
Abstract
There is a growing interest in the potential of mesenchymal stem cells (MSCs) for implementing regenerative medicine. We assessed the effect of intravenous administration of human bone marrow-derived MSC on the life span of a single Sprague-Dawley female rat. The treatment was started when the rat was 6 months old and the cells were administered every 2 weeks afterward. The treatment did not induce any obvious changes in body growth or behavior and the rat showed the typical age changes for this strain, except that, unlike intact counterparts, the animal did not develop mammary tumors or pituitary gland hyperplasia. The more remarkable effect of the treatment was on life span, which was 44 months compared with an average of 36 months for intact laboratory rats. We conclude that despite the low N value, it is likely that the MSC treatment was responsible for the exceptionally long survival of the rat. The potential rewards of confirming the present findings warrant further studies involving higher N values.
Introduction
M
There is a growing interest in the therapeutic potential of MSC; thus, administration of MSC in different animal models was found to have restorative effects in liver fibrosis, 6 myocardial infarction, 7 and Alzheimer's disease (AD). 8 There is also evidence that with age, the pool of MSCs becomes progressively smaller and the remaining cells lose vitality, a fact that led one of us (E.M.) to hypothesize that there is an age-related stem cell exhaustion syndrome, which may be responsible for a number of chronic pathologies, including aging itself. 9 In this study, we report that long-term intravenous administration of human bone marrow-derived MSCs in a single female rat was associated with a remarkably long survival, 44 months, compared with an average life span of 36 months for most albino rats.
Materials and Methods
Animals
Sprague-Dawley rats are raised in our rat colony at INIBIOLP. They are housed in a temperature-controlled room (22 ± 2°C) on a 14:10-hour light/dark cycle. Food and water are available ad libitum. In our rat colony, the average 50% survival time for females, studied in groups of 50–60 animals, is ∼32 months, provided mammary tumors are systematically removed when detected by palpation. The rat under treatment belonged to a group of 50 virgin females to be used in the future when they were between 27 and 29 months old. The rat was placed alone in a standard stainless steel square cage. Typically, our female rats show an almost flat survival curve (100% alive) until age 24–26 months when they begin to die in progressively increasing numbers. All experiments with animals were performed in accordance with the Animal Welfare Guidelines of NIH (INIBIOLP's Animal Welfare Assurance No A5647-01). IACUC approved on April 3, 2007, and extended on December 19, 2011.
Human bone marrow-derived MSC isolation and preparation
Ten milliliters of heparinized bone marrow blood was sterilely obtained by needle aspiration from the ileac bone of a male (56 years old) cadaver donor, in a routine procurement procedure at the Buenos Aires Province Ministry of Health Transplantation Agency, CUCAIBA, after cardiac arrest and when all other organs and tissues suitable for transplantation had been already collected. In this procurement procedure, the donor's bone marrow blood was collected after fulfilling all requirements as needed by law, for organ, tissue, and cell transplantation and research. The bone marrow blood was transported and processed under sterile conditions. Briefly, blood was diluted in low-glucose Dulbecco's modified Eagle's medium (DMEM; Invitrogen), and mononuclear cells collected after density gradient centrifugation and cultured for the isolation of MSC in 75-cm2 tissue culture flasks at 37°C in a CO2 incubator (5% CO2) at 95% humidity with 10% of selected fetal bovine serum (FBS; Invitrogen). Cells were cultured overnight and the adherent cells were allowed to attach to the plastic flasks. Nonadherent cells were removed and the culture expanded for several days, feeding them every 3 days. MSCs appeared soon as typical round colonies and a monolayer started to form over 14–17 days in every flask. When the cells reached 80% confluence, they were detached from the culture flasks, expanded in a second passage, phenotyped as CD105-, CD73-, CD44-, and CD90-positive cells, lacking CD45, CD34, CD14, CD11b, CD79, CD19, and HLA-DR surface molecule expression, microbiologically tested, and karyotyped to confirm they were normal. The MSCs were then placed in low-glucose DMEM with 20% FBS and 10% dimethyl sulfoxide (DMSO) and cryopreserved in liquid nitrogen. This preparation was the source of MSCs for the whole study. For each injection, a cell aliquot was thawed and cultured, with regular examination under a phase-contrast inverted microscope. Finally, the MSCs were detached, concentrated by centrifugation, their concentration properly adjusted, and delivered to the rat as described below.
