
Abstract
The heterodimeric integrins are important receptors for the attachment of cells to their extracellular matrix. Here, we studied the attachment of human mesenchymal stem cells (MSCs) to type I collagen (col-1), which is part of the extracellular matrix in bone, skin, and connective tissues. Furthermore, we examined how TGF-β influences the integrin expression and attachment of MSC. Using flow cytometry, immunoblot, and RT-PCR, we report that MSC express several integrin subunits, including the α2β1 integrin (VLA-2, CD49b/CD29). TGF-β increases the expression of integrin subunits α2, α6, and β1 in MSC, thereby enhancing the attachment of MSC to col-1. The TGF-β-mediated up-regulation of the expression of the integrin subunits α2 and α6 is mainly mediated in MSC by Smad2.
Introduction
M
Because of their differentiation capacity, MSC are a highly promising tool in therapeutic approaches [14,15]. Once placed in the defective tissue in combination with an adequate carrier material, they are able to contribute to tissue regeneration. Important aspects in MSC research include finding optimal carrier materials for the tissue engineering application being developed and studying the effects of soluble factors that are deposited in the corresponding tissue during regenerative processes. Carrier materials should integrate easily into the target tissue, supporting the integration of MSC and providing factors necessary for their regenerative action.
In many tissues of the human body, collagens are prominent components of the ECM, so they would seem to be suitable candidates for tissue engineering carrier materials. Long-term bone marrow cultures and gene expression profiling revealed that bone marrow MSC themselves produce collagens as well as laminins 16 –19]. Furthermore, bone marrow contains cytokines and growth factors that are either soluble or bound to the ECM components [20]. Outside the bone marrow, MSC are situated near the basement membranes of blood vessels, which mainly consist of type IV collagen (col-4), laminin-411 (LM-411), and laminin-511 (LM-511) [21]. MSC express surface proteins for ECM attachment [22 –24] and growth factor receptors [25,26]. TGF-β is one of the factors to which the MSC respond [17,27].
In this study, we analyzed the effect of TGF-β on the attachment of MSC to type I collagen (col-1). Type I collagen (col-1) is the most abundant protein in the ECM. In bone 90% of the ECM protein mass is col-1, and it is found in other connective tissues as well. Receptors for the attachment to col-1 are integrins α1β1, α2β1, α3β1, α10β1, and α11β1 [28]. We also investigated the interaction of MSC with LM-111 and compared it to the attachment to col-1. LM-111 is the first laminin expressed during embryonic development [29]. In the adult, it is part of the basal membranes in epithelia. LM-111 could serve as a carrier material for bone tissue engineering or coating for prostheses, as it supports osteogenic differentiation of MSC [30,31]. Integrins binding to LM-111 are the α1β1, α2β1, α6β1, α7β1, and α9β1 receptors [29].
TGF-β is a growth factor involved in wound healing, in matrix turnover during fibrotic processes [32], as well as in osteogenesis [33], and it induces chondrogenic differentiation in MSC [1]. TGF-β regulates the attachment and integrin expression pattern in various cell types [34 –37], including synovial fibroblasts (SF) [38]. Here, we analyzed the attachment of MSC to col-1 and LM-111, and we examined the influence of TGF-β on the integrin expression pattern and the attachment of MSC.
Materials and Methods
Isolation and characterization of MSC
MSC were isolated by Ficoll-Paque™ (Amersham Biosciences, Freiburg, Germany) gradient centrifugation from femoral bone marrow aspirates of patients undergoing endoprosthetic surgery. This study was approved by the local ethics committee. MSC were grown in Mesenchymal Stem Cell Growth Medium (Lonza, Basel, Switzerland).
Characterization of MSC was performed by flow cytometry and differentiation assays [1,39]. Flow cytometry analysis of MSC included staining with antibodies against the cell surface inclusion markers CD73 (BD Pharmingen, Heidelberg, Germany), CD90 (R&D Systems, Wiesbaden, Germany), CD105 (Serotec, Düsseldorf, Germany), and exclusion markers CD34 (Biolegend, Echingen, Germany), CD14, and CD45 (R&D Systems). Differentiation of MSC into adipogenic, osteogenic, and chondrogenic lineages was induced by specific differentiation media.
Adipogenic differentiation medium contained 0.1 mM indomethacin (Calbiochem, Darmstadt, Germany), 0.01 mg/mL insulin, 0.5 mM 3-isobutylmethylxanthine, and 1 μM dexamethasone (all from Sigma-Aldrich, Munich, Germany). Differentiation was detected by Oil Red staining. For this purpose, a 0.5% Oil Red stock solution in isoporopanol was diluted to 3:2 with water. After filtration, the cells were incubated in Oil Red staining solution for 30 min. Then cells were washed 3 times with PBS and recorded by microscopy (Leica DM-IRBE). Expression of LPL and PPARγ mRNA was measured by quantitative RT-PCR (qRT-PCR) using specific primers (Table 1).
Osteogenic differentiation medium consisted of DMEM supplemented with 0.1 μM dexamethasone, 10 mM β-glycerophosphate (Merck, Darmstadt, Germany), and 50 μM
Chondrogenic differentiation was initialized in pellet cultures using 10 ng/mL TGF-β3 (R&D Systems). After 4 weeks, proteoglycans in the pellets were stained with Alcian blue using a 1% Alcian blue solution in 3% acetic acid, and types I and II collagen were detected by immunohisto-chemical staining (monoclonal antibodies for types I and II from MP Biomedicals, Illkirch, France) and documented by microphotography [40].
Cell attachment assays
Attachment assays [41] were performed to investigate the binding of MSC to col-1 and LM-111 and to display the effect of TGF-β on the adhesion of MSC to these proteins. In brief, 0.01 fmol to 10 fmol of col-1 or LM-111 (both from BD Falcon Heidelberg, Germany) were dissolved in 2 μL PBS or 0.2 N acetic acid, respectively, spotted onto tissue culture dishes and air-dried, followed by a blocking step using 10 mg/mL BSA/PBS. Spots of 100 fmol and 1 pmol poly-
To detect TGF-mediated effects, the MSC were preincubated with 10 ng/mL TGF-β1 (R&D Systems) for 24 h prior to the attachment assay or left untreated as controls. Then, cells were harvested by mild detachment, resuspended in serum-free medium with or without integrin-blocking antibodies (listed in Table 2)for 30 min, and seeded onto the protein spots in the cell culture dishes. After 15 min of incubation on the spots, the floating cells were removed and the adherent cells were recorded by microphotography.
