
Abstract
Umbilical cord mesenchymal stem cells (UCMSCs) have been identified as a potentially ideal cell type for use in regenerative therapeutic contexts owing to their excellent paracrine secretory abilities and other desirable properties. Previous work has shown that stem cell-derived exosomes can effectively reduce skin aging, but few studies have specifically focused on the role of UCMSC-derived exosomes in this context. In this study, we isolated exosomes derived from UCMSCs grown in a three-dimensional culture system and explored their ability to modulate the photo-aging of HaCaT keratinocytes. Cell viability and proliferation were assessed using CCK8 assay, whereas wound healing and transwell assays were used to assess cell migratory capabilities. UVB irradiation (60 mJ/cm2) was used to induce photo-aging of HaCaT cells. TUNEL and SA-β-Gal staining were used to explore HaCaT cell apoptosis and senescence, respectively, whereas real-time quantitative PCR was used to assess the expression of relevant genes at the mRNA level. We found that UCMSC-derived exosomes were able to enhance normal HaCaT cell proliferation and migration while also inhibiting UVB-induced damage to these cells. These exosomes also reduced HaCaT cell apoptosis and senescence, increasing collagen type I expression and reducing matrix metalloproteinase (MMP1) expression in photo-aged HaCaT cells. Together, these findings indicate that UCMSC-derived exosomes have the potential to be used therapeutically to suppress skin aging.
Introduction
The aging of skin occurs owing to a confluence of exogenous and endogenous aging process, with chronological- and photo-aging being primary drivers of these process. 1 Intrinsic chronological aging at present is inevitable and results from genetic and physiological processes, whereas extrinsic aging leads to the development of morphological changes such as reduced skin elasticity, increased skin roughness, and the development of wrinkles. This process is characterized by reductions in basal collagen type IV and collagen type VII levels in the skin as a consequence of environmental exposure to pollution, smoke, or ultraviolet (UV) radiation. 2 UV radiation derived from sunlight exposure is the primary driver of photo-aging, with UVA (320–400 nm), UVB (280–320 nm), and UVC (200–280 nm) forms of radiation all contributing to this process. 3 As UVB and UVC radiation can easily penetrate the atmosphere of the Earth, they are the primary drivers of epidermal damage and associated skin damage.
UVB rays are important for facilitating antimicrobial peptide production and vitamin D absorption, but as they are more energetic than UVA rays they are better able to induce DNA damage in epidermal cells. 4 UVA radiation can, in contrast, penetrate to deeper layers of the dermis and cause photo-aging over extended periods of exposure. The keratinocytes composing of the epidermis serve as a barrier that prevents environmental pathogens and irritants from entering into the body. These cells are organized in the form of a stratified squamous epithelium, with keratinocyte stratum corneum formation being important for protecting against extrinsic damage. 5 Absorption of light by the epidermis can lead to damage to nucleic acids and amino acids, in addition to the generation of harmful free radicals. 6
At present, the treatment and prevention of skin aging relies upon both surgical approaches such as wrinkle removal and tissue filling, as well as nonsurgical interventions including laser therapy and chemical exfoliation. 7 There are, however, significant disadvantages to these forms of treatment, leading many researchers to focus on mesenchymal stem cells (MSCs) as a potentially ideal means of mediating epidermal regeneration. MSC transplantation has been shown to be an effective means of treating skin, with the secretory functions of these cells being particularly important for their therapeutic efficacy. Recent studies have further explored the value of MSC-derived exosomes as key mediators of paracrine therapeutic activity. 8,9 Cell-based therapies offer certain drawbacks that make cell-derived exosomes more ideal for most therapeutic applications. Exosomes are small (30–200 nm) lipid vesicles that can enclose a range of molecule types including cytokines and growth factors, other proteins, as well as mRNAs, micro RNAs (miRNAs) and long noncoding RNAs (lncRNAs). 10,11 These exosomes can transmit these molecules between cells, thereby facilitating intercellular communication. Previous work has shown that umbilical cord MSC (UCMSC)-conditioned media can stimulate keratinocyte proliferation. 12,13
Based on these findings, we hypothesized that UCMSC-derived exosomes may be key mediators of photo-aging recovery. To confirm whether these UCMSC-Exo were able to reduce UVB-mediated damage to HaCaT cells, we explored their impact on the proliferative and migratory activity of these cells under control and UVB irradiation conditions. We further explored the ability of these exosomes to protect HaCaT cells against UVB-induced senescence and apoptotic death. Finally, we explored the ability of these UCMSC-derived exosomes to modulate the expression of genes important for extracellular matrix (ECM) repair after UVB irradiation.
Materials and Methods
Cell culture
We obtained human UCMSCs as in our past studies, characterizing these cells based on the expression of defined surface markers and morphological characteristics. 14 HaCaT cells were from the Kunming Institute of Zoology, and were grown using Dulbecco's modification of Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) and penicillin/streptomycin (Thermo Fisher Scientific, Waltham, MA, USA) in a 37°C 5% CO2 incubator (Thermo Fisher Scientific, Waltham, MA, USA). To grow cells using a three-dimensional (3D) culture system, 3 × 108 UCMSCs in α-MEM containing FBS were added to a 3D Fibercell System (Fibercell Systems, NY).
Purification, identification, labeling, and treatment of UCMSCs-Exo
UCMSCs were grown in the logarithmic phase according to the glucose concentration consumed by the culture medium, at which time supernatants were removed and replaced with serum-free media, after which exosomes were collected by classic ultracentrifugation through TheryC's studies. 15 The first step was designed to eliminate cells, large dead cells and large cell debris at 300 and 2000 g for 20 minutes, respectively. The following step was to centrifuge again at 10,000 g for 30 minutes to remove small cell debris. At each of these steps, the pellet was thrown away, and the supernatant was used for the following step. The final supernatant was then ultracentrifuged at 120,000 g to pellet the small vesicles and protein molecules that correspond to exosomes. The pellet was washed in a large volume of phosphate-buffered saline (PBS) to eliminate contaminating proteins and centrifuged one last time at the same high speed. 15 After isolating these exosomes, they were resuspended in PBS and stored at −80°C. A nanoparticle tracking analysis (NTA) was used to assess exosome concentrations and particle size with zetaview PMX 110 (Particle Metrix, Meerbusch, Germany). In brief, exosome suspensions were injected into the laser chamber, with Brownian motion over 30 seconds being recorded three times. The ZetaView 8.04.02 software was then used to assess exosome size and electrical potential. Exosome morphology was assessed using transmission electron microscopy (TEM; JEM-1010; JEOL, Tokyo, Japan). Exosomes were initially adsorbed onto a copper mesh for 10 minutes, after which 2% paraformaldehyde was used to fix cells. TEM analysis was conducted by staining these cells for 10 minutes using 2% uranyl acetate and then imaging them with a TEM at 60 kV.
