study, we investigated any possible deposition of amyloid peptide (APP and Aβ) in the transplanted tissue. We used the specific antibody 6E10 that recognizes both full-length APP and APP cleavage products containing the Aβ domain (APP/Aβ). As expected from the transgenes expression, diaminobenzidine immunohistochemistry revealed an abundant expression of intracellular APP/Aβ in the pyramidal cell layer of the 3xTg-AD host hippocampi. Notably, also some cells of the transplanted tissues showed positive immunoreactivity with this antibody (Fig. 3). However, neither the host nor the graft developed an extracellular amyloid deposition.
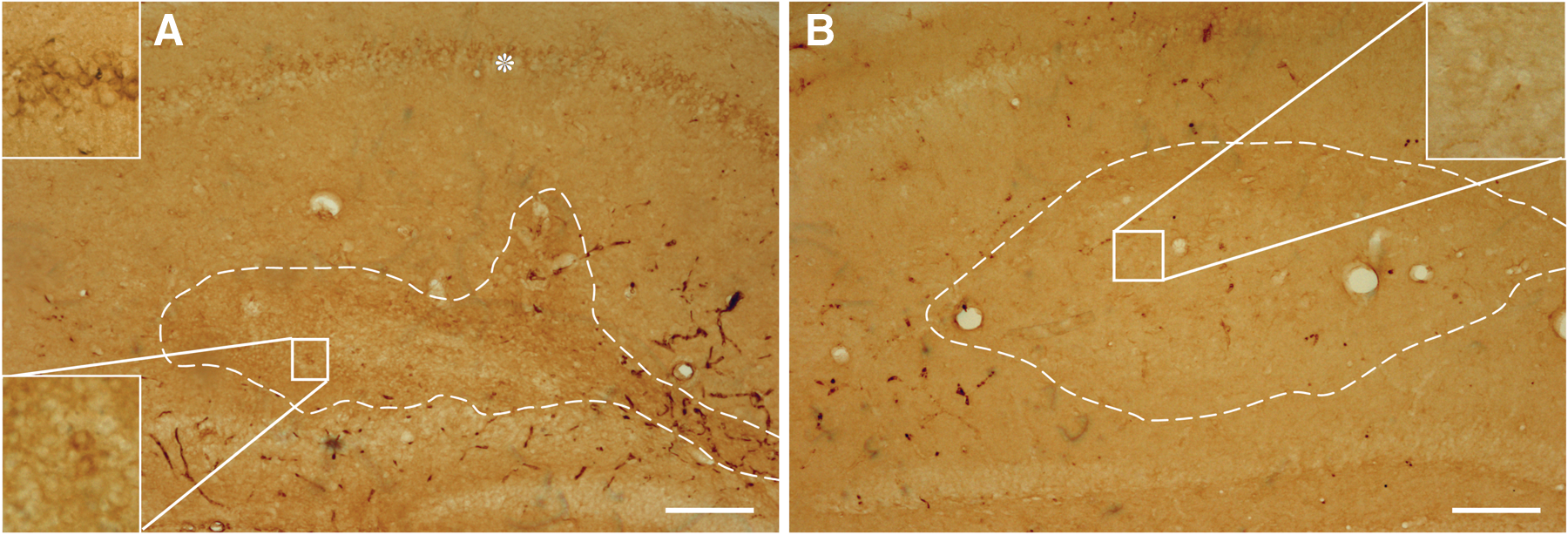
Intracellular APP/Aβ deposition in WT tissue grafted in 3xTg-AD mice 3 months post-transplantation. Representative images of coronal hippocampal sections of 9-month-old 3xTg-AD (
Graft processes inside host tissue
Numerous processes emanating from the graft and elongating inside the host formed synaptic contacts and dendritic spines, indicating integration and maturation of the transplanted tissues within the hippocampal region (Fig. 1B–E).
Neurites total number
The processes extending from the graft into the host tissue were analyzed using Neurolucida and Imaris software. The number of neurites and the cumulative length of all neurites associated with grafts were calculated for each condition. The tissues grafted in the affected hosts (3xTg-AD) presented a significant decrease (<35.38%; ANOVA one-way test p < 0.05) in total number of their neurites when compared to the grafts in normal SVBL6 control animals (Fig. 4). Similarly, the migration area of the neurites in the 3xTg-AD host, defined as the ratio of the area occupied by neurites to the total graft bulk surface, was reduced by 59.35% compared to the neurites in the control condition (p < 0.05) (Fig. 4).