Analysis of integrin expression by flow cytometry
The cells were either stimulated with 10 ng/mL TGF-β1 for 24 h or left untreated as controls. After harvesting and washing in PFEA buffer (PBS with 2 mM EDTA, 2 % FCS, and 0.01% sodium azide), primary antibodies specific for integrin β1 (clone 4B4; Beckmann Coulter, Krefeld, Germany), α1 (PE-labeled, clone SR84; BD Pharmingen), α2 (PE-labeled, clone 12F1; BD Pharmingen), α3 (clone P1B5; Chemicon, Schwalbach, Germany), α6 (PE-labeled, clone 450–30A; Abcam, Cambridge, MA), α9 (clone Y9A2; Abcam), and α11 (Abnova, Taipei, Taiwan) were incubated for 20 min at 4°C. In the case of the unlabeled primary antibodies, the incubation with the PE-labeled secondary antibody (Jackson Immunoresearch, Newmarket, UK) was carried out for 20 min at 4°C. Cells were analyzed in a flow cytometer (BD LSR II) using the FACS® DIVA software.
Quantitative RT-PCR analysis
For analysis of transcripts, RNA was isolated from MSC, which were either left untreated or stimulated with 10 ng/mL TGF-β1. To perform reverse transcription, we utilized 1 μg RNA (RT-for-PCR-Kit, Clontech, Saint-Germainen-Laye, France). Equal amounts of the cDNA were analyzed by qRT-PCR. Hot Start Taq polymerase and SybrGreen from the Light Cycler® FastStart DNA Master kit (Roche, Penzberg, Germany) were employed for amplification and detection of the PCR products. Primers for the differentiation markers were designed according to published sequences [43] and are listed in Table 1. Integrin PCR primers are listed in Table 3. Primers for the differentiation markers are listed in Table 1. Primers for GAPDH and a recombinant GAPDH standard were obtained from Search LC (Heidelberg, Germany). The amplification was performed in the Light Cycler 2.0 (Roche), and relative quantification of the amplificates was carried out using the Fit Points method [44].
Western blot analysis
To investigate the expression of proteins, western blot analyses were performed as described [38]. MSC were lysed in RIPA buffer (c-c-pro, Oberdorla, Germany). The protein concentration in lysates was measured using the RCDC Kit (BioRad, Munich, Germany), and 100 μg of protein per lane were applied to the SDS-PAGE. After blotting, the nitrocellulose membrane was blocked with 5% milk/PBS/0.1% Tween-20 (blocking buffer). For detection of the integrin α7 subunit, the blots were incubated overnight with antiintegrin α7 antibody (1:1,000; Abcam) in blocking buffer. Secondary HRP-labeled goat anti-mouse antibody (Jackson Immunoresearch) was used in the ratio of 1:1,000 in blocking buffer. Binding of antibodies was visualized by chemiluminescence (ECL; GE Healthcare, Freiburg, FRG) and recorded by a luminescence-sensitive CCD camera system (Diana, Raytest Inc., Straubenhardt, FRG).
TGF-β signaling pathway
For investigation of the TGF-β-dependent signaling and gene regulation, qRT-PCR was employed. The cells were preincubated for 30 min with inhibitors of the TGF-β signaling pathway. The following inhibitors were used at the given concentrations: 100 nM of TGF-β RI kinase inhibitor, 20 μM of the MEK inhibitor PD98059, and 20 μM of the p38 inhibitor SB203580 (all from Calbiochem). After preincubation with the inhibitors, cells were exposed to 10 ng/mL of TGF-β1. Mock-treated MSC or MSC activated with 10 ng/mL TGF-β1 without inhibitors served as controls. After 24 h of incubation, RNA was isolated and qRT-PCR was performed to quantify integrin mRNA transcripts.
Statistical analyses
Statistical significance was assessed by 2-sided Student's t-tests. Probability values (P) ≤0.05 (*), 0.01 (**), or 0.001 (***) were considered to be statistically significant and marked in the figures accordingly.
Results
Characterization of MSC
To characterize the isolated and expanded MSC, we analyzed their differentiation potential and the expression of typical surface marker proteins [39]. The cells purified from bone marrow aspirates displayed strong staining for CD73, CD90, and CD105 and expressed low levels of CD14, CD34, and no CD45 (Fig. 1). Furthermore, upon appropriate stimulation, MSC differentiated into cells of the adipogenic, chondrogenic, and osteogenic lineages (Fig. 2). Figure 2A displays the spindle-like shape of undifferentiated MSC. Adipogenic differentiation of MSC was confirmed by Oil Red staining of the intracellular lipid vesicles (Fig. 2B) and by the enhanced expression of the adipogenic marker genes PPARγ (10.3 ± 1.7-fold, n = 4, P < 0.01) and LPL (7,425 ± 7,079-fold induction over undifferentiated control cells, n = 4, P > 0.05, Fig. 2C). Chondrogenically differentiated cell pellets contain proteoglycans, which were detected by Alcian blue staining (Fig. 2D). Immunostaining demonstrates that these pellets contain collagen type I (Fig. 2E), but more important for the chondrogenic differentiation, collagen type II (Fig. 2F), corroborating our recent in-depth study [40]. In osteogenically differentiated MSC, mineralization was proven by Alizarin Red (Fig. 2G) and von Kossa staining (Fig. 2H). Enhanced levels of mRNA encoding alkaline phosphatase (29 ± 39.8-fold induction over undifferentiated control cells, n = 4, P > 0.05), osteocalcin (3 ± 2.3-fold, n = 4, P > 0.05), osteopontin (2 ± 0.5-fold, n = 4, P < 0.05), and Runx-2 (6 ± 5.7-fold, n = 4, P > 0.05) were detected in osteogenically differentiated MSC in comparison to undifferentiated MSC (Fig. 2I).