To examine the uptake of UCMSC-Exo by HaCaT cells in vitro, UCMSC-Exo were labeled using a PKH67 green fluorescent labeling kit (MINI67; Sigma-Aldrich, St. Louis, MO). In brief, the exosomes dissolved in 1 mL PBS were mixed with 1 mL Diluent C into an EP tube, and then 4 μL PKH67 and 1 mL Diluent C were mixed and placed into another new EP tube. The reagents in the two EP tubes were again mixed evenly, incubated for 4 minutes, and then added to the serum of the same volume to stop the reaction for 1 minute. Then the exosomes labeled with PKH-26 were re-extracted. After plating HaCaT cells into 24-well plates on coverslips (4 × 105 cells/cm2), the labeled exosomes were then incubated with the HaCaT cells at 37°C for 24 hours. Cells on coverslips were then fixed with methanol, washed using PBS, and stained overnight with rabbit anti-β-tubulin (Proteintech, Rosemont, IL) at 4°C. A secondary goat anti-rabbit IgG (Abcam, Hong Kong, China) was then used to detect the protein, whereas DAPI (Abcam) was used for nuclear staining. A Nikon digital sight DS-Ri1 was used to image cells.
CCK8 assay
Cells were added to a 96-well plate (2 × 105 cells/cm2) for 24 hours, after which they underwent starvation in serum-free media for an additional 24 hours, and then 10 μL CCK8 solution (Dojindo, Kumamoto, Japan) was added per well and cells were incubated for a further 1–4 hours. Absorbance at 450 nm was then quantified with a microplate reader.
Migration assay
A total of 5 × 105 cells were added per well in an insert (ibidi, Martinsried, Germany), and after attachment and growth to 95% confluence a scratch wound was generated, and 20 × 108 exosomes/mL in serum-free DMEM were added to wells for 9 hours. Movement into the wound area was then analyzed using microscopy. In transwell assays, 4 × 104 HaCaT cells were added to upper chambers of transwell plates (Corning, MA), with UCMSC-derived exosomes in serum-free DMEM being added to the lower chambers. Plates were then incubated for 36 hours, after which the number of cells that had migrated across the transwell membrane was quantified by removing cells from the upper chamber and then fixing the remaining cells with 4% paraformaldehyde before staining them using 0.5% crystal violet (Sigma-Aldrich) and counting cells on an inverted microscope (Zeiss, Gottingen, Germany).
Real-time quantitative PCR
An RNeasy mini kit (Qiagen, Pudong, Shanghai, China) was used to extract RNA from treated HaCaT cells, after which a GoScript reverse transcription mix (Promega, WI) was used to reverse transcribe 500 ng RNA per sample. EVA Green Supermix (Bio-Rad, CA) was then used for real-time quantitative PCR, with β-actin used as a normalization control. Primers for MMP1, MMP2, laminin, collagen type I, procollagen type I, and TIMP1 were designed with Primer5 as given in Table 1. The 2−ΔCt method was used to quantify relative gene expression.
The Primers Used in Our Experiment
SA-β-Gal assay
HaCaT cells (2 × 105/well) were added to slides in a 24-well plate for 12 hours, after which 60 mj/cm2 UVB irradiation and UCMSC-derived exosomes treatment (80 × 108 particles/mL) were carried out. Cells were allowed to incubate for 24 hours further, after which they were washed in PBS and stained using an SA-β-Gal staining kit (Cell Signaling Technology, Beverly, MA) based on provided directions. A total of three fields of view per slide were imaged, and ImageJ was used to quantify SA-β-Gal-positive cells.
TUNEL apoptosis assay
HaCaT cells were added to slides in a 24-well plate (2 × 105/well) and subjected to UVB irradiation and exosome treatment as appropriate, after which a TUNNEL kit (meilunbio; Dalian, Liaoning, China) was used to assess cell apoptosis based on provided directions. In brief, cells were rinsed in PBS, fixed for 30 minutes with 4% paraformaldehyde, permeabilized for 3 minutes using 0.25% Triton X-10 in PBS, incubated for 1 hour appropriately, and then prepared by terminal deoxynucleotidyl transferase reaction mix at 37°C. Cells were then washed two more times, slides were sealed, and fluorescence imaging at 10 × magnification (Zeiss) was conducted, with three fields of view per slide being assessed.
Flow cytometry
Cells were added to six-well plates (2 × 105/well) and were treated as appropriate. After 24 hours, cells were stained with Annexin V and PI using a kit (Beyotime, Shanghai, China) based on provided directions before flow cytometric analysis. The identity of exosomes was confirmed by the characteristic surface markers CD9, CD63, CD81 by flow cytometry as well. In brief, exosomes were attached to 4 μm aldehyde/sulfate latex beads by mixing exosomes in beads at room temperature with continuous rotation for at least 15 minutes. Then the reaction was stopped with 100 mM glycine and 2% bovine serum albumin (BSA). After CD9, CD63, CD81 beads (System Biosciences, Palo Alto, CA) were added to bond with exosomes for 2 hours, they were washed with PBS and centrifuged at 10,000 rpm, blocked with 10% BSA with rotation at room temperature. The percent positive bead was calculated by FACSAria III instrument (BD).
Western blotting
Cells were lysed, and lysates were subjected to centrifugation to obtain protein-containing supernatants. A total of 15 μg protein per sample then underwent 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) separation and transfer to a polyvinylidene fluoride membrane (Millipore). Membranes were blocked for 1 hour using 5% skim milk, after which they were incubated overnight at 4°C with appropriate primary antibodies, which contained TSG101, CD8, CD63, CD9, and Calnexin. Blots were then washed thrice using TBST and stained for 1 hour with appropriate secondary antibodies. Finally, protein was detected with a supersensitive ECL luminescence reagent (meilunbio; Dalian, Shandong, China), with a ChemiDoc Touch Imaging system (Bio-Rad) used to develop images. All antibodies used in western blotting were purchased from Proteintech.
UVB irradiation
After plating into 96- or 24-well plates, 20 × 108 UCMSC-Exo/mL in serum-free media were used to treat HaCaT cells for 24 hours, after which cells were washed thrice using PBS and then irradiated with a UVB (315 nm) instrument (Sigma High-tech, Shanghai, China). Total UVB dosage was determined based on both irradiation time and the distance between the plate and the radiation source. After irradiation, cells were resuspended in normal growth media, with exosomes being added as appropriate for an additional 24–48 hours. For TUNNEL staining and apoptosis assays, cells were separated into treated and untreated groups, whereas for RT-PCR cells were separated into nonirradiated, irradiated but exosome untreated, and exosomes-treated groups.