Treatment with MSC
A suspension of MSC in low-glucose DMEM (5 × 106 cells in 0.5 mL) was injected with a tuberculin syringe fitted to a 25G needle in one of the tail veins. The treatment was started at 6 months of age and the MSC injections were administered every 2 weeks until the end of the study.
Results
The MSC injections did not cause any observable changes in the rat, either beneficial or detrimental. As the animal aged, it progressively developed bilateral cataracts that became severe after 30 months of age. This pathology was also observed in the majority of the intact counterparts. Although there is a high incidence of nonmetastatic mammary tumors in our aging females (which are surgically removed shortly after detection to prolong the animals' life), none was detected in the treated rat. The body weight of the experimental rat showed a slight increase as it aged. It was 291 g at 27 months, 314 g at 36 months, and 321 g at 42 months. Taken as a reference, fifteen 27-month intact counterparts weighed (X ± SEM) 287 ± 4 g. At 30 months of age, a 7-day-long assessment of vaginal smears revealed that the animal was, as expected, in constant anestrus. At 42 months of age, the treated rat showed a mobility and exploratory activity roughly comparable to that of 26-month-old intact counterparts when placed on the Barnes maze. At this age, the rat showed no paralysis or dragging of the hind legs and displayed a normal posture (Fig. 1). The animal died at 44 months of age, during the night, without having shown overt signs of disease during the previous days. The necropsy revealed no tumors or gross pathology in the major organs. A remarkable finding was the preserved size of the pituitary gland, which is almost invariably enlarged or even tumoral in our old female rats at 27–29 months of age.
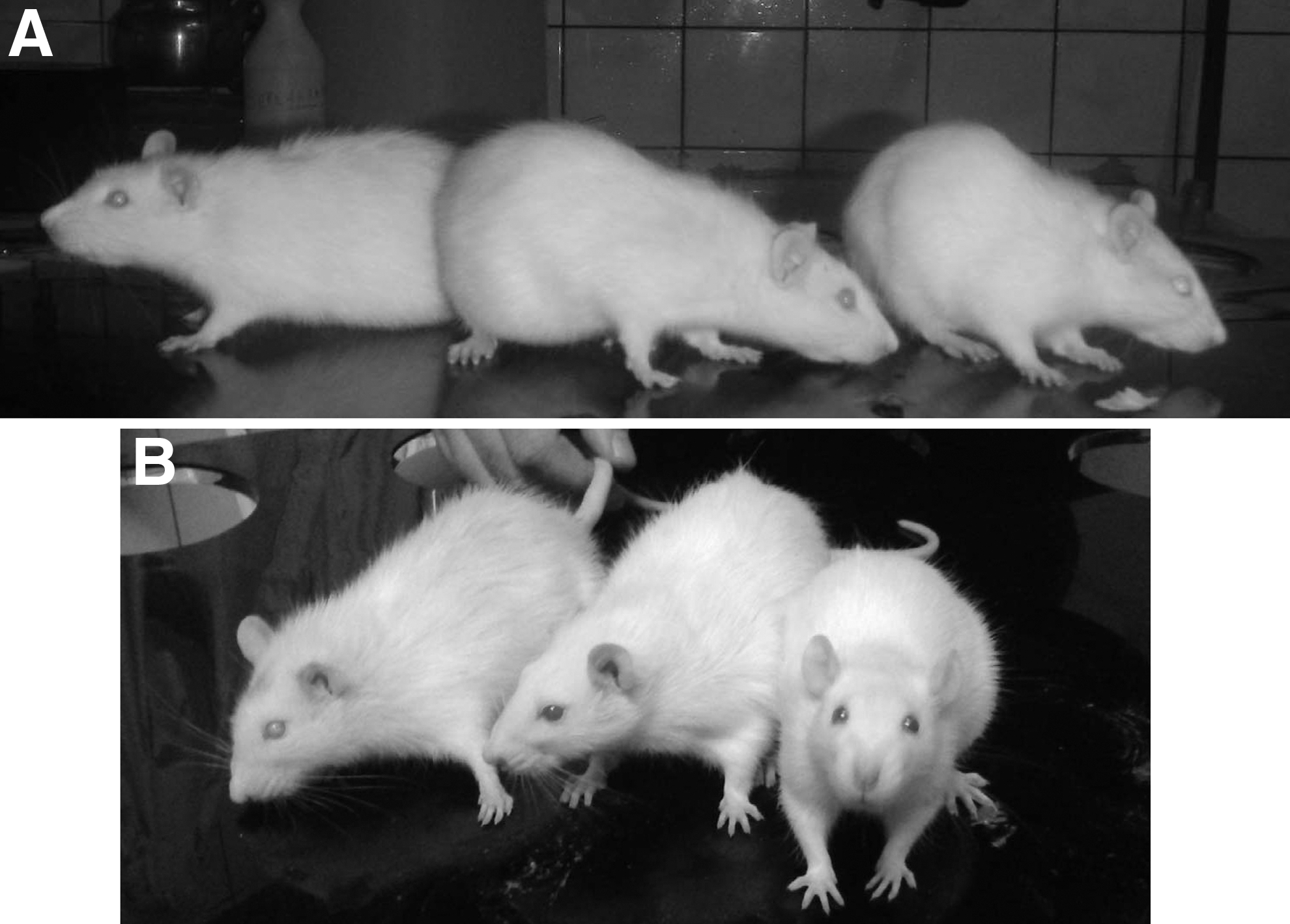
Panel A shows from left to right, the treated rat at 42 months of age, an old rat at 26 months, and a 3-month-old young rat. Panel B shows from left to right, the treated rat at 43 months, a 28-month-old rat, and a 4-month-old young rat. All animals are females. The young and old rats are different animals in