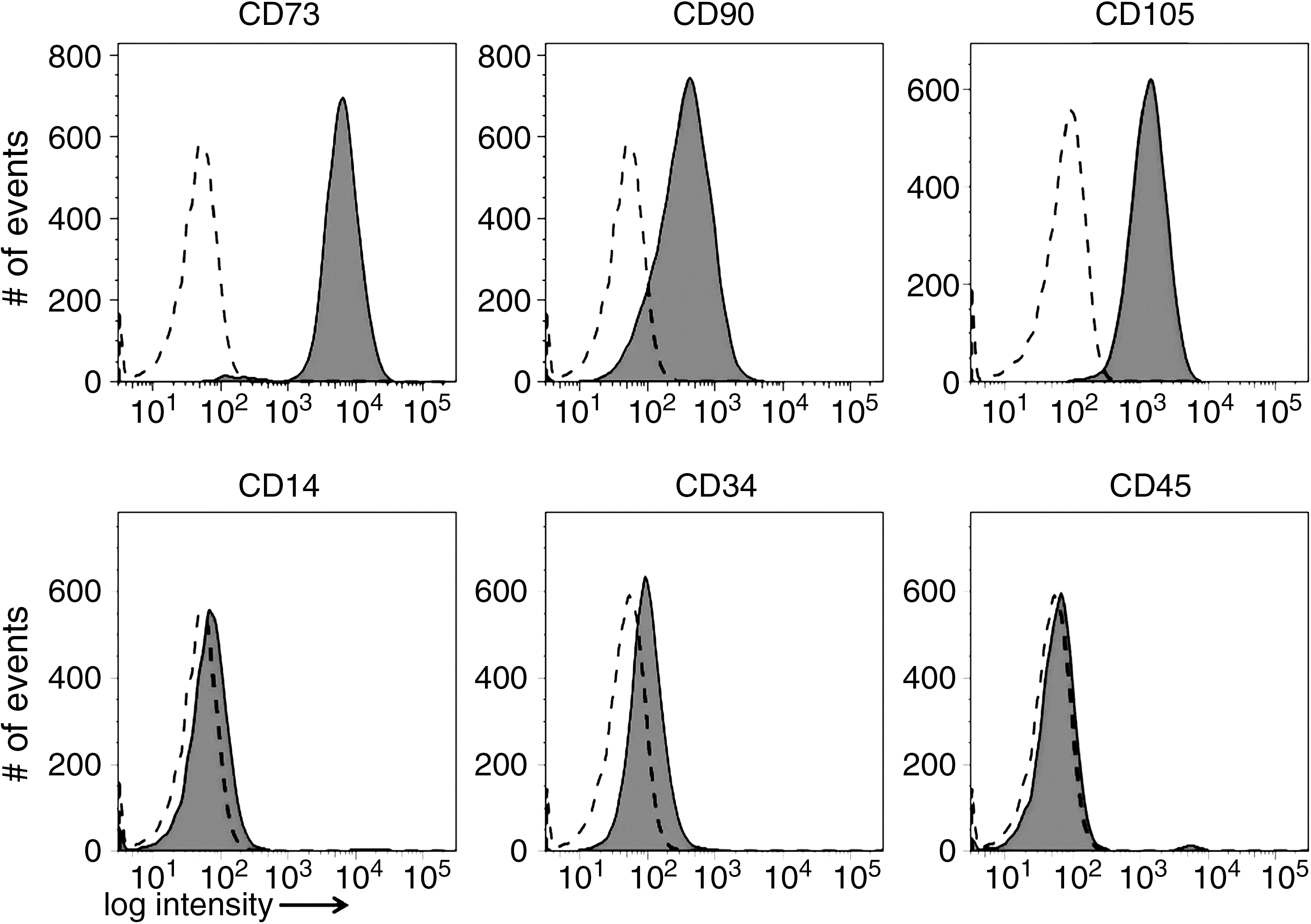
Expression of cell surface antigens on mesenchymal stem cell (MSC). Human bone marrow-derived MSC were investigated for the expression of the MSC marker antigens CD73, CD90, and CD105 by flow cytometry. For MSC exclusion marker antigens are CD14, CD34, and CD45. Flow cytometry analysis revealed that cells isolated from bone marrow expressed the typical surface markers for MSC [39]. The dashed lines denote unstained cells, tinted areas the fluorescent-stained cells. The signal intensities are presented in the histogram mode on the x-axes and the number of cells staining on the y-axes.

Differentiation of mesenchymal stem cell (MSC) into adipogenic, osteogenic, and chondrogenic lineages. To confirm the differentiation capacity of the MSC after expansion in vitro, the cells were split, incubated for 28 days adipogenic, chondrogenic, or osteogenic differentiation media, and analyzed for a characteristic phenotype and gene expression. (
Attachment of MSC to extracellular matrix proteins
The attachment of MSC to immobilized spots of col-1 and LM-111 was investigated. MSC attach well to spots of 10 fmol and 1 fmol, to some extent to 0.1 fmol, and only weakly to 0.01 fmol of col-1. Furthermore, they attach well to 10 fmol or 1 fmol of LM-111, whereas 100 fmol of LM-111 per spot seemed to be above optimal ranges (Fig. 3). Attachment to 100 fmol of PLL served as control (Fig. 3).