Intracellular reactive oxygen species quantification
An ROS Assay Kit (Solarbio) was used to stain for reactive oxygen species (ROS) activity with the 2,7-dichlorodi-hydrofluorescein diacetate (DCFH-DA) probe based on provided directions. In brief, appropriately treated cells were treated with 10 μM DCFH-DA for 20 minutes at 37°C, after which cells were washed in PBS and analyzed using fluorescence microscope.
Statistical analysis
Data are given as mean ± standard deviation of three independent experiments, with p < 0.05 as the significance threshold. GraphPad Prism 6 was used for all statistical testing, with data being compared using Kruskal–Wallis one-way analysis of variance (ANOVA), two-way ANOVAs, or unpaired t-tests as appropriate.
Results
UCMSC-derived exosome characterization
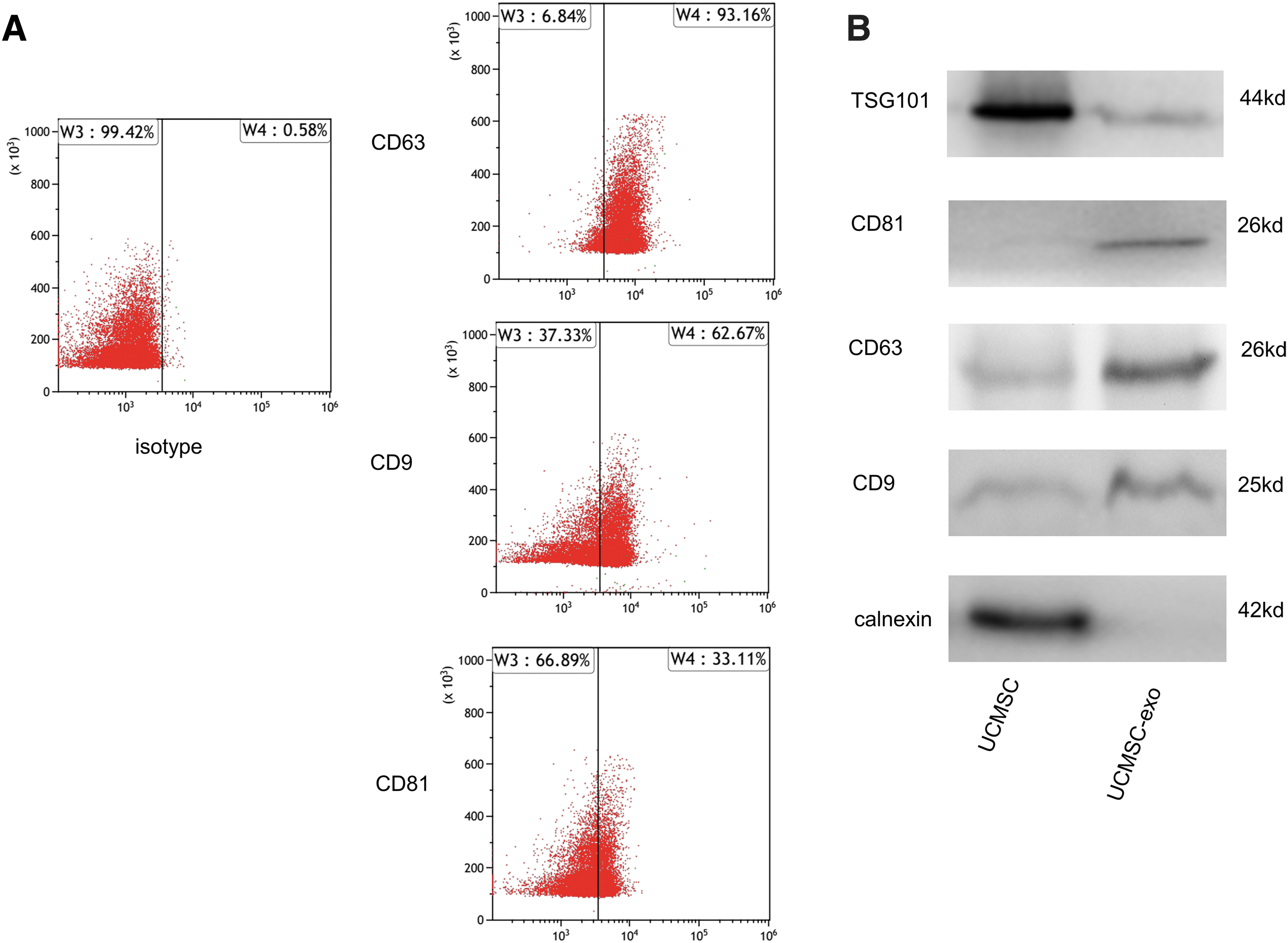
We began by growing UCMSCs with a 3D fibercell system, with SEM used to assess the morphology of 3D fiber material (Fig. 1A), revealing that they provided multidimensional space for cell growth. Exosomes derived from these cells were then collected in serum-free media, and were subjected to an NTA of particle Brownian motion to assess their size and numbers (Fig. 1B). This analysis revealed these particles to have a mean 96 nm diameter with a large peak at ∼120 nm consistent with these particles being exosomes (Fig. 1C). TEM analysis confirmed that these particles were from 55.81 to 136.74 nm in size (Fig. 1D). Dynamic light scattering revealed these UCMSC-derived exosomes to have an average zeta potential of −15.2 mV (Fig. 1E). Analysis of exosomal surface marker expression using flow cytometry revealed these particles to be 93.16%, 62.67%, and 33.11% positive for CD63, CD9, and CD81, respectively (Fig. 2A). Western blotting was consistent with the presence of these surface markers in isolated exosomes and UCMSC-Exo were negative for calnexin (Fig. 2B). Together these findings confirmed that the exosomes derived from UCMSCs in this study were similar to exosomes from other cell sources with respect to their morphology and properties.