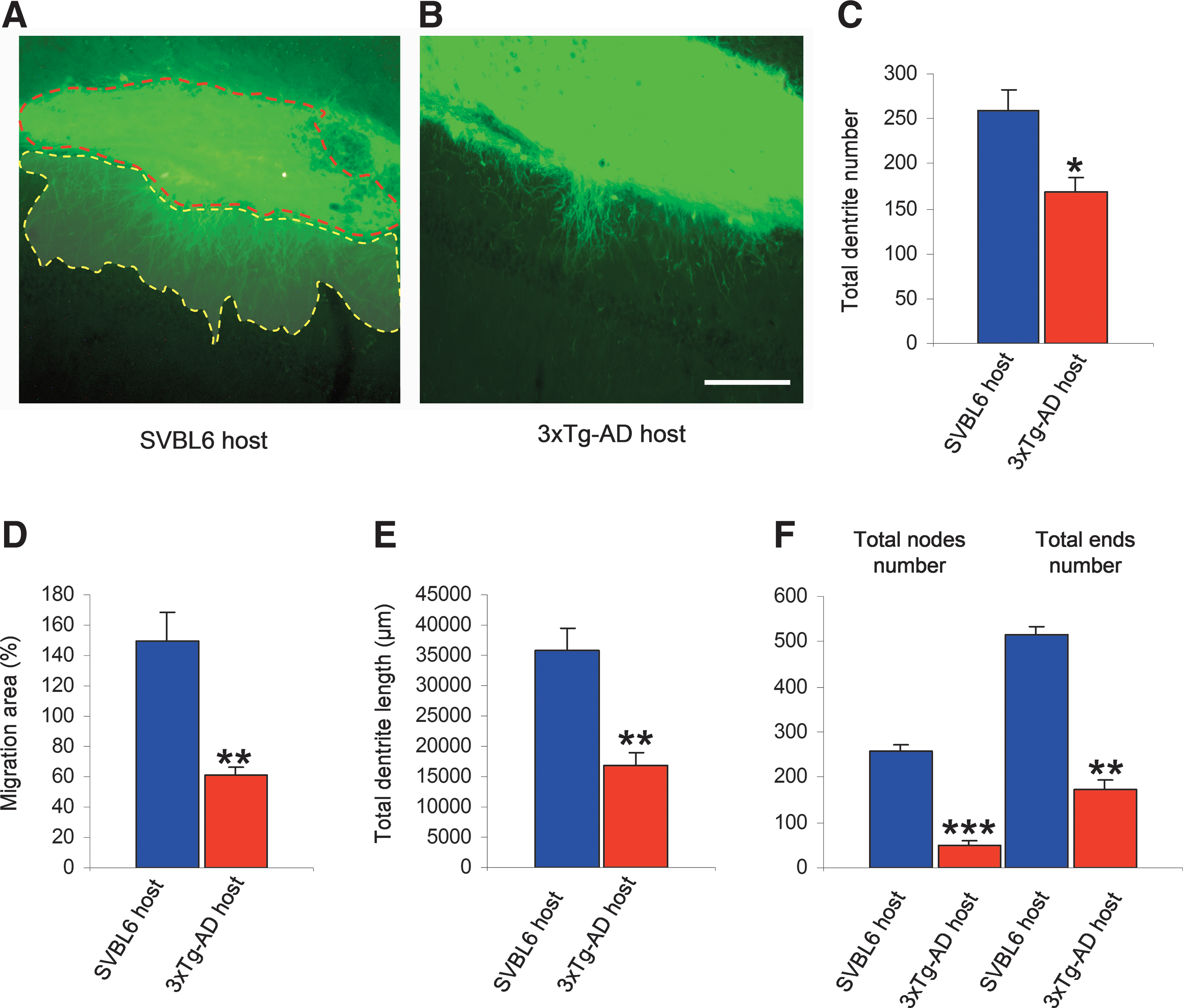
Neurite outgrowth from the transplant is reduced in 3xTg-AD mice. Representative images of wild-type (WT) transplant's neurites in WT control host (
Neurites total length and branching
The measurement of cumulative and average neurite length revealed that the tissue transplanted in WT mice was more effective in increasing the length of its neurites (53%; p < 0.05) in comparison to the graft in the AD mouse model 3xTg-AD (Fig. 4). Moreover, in an attempt to explore the negative impact of the affected host on neuronal differentiation, we examined the morphology of neurites emanating from each graft. We observed that the transplanted tissue suppressed the neuritic branching in 3xTg-AD host by 75.54%, and reduced the ending points by 63.34%, compared to SVBL6 WT host.