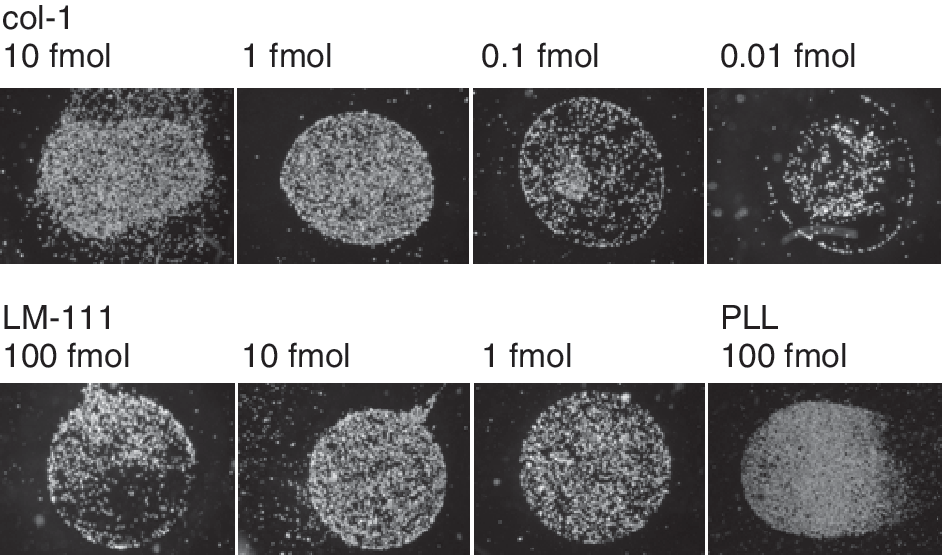
Attachment of mesenchymal stem cell (MSC) to extracellular matrix (ECM) proteins. To investigate the attachment of MSC to type I collagen (col-1) or LM-111, MSC were harvested and incubated in serum-free medium for 15 min on serial dilutions of col-1 (spots of 10 fmol and 0.01 fmol) and LM-111 (spots of 100 fmol and 1 fmol). Floating cells were removed and the MSC attached to the spots were recorded. Attached cells are visible as white dots on the protein spots, while areas coated by BSA only and those free of cells are visible as black zones. The MSC attached well to spots of 10 fmol and 1 fmol of col-1, to some extent to 0.1 fmol, and only weakly to 0.01 fmol of col-1 (top). MSC attached well to 10 fmol or 1 fmol of LM-111, to a lesser extent to 100 fmol of LM-111 (bottom). Attachment of MSC to poly-
The integrin β1 subunit is part of the major extracellular matrix receptors for the binding of cells to collagens and laminins. Using the integrin β1-specific antibody 4B4, we could completely block the attachment of MSC to col-1 and LM-111 (Fig. 4). Furthermore, the attachment of MSC to col-1 could be reduced by the integrin α2-specific antibody P1E6, while the attachment to LM-111 was not affected by this antibody (Fig. 4). Using function-blocking antibodies against integrins α3, α6, and α9 had no effect on the attachment of MSC to col-1. Anti-integrin α9 antibody treatment had a minor effect on binding of MSC to LM-111. Cells on PLL spots served as controls. Incubation of the cells even with the β1-specific integrin-blocking antibody had no effect on the attachment of MSC to PLL (Fig. 4).
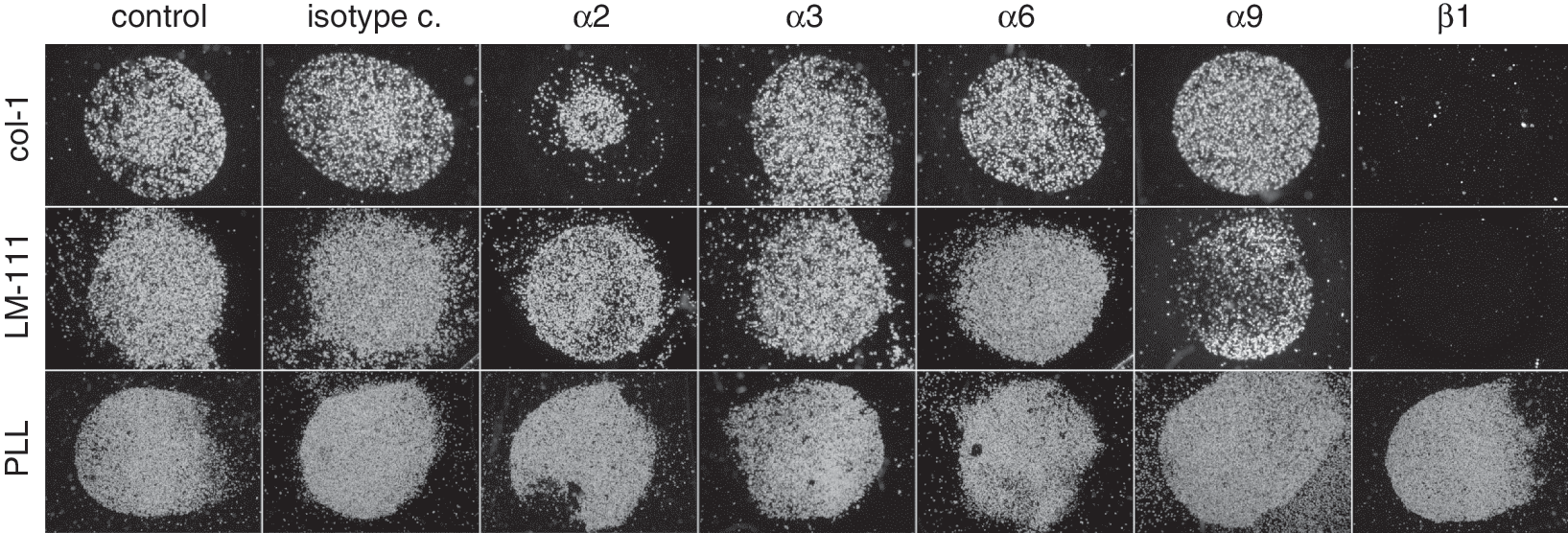
Contribution of individual integrins to the attachment of mesenchymal stem cell (MSC) to type I collagen (col-1) and LM-111. The MSC were harvested and either preincubated with function-blocking antibodies (see Table 2)or left untreated, to investigate the binding on spots of col-1, LM-111, or poly-
TGF-β influences the attachment of MSC to type I collagen
The effect of TGF-β1 on the attachment of MSC to col-1 and LM-111 was analyzed in a concentration-dependent cell attachment assay (Fig. 5). Cells were either left untreated or preincubated with TGF-β1 for 24 h prior to the attachment assay. TGF-β1 enhanced the attachment of MSC to col-1, as TGF-β-activated MSC displayed binding to 1 fmol, 0.1 fmol, and to some extent even to 0.01 fmol, whereas the mock-treated MSC failed to attach to 1 fmol of col-1. Spotting with concentrations as low as 10 fmol of col-1 revealed a difference in cell density of TGF-β1-activated cell attached to col-1 compared to the mock controls (Fig. 5). In contrast, the attachment of MSC to LM-111 was not affected by TGF-β1 (Fig. 5). The naïve MSC attached to spots of 10 fmol and 1 fmol of LM-111 with the same density as the TGF-β1-activated cells did. At 0.1 fmol or 0.01 fmol of LM-111 per spot specific binding of MSC fainted. Only some TGF-β1-activated MSC spread anywhere on the slides, but not specifically on the LM-111-coated spot (Fig. 5).