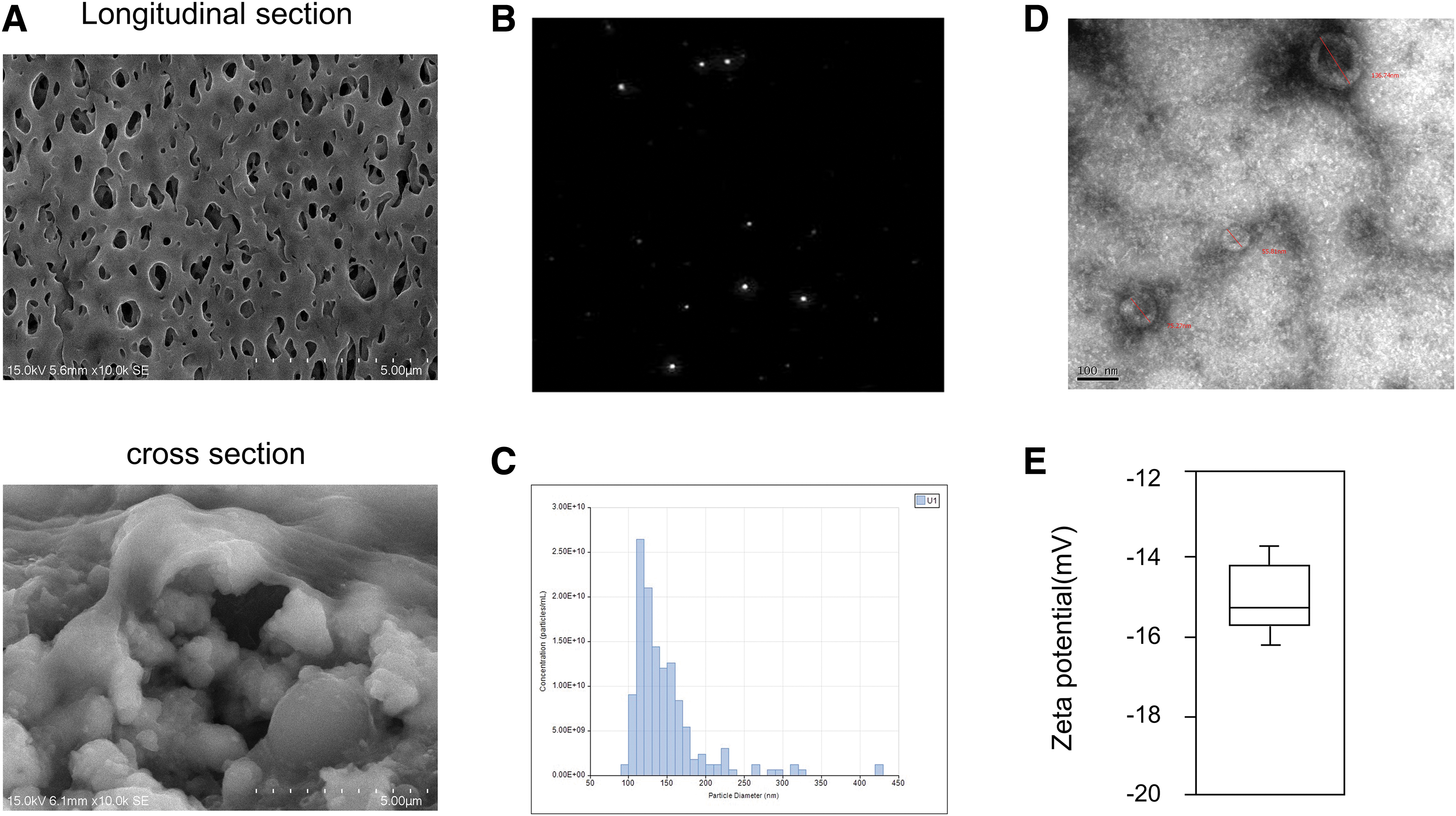
Characterization of UCMSC-Exo.
UCMSC-derived exosomes promoted HaCaT cell proliferation and migration
As UCMSC conditions media was previously shown to drive HaCaT cell proliferation, we next assessed whether the exosomes present in these conditioned media samples were sufficient to drive this proliferation. We found that treating HaCaT cells with 40 × 108 particles/mL of UCMSC-derived exosomes significantly enhanced proliferation relative to control cells in a CCK8 assay. We selected an exosome dose of 80 × 108 particles/mL for further experiments, as values above this dose did not further enhance cell proliferation (Fig. 3A). We next conducted a wound healing assay to explore the ability of these exosomes to modulate keratinocyte migration, revealing that wound closure was enhanced for exosome-treated cells relative to controls (Fig. 3B). This was consistent with UCMSC-derived exosomes directly enhancing HaCaT cell migratory activity. We further confirmed this effect in a transwell migration assay wherein exosome treatment markedly enhanced the number of migratory HaCaT cells in this assay (Fig. 3C). After statistical analysis of the percentage of healing area by unpaired t-test, it was found that the healing area was statistically significant at 3 (p < 0.05), 6 (p < 0.05), and 9 hours (p < 0.05) compared with the control group (Fig. 3D). It can be seen that the penetration of HaCaT cells through the compartment membrane was increased from 12 to 36 hours compared with the control group, which is statistically significant compared with the control group, as given in Figure 3E.