The brain was removed and the remainder of the head was frozen and sent to Dr. Steven Clarke, Department of Chemistry and Biochemistry and the Molecular Biology Institute, University of California at Los Angeles, for an independent determination of age by analysis of racemization and isomerization of teeth and eye lens proteins. The results are reported in this issue. 10
Discussion
The average life span of laboratory rats is 3 years 11 and up to now, calorie restriction remains as the only proven intervention able to substantially extend life span in rodents. It was first demonstrated by the pioneering studies of McCay et al. 12 in rats. The authors used severe caloric restriction started after weaning to retard growth, keeping the body weight of the animals at about half the weight of the ad libitum fed counterparts. In the females, the maximum life span increased from 1189 days (39.6 months) in the fully fed rats to 1421 days (47.4 months) in the calorie restricted females after weaning. The mortality was high as reflected by the fact that the average life span of the surviving females was 801 (26.7 months) and 775 (25.8 months) days for control and calorie restricted females, respectively. Comparable results were recorded in males. In young female Sprague-Dawley rats, it was shown that 50% food restriction induces a severe panhypopituitarism as well as an arrest in ovulation and estrous cyclicity. Upon refeedeing, the rats resumed estrous cycles and remained cycling for longer than their full-fed counterparts. 13
Unlike calorie restricted rats, the MSC-treated rat gained weight at the same rate as the untreated counterparts and did not show any signs of disease or behavioral abnormalities (severely calorie restricted rats show profound behavioral changes, including cannibalism). Mammary tumors and pituitary hyperplasia or adenoma are the two most frequent pathologies in aging Sprague-Dawley female rats. 14 Interestingly, our experimental rat did not develop mammary tumors during its extended life nor was its pituitary gland enlarged at the time of death.