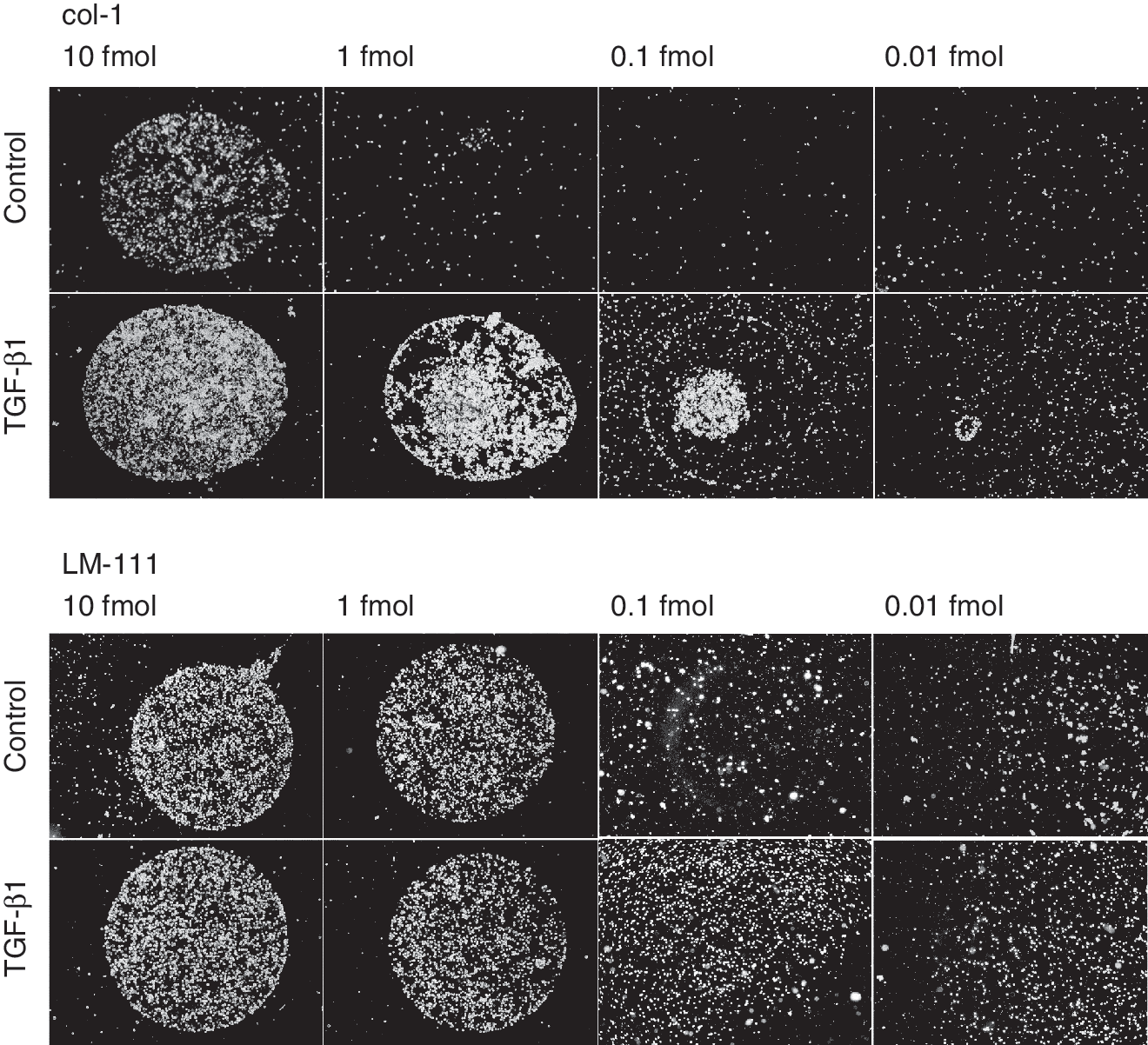
Influence of TGF-β1 on the attachment of mesenchymal stem cell (MSC) to type I collagen (col-1) and laminin-111. The activation of MSC by TGF-β1 (10 ng/mL, 24 h) enhances the attachment of the cells to col-1 (10, 1, 0.1 fmol) in comparison to untreated MSC. An enhanced attachment of TGF-β1-activated MSC to LM-111 (10, 1, 0.1, 0.01 fmol) was not observed.
Integrin expression pattern on MSC and SF
Attachment to col-1 and LM-111 is accomplished by integrin receptors, while adhesion to PLL occurs via electrostatic interactions between the negatively charged lipid membrane and the positively charged PLL. Our results suggested that TGF-β may change the integrin expression pattern in MSC, resulting in a stronger attachment to col-1. To confirm this hypothesis, integrin expression patterns were analyzed in more detail.