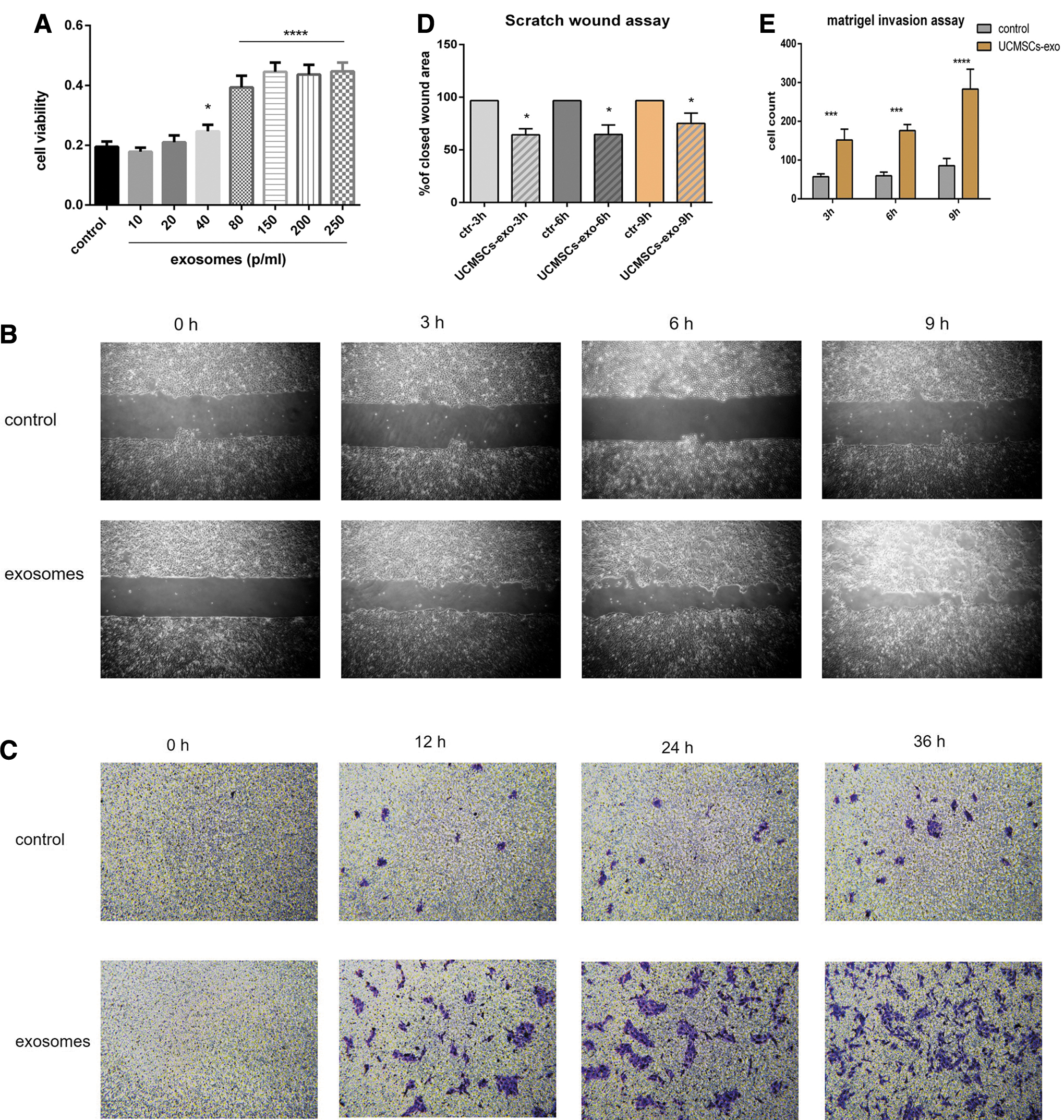
Effect of UCMSC-Exo on the keratinocytes in the proliferation and migration ability.
UCMSC-derived exosomes alter cell viability to HaCaT cells by UVB
We next assessed the ability of USCMC-derived exosomes to protect HaCaT cells from UVB irradiation by treating these cells with exosomes at 80 × 108 particles/mL dose. Although it took time for the UCMSCs-Exo to contact cells through endocytosis and played a role in mediating the cellular signal ways induced by UVB, it was considered that the treatment of UCMSC-Exo after UVB irradiation did not show significant differences in recuperative capacity. Cells irradiated with 30 or 60 mJ/cm2 UVB exhibited significant reductions in viability after 24 and 48 hours, whereas UCMSC-derived exosomes treatment was associated with a significant increase in cell viability relative to untreated controls at both time points (Fig. 4A, B).
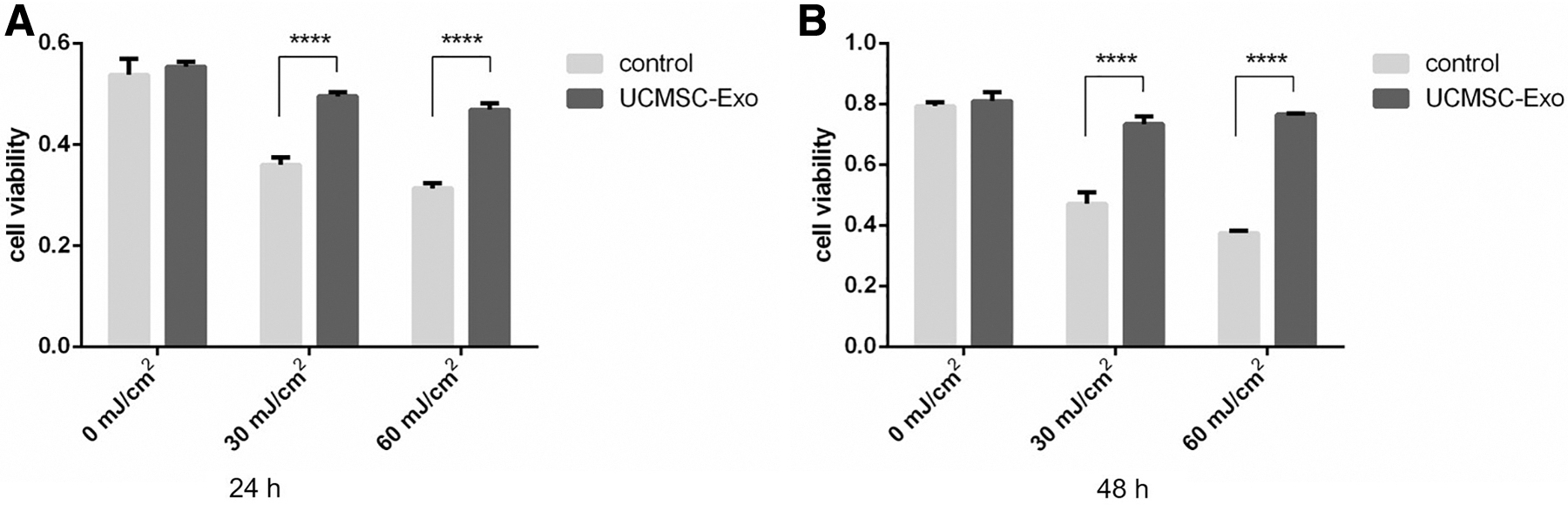
UCMSC-Exo protects the UVB-induced HaCaT cells from photo-aging.
UCMSC-derived exosomes reduced the UVB-induced apoptosis of HaCaT cells
We next directly examined the ability of UCMSC-derived exosomes to inhibit the senescence and apoptotic death of HaCaT cells after 60 mJ/cm2 UVB irradiation. By flow cytometry, we observed a significant increase in the number of apoptotic cells after irradiation, with exosome treatment significantly reducing the frequency of apoptotic cells (Fig. 5A). In a TUNEL staining assay, we further confirmed that UVB irradiation enhanced HaCaT cell death and UCMSC-derived exosomes were able to reduce this apoptotic induction (Fig. 5B). The t-test showed that the number of late apoptotic cells was statistically different after being treated with UCMSC-Exo (p < 0.05) compared with the UVB-treated group, as given in Figure 5C. Meanwhile, the statistics of Tunnel staining also indicated that the number of apoptotic cells was significantly reduced after UCMSC-Exo treatment (p < 0.05), as given in Figure 6G.