The fact that we used a single experimental animal limits the strength of the conclusions that can be drawn. Nevertheless, it should be pointed out that it is highly unlikely that, although possible, the rat chosen at random for the treatment was one of the extremely rare very long-lived individuals that exist in a colony of Sprague-Dawley rats. Interestingly, heterochronic parabiosis has been reported to extend the life span of the older member of the parabiotic pair. 15 The intervention extended the maximum life span of Buffalo female rats from 1100–1150 days (37 months) in single intact females to 1150–1200 days (39 months) in the older member of the heterochronic parabiotic pair. Interestingly, Shen et al. 16 reported that bone marrow-derived MSCs from young, but not from old, mice implanted in full body X-irradiated old (18–24 months) female mice, moderately but significantly increased their life span and slowed their age-related loss of bone density. Furthermore, a recent study in F344 male rats showed that long-term intravenous treatment (one injection a month) of the animals with human amniotic membrane-derived mesenchymal stem cells (AMMSCs) or adipose tissue-derived mesenchymal stem cells (ADMSCs) extended their maximum life span from 23 months (saline-injected controls) to 30 and 34 months, respectively. The treatment, which did not elicit a significant immune rejection, also improved the increased concentration of acetylcholine and recovered neurotrophic factors in the brain and muscles, leading to restoration of microtubule-associated protein 2, cholinergic and dopaminergic nervous systems, microvessels, muscle mass, and antioxidative capacity. 17 Although these rats were shorter lived than average rat strains (typical life span, 36 months), the results are fully in line with ours and support the hypothesis that stem cell depletion is relevant to the process of systemic aging.
Another study suggests that the plasma from young mice contains molecules able to improve cognitive functions in old mice. 18 Taken together, the above studies are consistent with the hypothesis of a stem cell exhaustion syndrome as a relevant (although not exclusive) cause of aging and chronic pathologies. 9 The heterologous MSCs used in the present study came from a patient approaching old age, yet the treated rat had a life span 22% longer than the average life span of laboratory rats. Since this is, to our knowledge, the first report of a life extending activity of MSC in rats, it would be premature to postulate a mechanism for this effect. Clearly, the tolerance of the rat to these heterologous MSCs was very high. Importantly, preliminary experiments, in which we intracerebroventricularly administered human MSCs to 27-month-old female rats or streptozotocin-lesioned young males (an AD model), revealed only a mild restorative effect on spatial memory. This suggests that MSC therapy must be started during adulthood or youth to significantly extend life and delay the onset of age-related pathologies.
Concluding Remarks
This is a 4.5-year-long experiment with a design weakness that stems from the minimal N value used, a limitation that is at least partly counterbalanced by the high unlikelihood of finding by chance such a long-lived individual in our rat colony. Clearly, the results are not conclusive but they invite further exploration. We believe that the potential rewards of confirming the present findings warrant investing the time and resources needed to replicate the study with a higher N value.
Footnotes
Acknowledgments
The authors are grateful to Dr. Aubrey D.N.J. de Grey of the SENS Research Foundation (Mountain View, CA) for useful comments on the data and for his active role in promoting complementary collaborative studies of our rat material. R.G.G. also thanks Mr. Ben Best, former Cryonics Institute president, for stimulating discussions on life extension strategies and for assisting us to disseminate the present results. This study was supported, in part, by grant # R01AG029798 from the Fogarty International Center and the National Institute on Aging, NIH, to R.G.G., and by funding from the Buenos Aires Ministry of Public Health to E.M.
E.M. designed the study and monitored the preparation of the MSC used. R.G.G. codesigned the study and supervised the animal experiments. G.R. performed the MSC preparative procedures. Y.E.S. performed MSC injections, monitored animal health, and was the main experimenter in charge of handling the treated rat. A.T. provided technical advice and support in the human stem cell procurement process. R.G.G. and E.M. wrote the article. All authors read and approved the article.
Author Disclosure Statement
R.G.G is a career scientist of the Argentine Research Council (CONICET). E.M. is head of the Tissue Engineering, Regenerative Medicine and Cell Therapies Laboratory, CUCAIBA, Buenos Aires Province Ministry of Public Health, Argentina. G.R. is a senior technician at CUCAIBA. Y.E.S. is a CONICET senior research professional. A.T. is a senior clinician at CUCAIBA.