Performing RT-PCR analyses, we could detect the designated amplificates for integrin subunits α1, α2, α3, α6, α7, α9, α10, α11, and β1 in MSC (Fig. 6A). Transcripts were then quantified by qRT-PCR (Fig. 6B). MSC show high transcript levels for integrins α1 (5.9 × 10−;3), α3 (2 × 10−;2), α7 (1.9 × 10−;2), α11 (2.2 × 10−;2), and β1 (1.3 × 10−;1) and low expression of α2 (1.9 × 10−;4), α6 (6.8 × 10−;4), α9 (2.8 × 10−;5), and α10 (5.3 × 10−;4) mRNA. The depicted transcript levels are normalized to the expression of the housekeeping gene GAPDH. Flow cytometry analyses revealed high expression of integrin subunits α1, α2, α3, and β1 on MSC, and a mid-level expression of integrin subunits α6, α9, and α11 (Fig. 6C). A strong expression of integrin α7 was detected on MSC by western blotting (Fig. 6D).
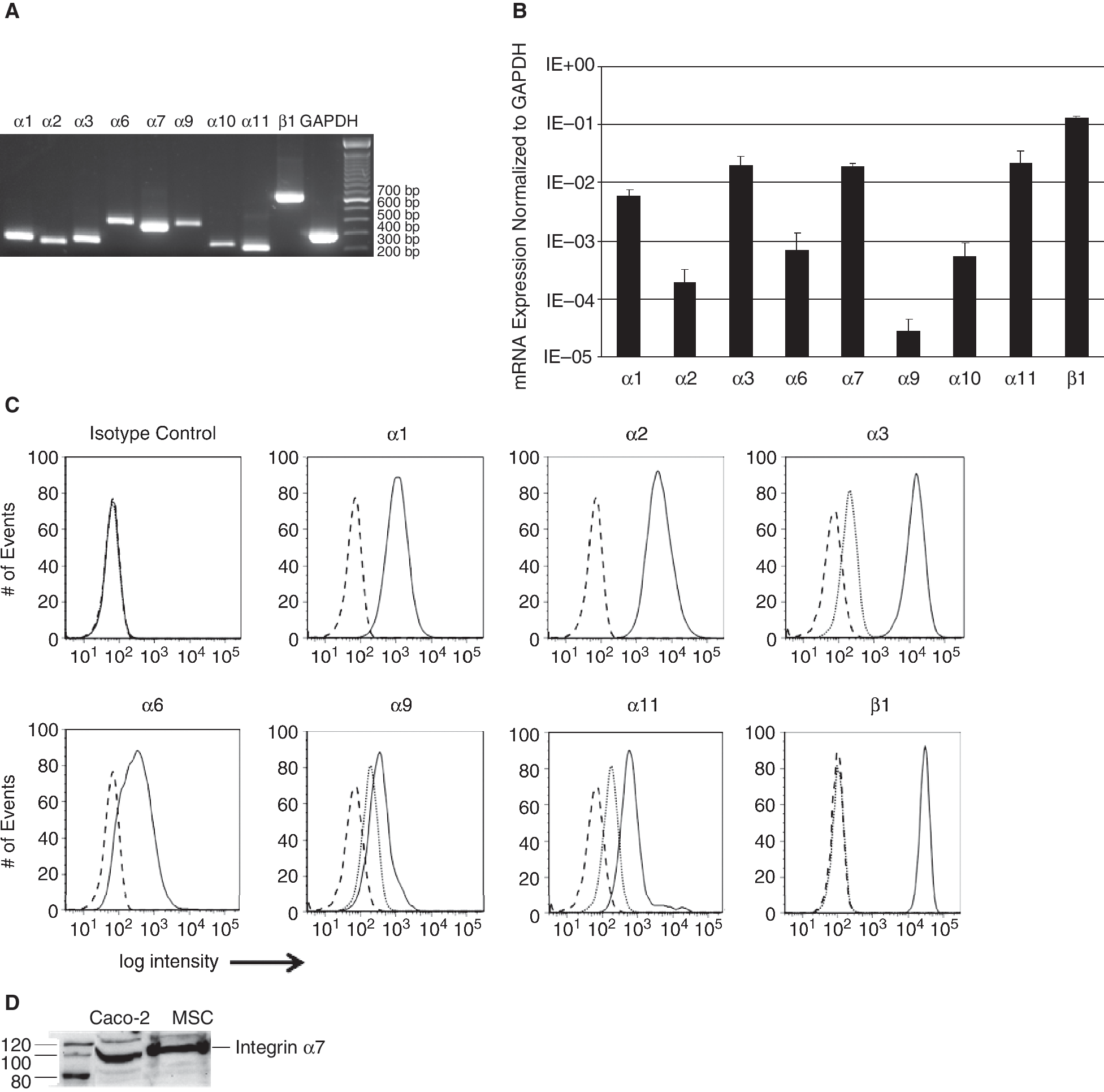
Integrin expression in mesenchymal stem cell (MSC). The expression of type I collagen (col-1)- and LM-111-binding integrins was analyzed by RT-PCR and qRT-PCR, flow cytometry, and immunoblot. (
Influence of TGF-β on the integrin expression of MSC
To determine the effect of TGF-β1 on integrin expression, MSC were stimulated with TGF-β1 for 24 h, and the expression patterns of the integrin subunits were investigated by qRT-PCR and flow cytometry (Fig. 7). In MSC, TGF-β1 enhanced the mRNA encoding integrin subunits α2 (15.8-fold induction over untreated controls, P ≤ 0.05), α6 (4.7-fold induction, P ≤ 0.05), and β1 (1.8-fold induction, P ≤ 0.01) statistically significant (Fig. 7A). The gain of the integrin β1 transcripts was rather low, as this subunit was expressed in naïve MSC at high levels (Fig. 6B and 6C). The transcript levels of the other integrin chains under investigation were not regulated by TGF-β1 (Fig. 7A). The cell surface expression of the α1, α3, and α9 integrin subunits was not regulated by TGF-β1, the signal observed for the α11 integrin was reduced, but the expression of α2, α6, β1 integrin subunits was slightly elevated by TGF-β1 (Fig. 7B). The TGF-β1-mediated gain of integrin α2 signal intensity observed by FACS was low, and the peaks of the histograms were not clearly separated. In contrast, the mean of intensity for integrins α6 and β1 shifted to higher signal intensity, indicating an elevated assemblage of both subunits on the surface of the MSC (Fig. 7B).