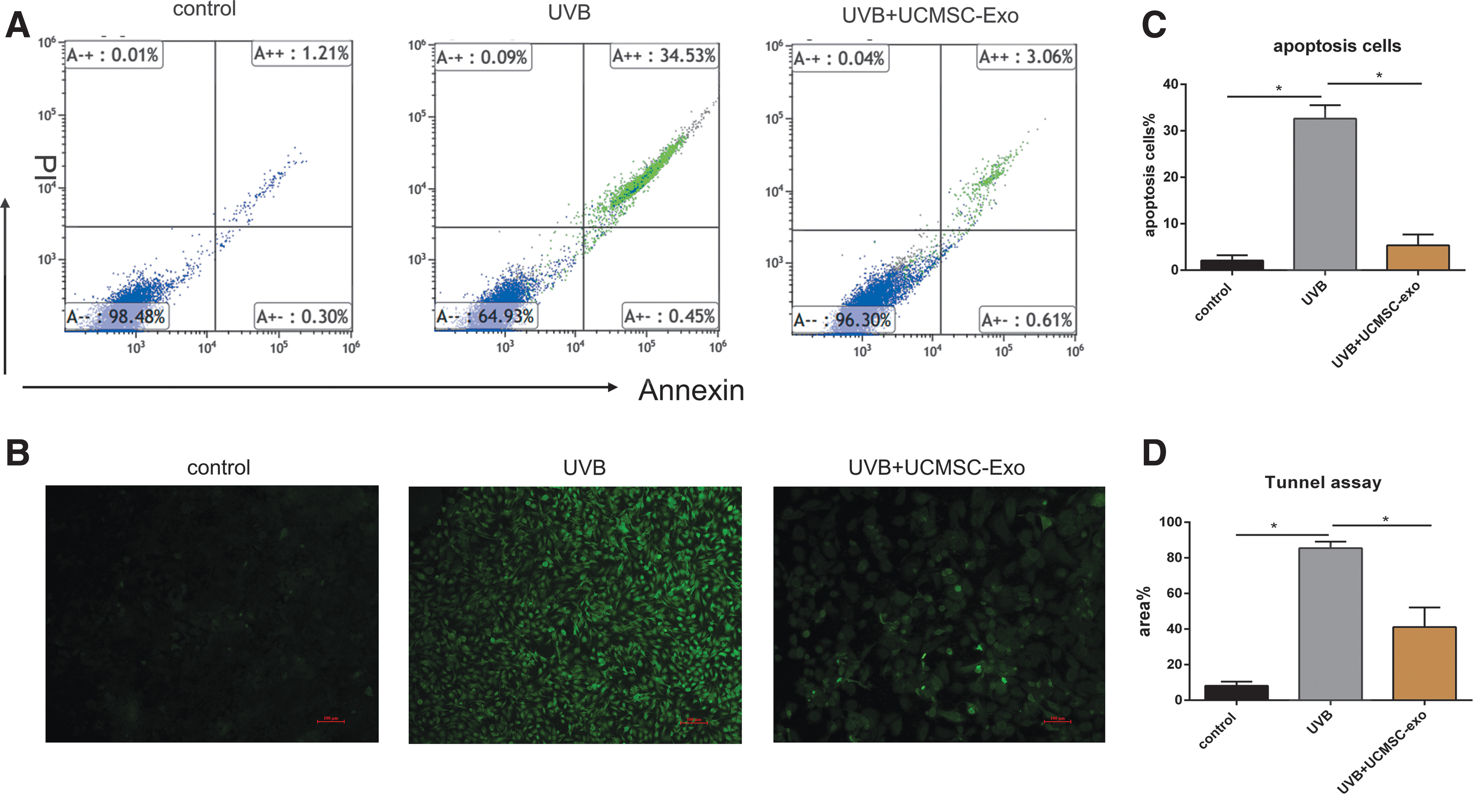
UCMSC-Exo inhibited the production of apoptotic cells induced by UVB irradiation.
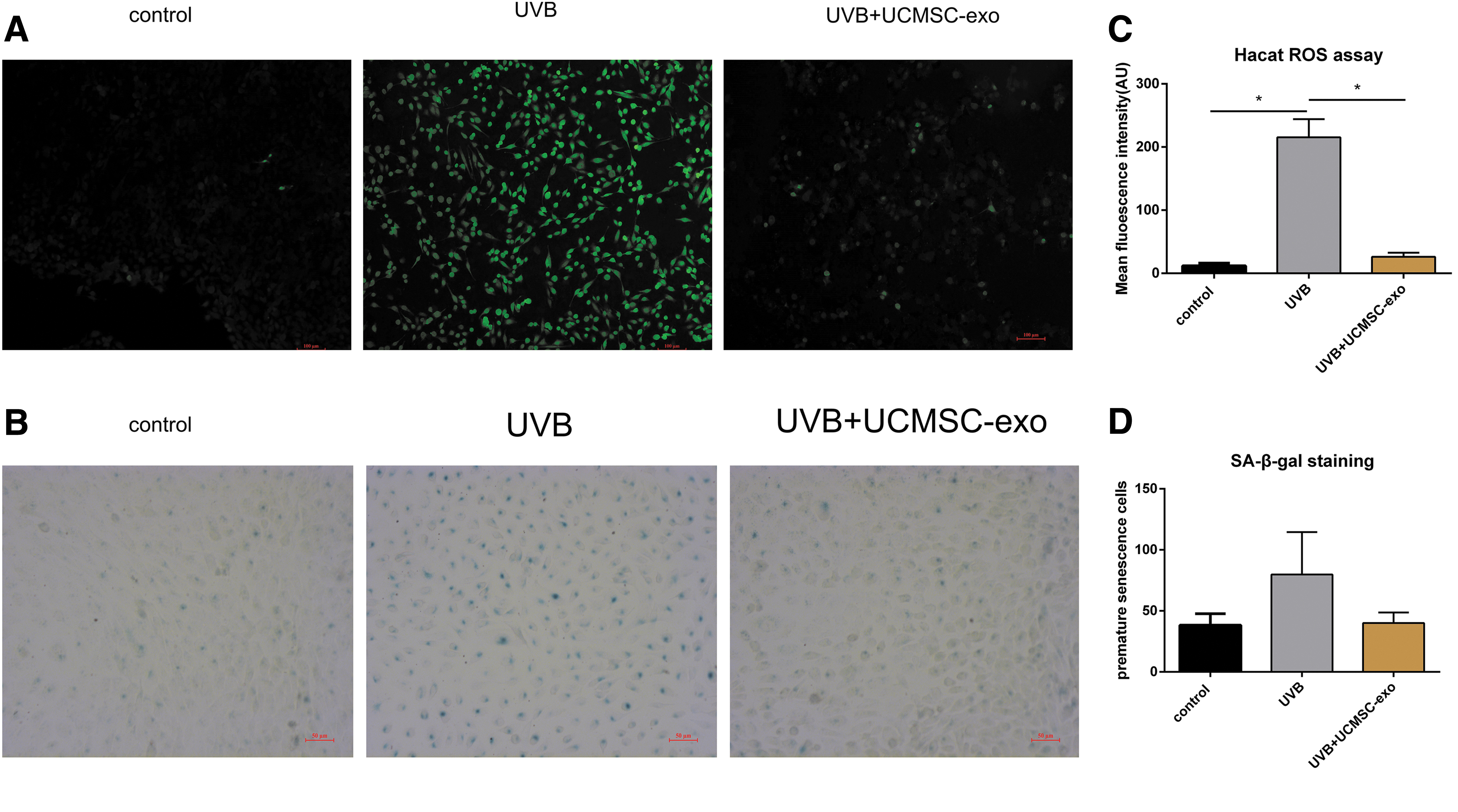
UCMSC-Exo attenuated the ROS generation and play a role in the anti-premature senescence process.