Spine density and morphometric analysis
At high magnification, a huge number of grafts processes displayed dendritic spines. To determine the degree of maturation of such dendrites of the transplanted tissue, we analyzed the density of dendritic spines inside WT and transgenic mice hosts. A total number of 107–135 neurites from different grafts in each environmental condition were included in final analysis of dendritic spine density and spine morphology using NeuronStudio software. The density was calculated by dividing the total number of spines by the dendritic length of the segment. In 3xTg-AD mice, the spine density was reduced by 41.34% (p < 0.05) compared with SVBL6 WT hosts. Moreover, in the 3xTg-AD host, some abnormal varicosities were detected that were not present in SVBL6 controls (Fig. 5).
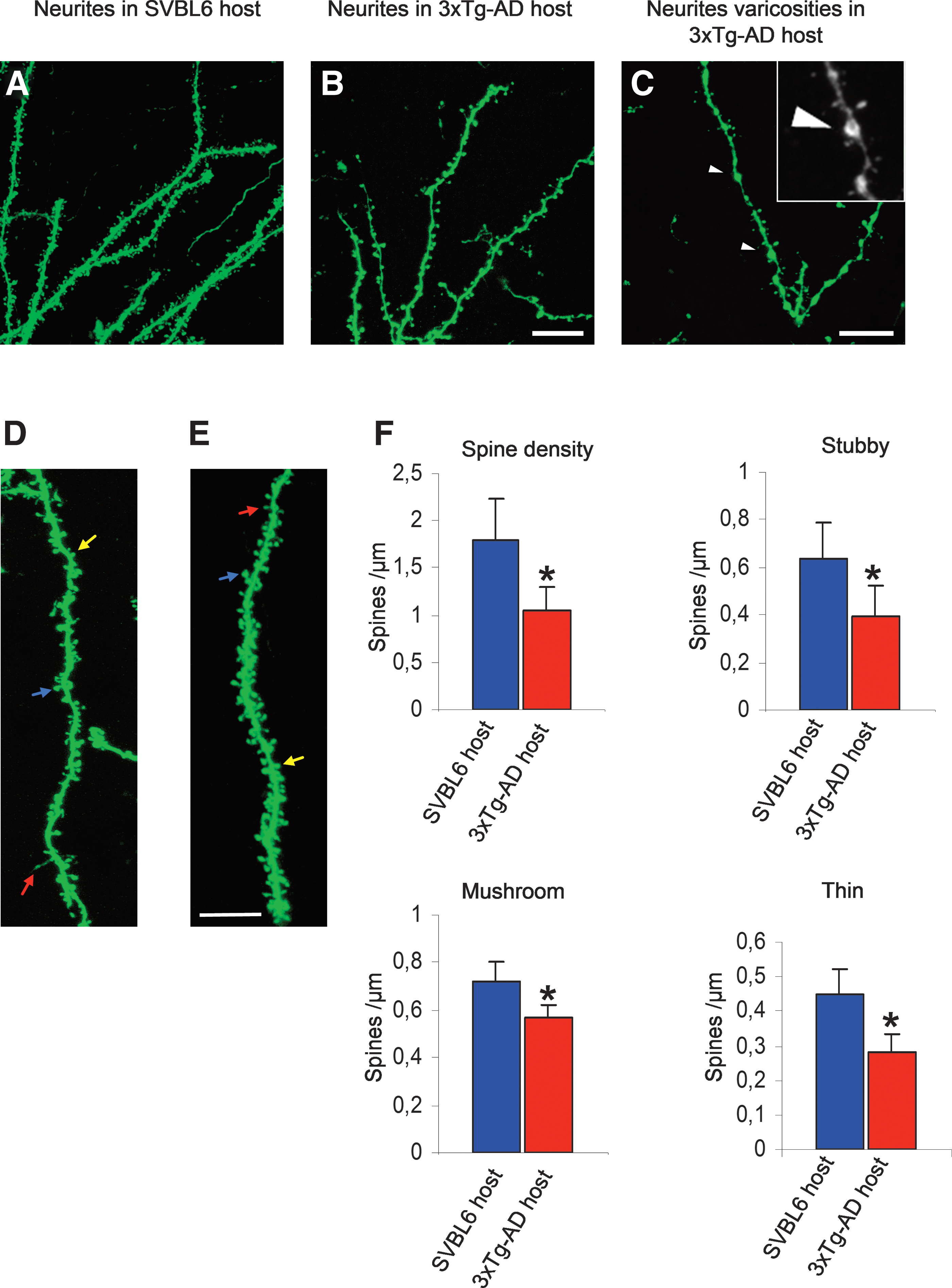
Analysis of dendritic spine density in the CA1 hippocampal region. Representative confocal images of wild-type (WT) dendrites (green fluorescent protein [GFP], green) extending and forming spines in the nontransgenic SVBL6 host (
Dendritic spines were then classified in three classes on the basis of their relative size of head and neck 22 —thin spines with a small head and narrow neck (head diameter <0.4 μm), stubby spines with no neck constriction, and mushroom spines characterized by a constricted narrow neck and a large irregular head (maximum spine head diameter >0.4 μm). All spine measurements were performed in three-dimensional reconstructions from z-stacks. Statistical analysis indicated a significant decrease in the three main spine classes within