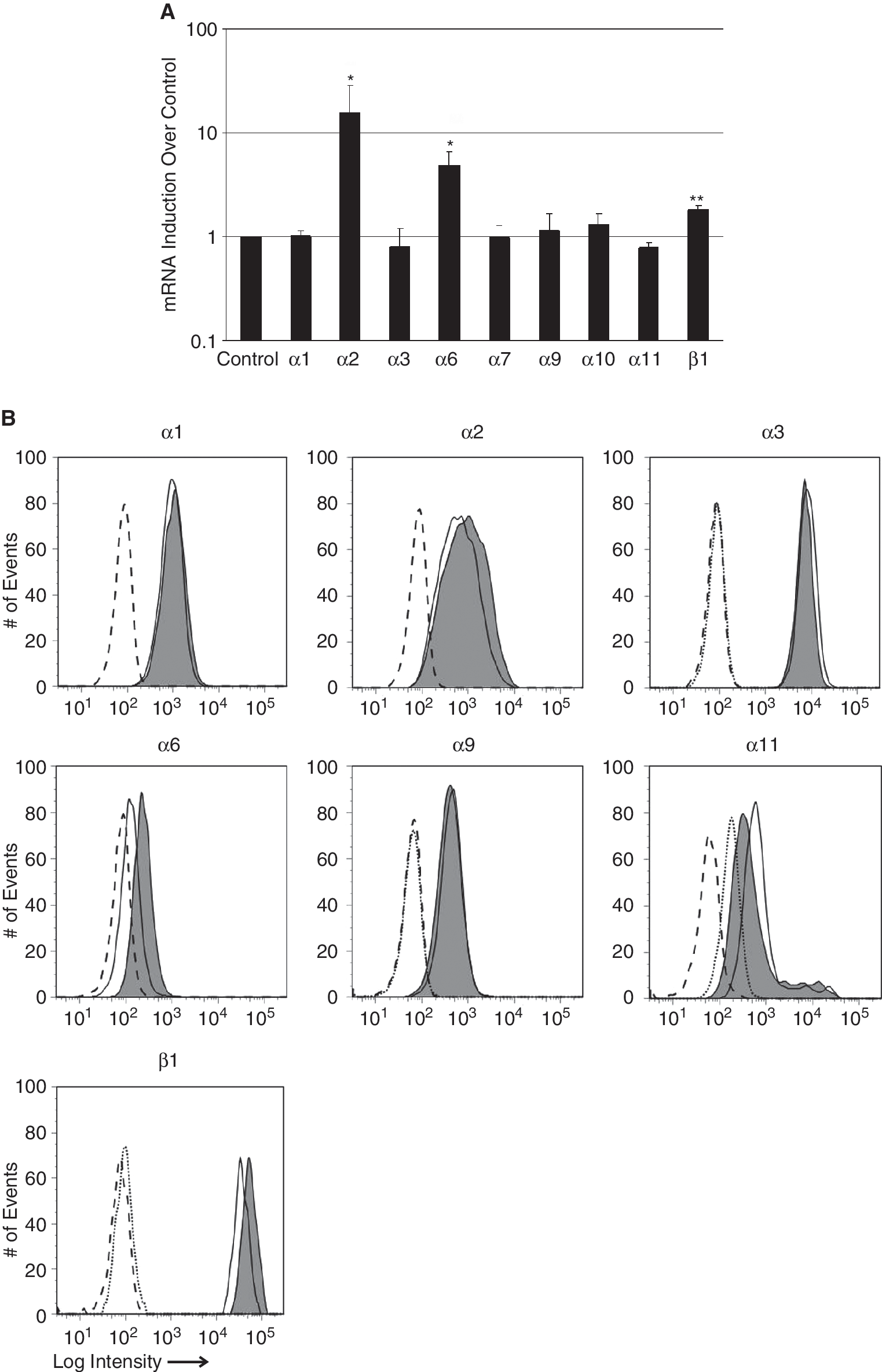
Regulation of integrin expression in mesenchymal stem cell (MSC) by TGF-β1. MSC were activated by TGF-β1 (10 ng/mL, 24 h) or left untreated and regulation of integrin expression was investigated. (
TGF-β-mediated signaling pathway in MSC
MSC were incubated with TGF-β1 and in parallel exposed to inhibitors for different components of the TGF-β-signaling pathway, including SB203580, PD98059, and TGF-β RI kinase inhibitor. Twenty-four hours later, mRNA expression of TGF-β1-regulated integrin subunits α2, α6, and β1 was measured by qRT-PCR.
The expression of the β1 integrin was least modulated by interfering with the TGF-dependent intracellular signaling and p38 MAPK blocker SB203580, ERK blocker PD98059, TGF-β RI kinase inhibitor, which interferes with the SMAD2 signaling, had no significant effects (Fig. 8). The expression of the α2 integrin was activated by TGF-β1 but the inhibition of the p38MAPK or ERK pathways by SB203580 or PD98059 had no effect. But blocking the SMAD2 signaling by addition of the TGF-β RI kinase inhibitor reduced the TGF-β1-induced expression of α2 integrin (Fig. 8). TGF-β1 enhanced the expression of the α6 integrin statistically significant (P < 0.05). Addition of SB203580 to the activated MSC did not reduce the expression of the α6 integrin, but PD98059 reduced its expression to some extent. Inhibition of the SMAD2-signaling pathway by addition of the TGF-β RI kinase inhibitor reduced the TGF-β1-induced expression of α6 integrin statistically significant (P < 0.05, Fig. 8). In MSC, TGF-β triggers the phosphorylation of Smad 2 at its C-terminus (Ser465/467), while the levels of phospho-ERK1/2 and phospho-p38 were not affected by TGF-β1 (data not shown). We conclude the phosphorylation of Smad 2 induces a signaling pathway that results in the enhancement of integrin α2, α6, and β1 expression in MSC.