UCMSC-derived exosomes inhibit cellular senescence and ROS production
We next assessed how UCMSC-derived exosomes influenced ROS production in cells, revealing that UVB irradiation was associated with an increase in intracellular ROS activity that was reduced after treatment with these UCMSC-derived exosomes (Fig. 6A). We also assessed how these exosomes influences HaCaT cell senescence by Sa-β-Gal staining of UVB-irradiated cells, as this is a biomarker frequently used for senescent cell detection, with epidermal keratinocytes expressing higher SA-β-Gal levels with age in human skin biopsies. We found UVB irradiation to significantly increase SA-β-Gal expression in HaCaT cells, whereas treatment with UCMSC-derived exosomes decreased the frequency of SA-β-Gal-positive cells. Through the statistical analysis of the fluorescence intensity of ROS, it is found that UCMSC-Exo significantly reduced ROS production after UVB treatment. However, in the statistical analysis of senescent cells labeled by SA-β-Gal, UCMSC-Exo treatment was not shown to be statistically different compared with the UVB-treated group.
UCMSC-derived exosomes affect HaCaT cell mRNA expression
We found that HaCaT cells were able to endocytose UCMSC-derived exosomes (Fig. 7A). UVB irradiation is known to alter gene expression, and as such we next explored the ability of UCMSC-derived exosomes to modulate these changes in gene expression in the context of photo-aging. We observed significant decreases in the mRNA levels for genes encoding collagen type I, procollagen type I, and TIMP1 after UVB irradiation, whereas MMP1 expression was markedly increased relative to control cells (Fig. 7B, E–F). The similar tendency shows after UCMSC-Exo treatment in collagen type I, procollagen type I, and TIMP1. In the comparison of UCMSC-Exo therapy and untreated group, the expression of collagen type I were significantly increased after UCMSC-Exo treatment. However, the expression level of MMP1 and procollagen type I was reduced significantly after UVB irradiation compared with the untreated group. No significant changes in laminin or MMP2 expression were observed in any context. Together these findings suggested that UCMSC-derived exosomes can enhance the activity of matrix-degrading enzymes like MMP1 so as to mediate recovery from photo-aging.
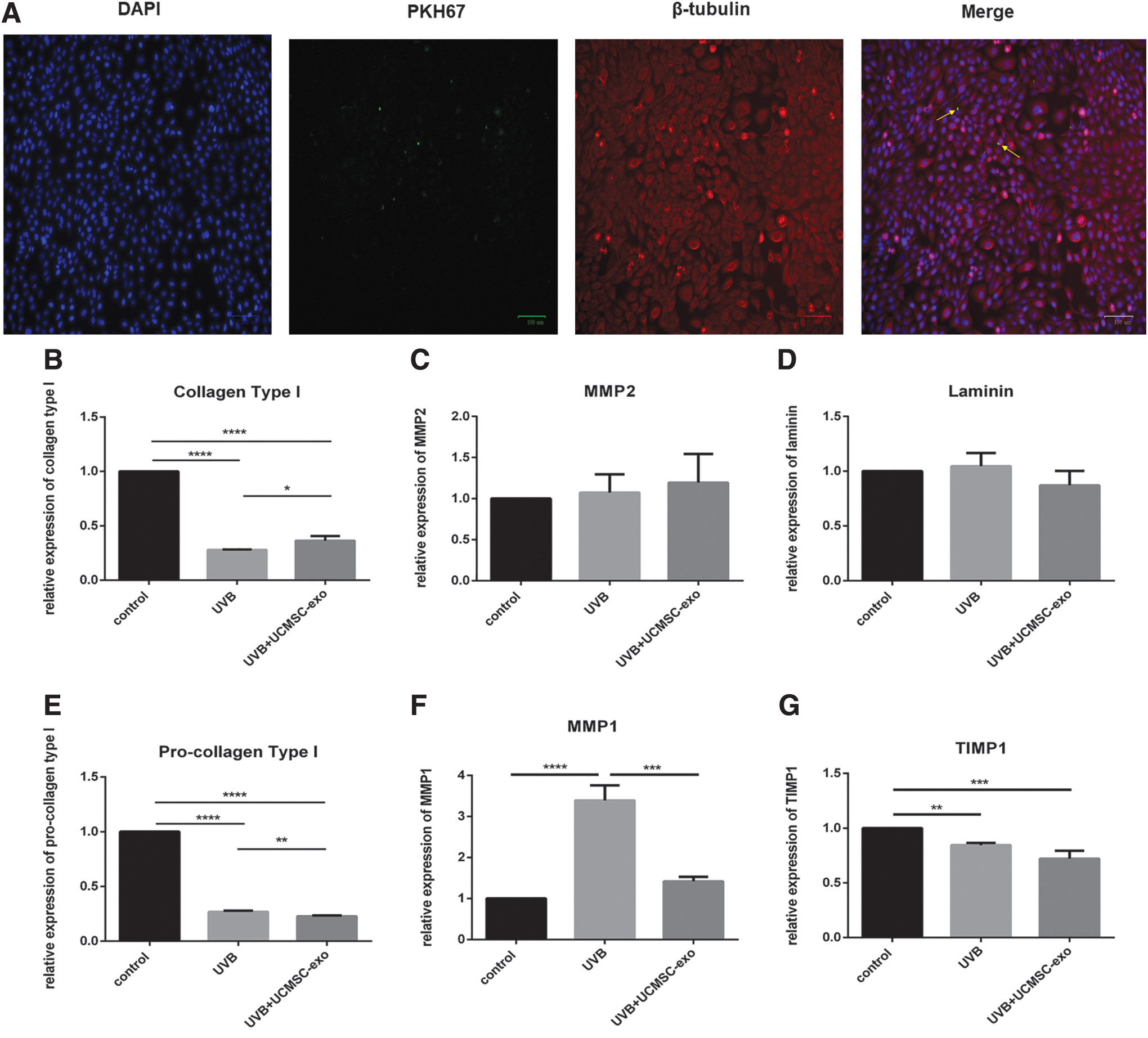
UCMSC-Exo affected the expression of senescence-associated markers after UVB irradiation through endocytosis.