the 3xTg-AD mice hosts compared to the SVBL6 control mice (Fig. 5). This reduction in density was noticed particularly in stubby spines, which manifested a decline of 38.09% (p < 0.05) and in thin spines (36.36% reduction; p < 0.05), whereas the mushroom spines showed a smaller but significant reduction of 19.71% (p < 0.01) (Fig. 5).
Glial reaction and neuroinflammation
To examine whether the impairment in dendritic morphology maturation of the transplanted tissue was associated with a host and/or graft glial response, GFAP and Iba1 immunostainings were performed to identify astrocytes and microglia respectively. Three months after transplantation, all of the grafts appeared to be surrounded by a bundle of astroglial cells, indicative of an immune response of the host. The analysis by microscopy of the GFAP-positive astroglial profiles showed a substantial astrocytic reaction in both transgenic AD animals (3xTg-AD) and in WT control hosts (Fig. 6). However, we found that within the graft less than 7% of these GFAP-positive astroglial cells expressed GFP, indicating that the majority of these astrocytes were derived from the hosts. In contrast to the astrocytic reaction surrounding the transplanted tissue, inside the grafted tissue the overall density of GFAP-positive astroglial cells showed no significant difference in the 3xTg-AD hippocampus (8.9% increase; p > 0.05) compared to the transplant in normal control hosts. Nevertheless, in several astrocytes within the grafts transplanted in 3xTg-AD mice, we noticed a reduction in branching, which is indicative of the process of atrophy (Fig. 6).
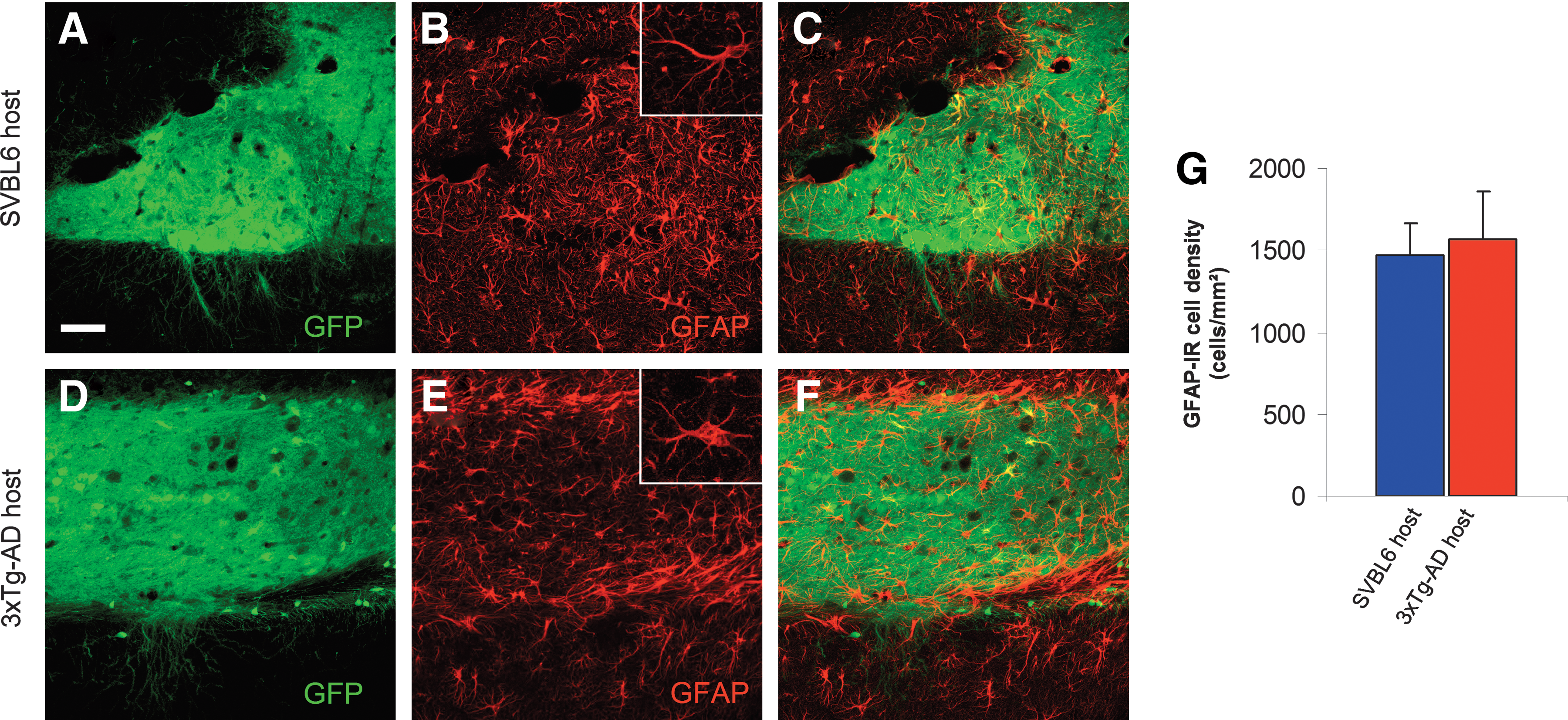
Astroglial reaction after intrahippocampal transplantation. Confocal images illustrating the hippocampal graft tissue in SVBL6 wild-type (WT) host (
On the other hand, the visualization of microglia with anti-Iba1 revealed a significant increase (40.6%; p < 0.05) in the cell density of immunoreactive microglia within the tissues transplanted in 3xTg-AD mice compared to those in WT control mice. This increase is manifested by activated microglia with cytoplasmic hypertrophy (Fig. 7).
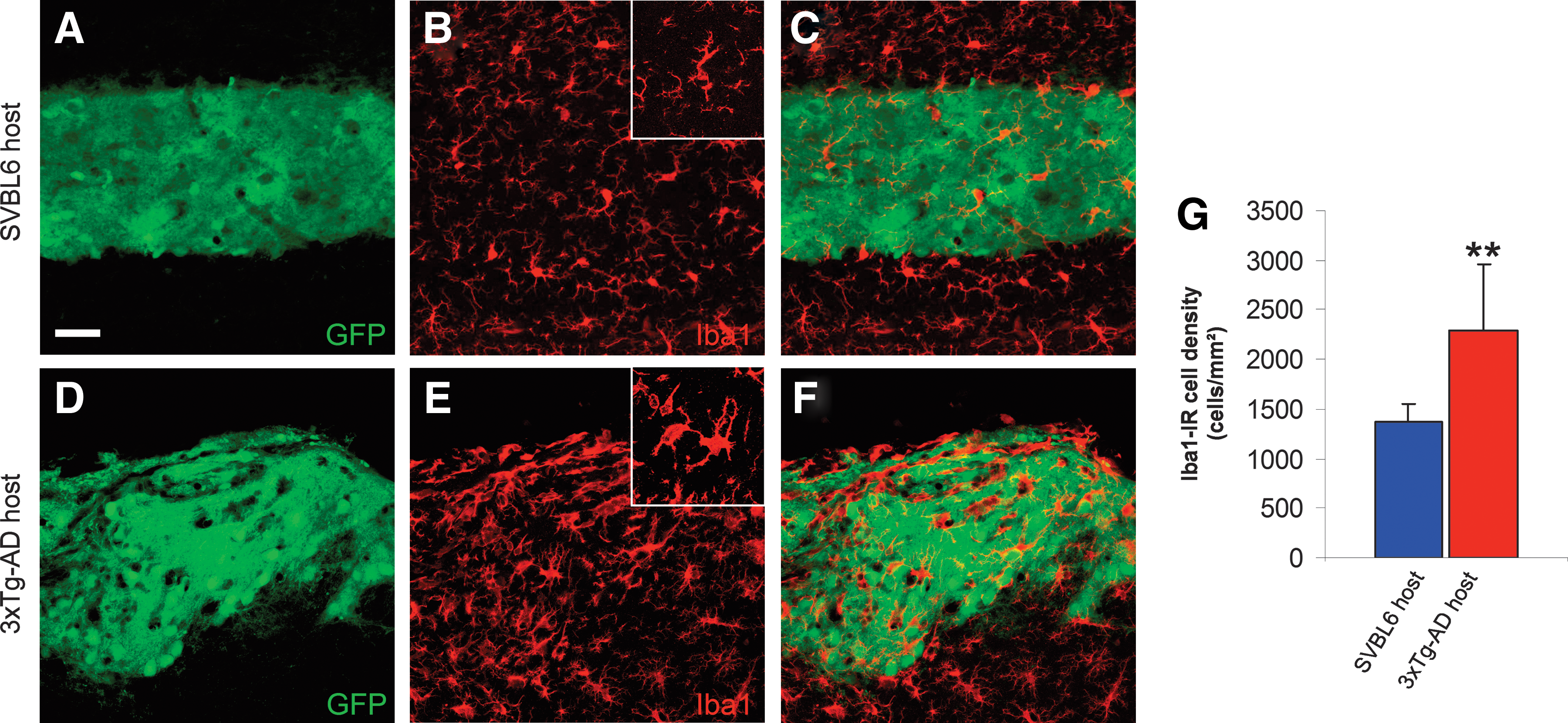
Activated microglia after intrahippocampal transplantation. Representative confocal images illustrating the hippocampal graft tissue 3 months post-transplantation in SVBL6 wild-type (WT) host (
Discussion
In this study, we propose a new methodology to study the mechanisms underlying the propagation of AD from affected to healthy tissue. Because several studies presented the small soluble Aβ oligomers as the important neurotoxic forms in AD, 5,23 we investigated the behavior and the fate of embryonic WT tissue transplanted into the hippocampus of either a WT host or a triple transgenic mouse 3xTg-AD harboring mutations for APP (Swedish), presenilin PS1 (M146V), and Tau (P301L) at an age when only soluble Aβ is abundant. In fact, selecting this period for hosting the transplant allowed us to rule out any other pathological aspect, such as amyloid plaques and neurofibrillary tangles, present in this animal model, hence studying separately the effects of soluble Aβ in the propagation of the pathology.