Discussion
Photo-aging is a potentially destructive process that can drive the development of wrinkles, hyperpigmentation, epidermal hyperplasia, and skin cancer. 16 Both UVA and UVB irradiation are major environmental drivers of photo-aging. MSCs have previously been shown to be ideal for treating a range of conditions including skin wounds, ischemic injuries, and photo-aging owing to the paracrine secretory abilities 3,17 and their ability to promote angiogenesis and collagen synthesis so as to drive wound healing. 18 In the previous studies, quantities of studies suggesting that the conditioned medium derived from stem cells had the potential regenerative and reparative effects on prematurely aged skin in vitro and in vivo, which MSCs derived from umbilical cord, amniotic membrane, and adipose tissue were the most common resources.
In addition, exosomes enriched in the conditioned medium have enabled development of cell-free strategies for anti-aging researches. MSC-derived exosomes have increasingly been explored as a valuable tool for mediating the healing of skin wounds, 18,19 and these exosomes have also been assessed in the context of tissue reconstruction. 20,21 Exosomes are potent mediators of intercellular communication, potentially transmitting cytokines, mRNAs, lncRNAs, or miRNAs between cells to mediate tissue regeneration. 22,23 The collection of homogenous and stable preparations of cell-derived exosomes, however, remains challenging, with work suggesting that growing cells in a 3D culture system can enhance their ability to mediate tissue repair. 24,25 Herein we effectively used a fiber-cell culture system to collect large quantities of UCMSC-derived exosomes in serum-free media. When the cells are cultured in our FiberCell Systems, the exosomes in the circulating medium will not enter the cell growth space, and the system is also easy to adapt to the serum-free environment. Furthermore we can obtain high purity and high concentration of exosomes as long as the cells grow well. Compared with 2D culture, the single harvest of medium-sized culture cylinder is equivalent to 200–250 T175 flasks. This is an efficient way to obtain exosomes and we further confirmed that these exosomes were physically and morphologically similar to other cell-derived exosomes.
Epidermal cells are directly impacted by UVB irradiation, and their repair is essential to overcome the damage induced by photo-aging, with compounds such as sunscreen, small peptides, and dietary compounds often being used in an effort to prevent or repair this damage. 26,27 Stem cells are also often used to mediate skin repair, with the injection of stem cells or stem cell-conditioned media being a promising therapeutic strategy. The use of UCMSC-derived exosomes may be associated with a significant reduction in the risk of immunological rejection. USCMSC-derived exosomes treatment has the potential to treat or delay skin damage by inhibiting the apoptotic death of epidermal cells. 28 Apoptosis is a key regulator of photo-aging recovery, 29 and we found that UCMSC-derived exosomes were able to reduce the frequency of apoptotic HaCaT cells after UVB irradiation. UVB exposure is also associated with significant oxidative stress in cells, reducing their integrity and the elasticity of tissues. 30 Stem cell-conditioned media has been shown to inhibit such oxidative stress. 31 UCMSC-derived exosomes were found to suppress ROS generation in response to UVB irradiation in our study; ROS can also be associated with cellular senescence. We found that UCMSC-derived exosomes treatment was associated with a significant reduction in numbers of SA-β-Gal-positive cells after UVB treatment consistent with the ability of these exosomes to suppress keratinocyte apoptosis and senescence. These results are consistent with the anti-aging researches about adipose tissue and amniotic-derived exosomes.
UVB irradiation and associated ROS generation are associated with changes in the expression of many genes, including MMPs and collagens. 32 UV irradiation has been shown to enhance MMP expression and suppress collagen type I synthesis, thereby accelerating skin aging. 33,34 MMPs are ECM-degrading enzymes that can breakdown collagen and elastic fibers in the process of skin aging, with MMP1 and MMP2 upregulation in the context of UV irradiation being particularly important for this process. 35 –37 We found that UCMSC-derived exosomes induced collagen type I expression and reduced the expression of MMP1, suggesting that these exosomes may contain proteins capable of promoting the repair of UVB-damaged cells. This result is consistent with the study of extracellular vesicles in anti-aging and the majority of these researches used human dermal fibroblasts as cellular senescence model. 38,39
Together our findings suggest that UCMSC-derived exosomes may contain essential antiapoptotic factors that allow them to regulate the survival of HaCaT cells. These exosomes are also able to promote repair activities in UVB-irradiated keratinocytes. Despite this being an earlier study that initially applicated UCMSC-derived exosomes in HaCaT cells of photo-aging, the exact molecular mechanisms whereby these exosomes help to reverse photo-aging remain to be identified. In addition, further work is needed to explore the field of exosome biology and confirm these effective components of UCMSC-derived exosomes, which will ultimately contribute to clinical application of exosomes.
Footnotes
Authors' Contributions
We declare that each of the authors made substantial contributions to the article. Experiments were planned and designed by L.Y. Laboratory work and data acquisition were performed by S.L., M.M., S.H., H.G., Y.Z., Y.Y., and Z.L. Data analysis were carried out by L.Y., K.Z., R.H., W.H., R.W., L.Y., W.W., L.L., X.W., Z.H., and L.L. S.L., M.M., and S.H. wrote the article. All authors revised the article. All authors approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from National Natural Science Foundation of China (No.81360245, 81660477, and 81460436) and grants from the Science and Technology Department of Yunnan Province (2017HC027, 2017FE468, 2017IB020, and 2018IA045). This study is also funded by CAMS Innovation Fund for Medical Sciences (CIFMS, 2016-I2M-2-001) and Kunming Science and Technology Innovation and Service Capacity Improvement Program Key Project (2016-2-R-07674).