We used solid pieces of tissue as transplants to preserve the intrinsic organization of the graft that may facilitate the detection of the effects of extrinsic factors of the new environment on the post-transplantation development of grafts. 16 Our histological and morphological analyses indicated that grafts of fetal hippocampal tissue survive transplantation in both WT and 3xTg-AD hippocampus and are also establishing connections with the hosts. Our data indicate that hosting a WT tissue for 3 months in a 3xTg-AD mouse affected by soluble Aβ causes a reduction of the neurite outgrowth of the transplant when compared to the graft in the normal condition. This reduction is manifested by the decrease of total number of neurites, neurites length, and dendritic arborization. This observation supports the notion that APP and/or Aβ produced by the transgenes had a detrimental effect on neurite outgrowth. 24 –26
Interestingly, we found that some WT cells of the tissue transplanted in the transgenic mouse contain intracellular deposits of Aβ/APP, suggesting the movement and the diffusion of amyloid peptide from the affected host to previously healthy neurons. Moreover, although this intracellular accumulation of Aβ/APP is not abundant in the cells of the graft, it may also contribute and explain some of the previously observed alterations in neurites outgrowth. These results support the idea that the toxic environment within the 3xTg-AD brain tissue is the major cause leading to dendritic pathology in healthy neurites, probably by the means of soluble Aβ, without requiring the transfer of APP/Aβ species into the graft tissue.
Subsequently we asked whether the transgenic host can also affect the structural plasticity of the transplant's neurites. Because dendritic spines represent the main sites of excitatory synaptic transmission, we analyzed the spine density of the dendrites emanating from grafts in each environment using NeuronStudio software. A previous study, aimed at analyzing the kinetics of dendritic spine loss within the somatosensory cortex in 3xTg-AD mice, by means of long-term two-photon in vivo imaging, has shown that dendritic spine loss did not occur in this region before 13 months of age. 27 Thus, the deficiency of spines that we observe in grafted neurons occurs earlier and is more severe than in the 3xTg-AD host. This suggests that the development of new spines in grafted cells is more severely affected than the already formed and stabilized spines of the host. However, other studies reported exaggerated endoplasmic reticulum Ca2+ signals, mediated by the ryanodine receptor, in hippocampal neurons of young 3xTg-AD mice (4 months old) compared to age-matched WT animals, resulting in spine and synapse loss in the CA3 and CA1 regions of the hippocampus. 28 –30
Our experiments provide evidence that the high level of soluble Aβ produced in the host may lead to the propagation of pathophysiology AD features to the neighboring healthy tissues by decreasing spine density compared to the control condition. Indeed, several studies reported that elevated levels of soluble Aβ contributed to cognitive deficits and synaptic loss in AD patients. 31,32 In addition, some in vitro studies demonstrated that high levels of soluble Aβ directly lead to synaptic loss. However, besides the accumulation of soluble Aβ reaching its highest level at 6 months in 3xTg-AD, 33 the presence of intracellular Aβ/APP in some cells of the graft may contribute also to this loss of spine density. This idea has been further corroborated by a recent study, which demonstrated that dendritic spine loss in a transgenic mice model can take place either with the occurrence of intracellular soluble or extracellular fibrillar Aβ alone, or the combination of intracellular soluble Aβ and hyper-phosphorylated tau. 27,34
To examine whether this influence on synaptic connections is confined to a specific spine shape, we classified the spines in three classes to analyze their morphology. Although a significant decrease appeared in the spine density for the three different shapes, it is remarkable that the impairment of mushroom spines was smaller (19.7%) compared to the stubby (38.1%) or thin spines (36.4%). Previous work proposed the mushroom-shaped spines as the most stable and mature spines that mediate strong synapses in mature hippocampus. 35 –37 In addition, the development of neurons required an increase of thin and mushroom spines and decrease of stubby spines. 38
Therefore, relying on these data, we hypothesize that the high level of soluble Aβ abundant in the host environment affected the synaptic connections of graft neurites by preventing principally the formation of new immature spines that can transit during development to stable mushroom spines and in second place the deletion of some early sprouted mushroom spines. However, even if suggestive, this hypothesis needs further investigation to be clarified. On the other hand, the results reported here do not rule out other related mechanisms by which the synaptic plasticity could be affected, like the calcium-overload hypothesis supported by several studies. 28 –30
The inflammatory reaction of the 3xTg-AD and WT hosts against the transplanted tissues was confirmed by the immunohistochemical examination of reactive astrocytes and microglia. The overall density of GFAP-positive astroglial cells inside the grafts did not change significantly, in agreement with previous studies. 39,40 However, we identified some astrocytes displaying evidence for atrophic features in the tissue transplanted within the 3xTg-AD hippocampus. In contrast, microglial cells with cytoplasmic hypertrophy were observed in the affected host tissue environment but not in the control, suggesting a potential contribution of microglial cells in the histopathological features present in the graft. Additionally, it has been shown that one of the microglial roles is to take up solutes and nutrients through endocytosis 41,42 and mediating the clearance of soluble Aβ. 43 The second observation to be considered was the absence of any glial cell migration from the graft toward the two types of host (3xTg-AD and WT) and only a massive accumulation around the interface between the graft and the host. This observation is in contrast to other studies demonstrating that glial and endothelial cells derived from solid hippocampal tissue (which were maintained in intermediate explant culture for a few days prior to transplantation for labeling with tritiated thymidine) showed a migration into the host. 44,45 This controversial observation could be explained, at least in part, by the difference between the nature of the graft used in our study and those passing through a cell culture prior to transplantation.
In conclusion, our results show that the propagation of AD pathology can occur at an early stage of the amyloid cascade, probably by means of soluble Aβ, with initial disruption of neurites morphology and spine density. Moreover, the intracellular APP/Aβ shows a spreading and diffusion to healthy tissues, in line with the hypothesis of a prion-like mechanism of propagation. Finally, the validation of this methodological approach will provide a suitable animal model to understand and uncover the mechanisms underlying the spreading of AD to the surrounding healthy regions.
Footnotes
Acknowledgments
The experiments were supported by a grant from the University of Torino for a research on “Integration of healthy embryonic nervous tissue in the adult brain of mice models of Alzheimer's disease.” The authors have no actual or potential conflict to disclose.
Author Disclosure Statement
No competing financial interests exist.