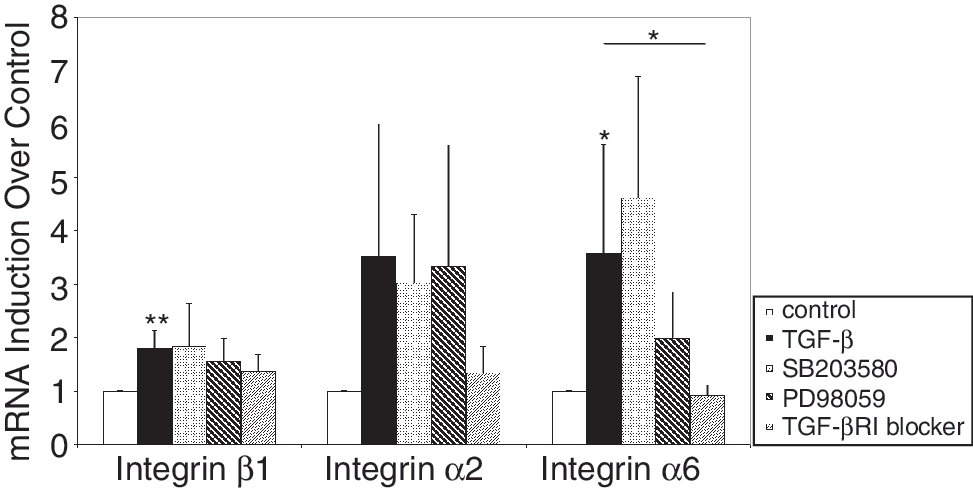
Modulation of the TGF-β1-induced expression of integrins. Mesenchymal stem cell (MSC) were incubated with TGF-β1 in the presence or absence of inhibitors for the TGF-β signaling pathway, and expression of mRNA encoding integrin subunits α2, α6, and β1 was measured by qRTPCR. Untreated cells (white bars) and TGF-β1-activated cells (black bars) served as controls. The expression of the mRNA encoding the β1, α2, and α6 integrin subunits was diminished by the TGF-β RI inhibitor, which precedes the phosphorylation of Smad2. The ERK inhibitor PD98059 reduced the induction of integrin α6 chain, but not of α2 and β1 expression, and SB203580 (p38MAPkinase) did not influence the mRNA expression of the analyzed integrins. The figure displays the normalized transcript levels of the integrins in treated MSC as indicated compared to untreated MSC.
Discussion
In this study, we investigated the attachment of MSC to ECM proteins col-1 and LM-111 in the presence and absence of TGF-β1.
In general, attachment of cells to col-1 and LM-111 can be mediated by integrin receptors α1β1 and α2β1 (col-1 and LM-111), α3β1, α10β1, α11β1 (col-1), α6β1, α7β1, and α9β1 (LM-111). We report that MSC express high amounts of integrin subunits α1, α2, α3, α7, and β1 and medium levels of integrin chains α6, α9, and α11 on the protein level. The expression of integrin α10 was observed by RT-PCR but not by flow cytometry.
The attachment of MSC to col-1 or LM-111 can be abolished by blocking the integrin β1 chain with the antibody 4B4. Furthermore, the attachment of MSC to col-1 is at least partially blocked by the integrin α2 antibody P1E6. The incubation with an antibody against integrin α3 did not block attachment of MSC to col-1, although they express high amounts of this integrin subunit. This indicates that the integrin α2β1 (VLA-2, CD49b/CD29) is a key factor for the attachment of MSC to col-1. The role of other collagen-binding integrin subunits, including α1, α10, and α11, must be further examined in this respect, when function-blocking antibodies become available.
The attachment of MSC to LM-111 could not be blocked by antibodies against integrins α6 and α9, although these integrin subunits are known to be laminin receptors [29]. The MSC strongly expressed integrins α7 and α1. These integrins chains may keep the MSC attached to LM-111, even when other integrins are blocked.
The integrins analyzed in this study are part of the interacting network in the bone marrow and the perivascular niche, anchoring MSC to the ECM of these tissues. Type I collagen is found in the bone marrow stroma, providing contact points for the MSC to attach. MSC themselves can express col-1 [17], thus contributing to the integrity of bone marrow stroma. Both the bone marrow and the basement membranes in blood vessels contain the laminin isoforms LM-411/421 and LM-511/521 [19,21]. Receptors responsible for the attachment to LM-411/421 and LM-511/521 include integrins α2β1, α3β1, α6β1, and α7β1. Showing strong expression of these integrins, MSC may attach well to laminins in the perivascular niche. Bone marrow-derived stromal cells, which contain to a large part MSC, expressed LM-411/421 and 511/521, and thus may contribute to the composition of their own niches [19].
TGF-β1 regulates the integrin expression pattern in MSC and enhances their attachment to col-1. The TGF-β1-induced signaling pathway mainly proceeds via TGF-βRI and subsequently phosphorylation of Smad2. In osteoblasts and vascular smooth muscle cells, an involvement of Smad proteins in the regulation of integrin expression was shown as well [45,46]. In chondrosarcoma cells, the LM-511 receptor integrin ανβ3 is up-regulated by TGF-β via focal adhesion kinase (FAK), PI3 kinase, and NF-κB [47]. It is possible that these components are involved in the regulation of integrins in MSC as well. An influence of TGF-β on the integrin expression pattern was further reported in MG-63 human osteosarcoma cells [35] and alveolar epithelial cells [34]. Enhanced expression of integrin subunits α1, α2, α3, α5, and β1 was reported in WI-38 lung fibroblasts after exposure to TGF-β [48]. In hepatocytes, TGF-β changes the expression of α5β1 and thus diminishes attachment to fibronectin [37]. By modulating the attachment of these cells to the ECM, TGF-β contributes to cell mobilization, organ development, wound healing, and fibrosis.
Recently, we reported that TGF-β1 enhanced the expression of matrix metalloproteinases and IL-16 in SF and also modulated the integrin expression pattern on SF [38,49]. TGF-β enhanced the expression of integrin β1 in SF as well, but in contrast to the MSC, an up-regulation of the integrin α2 or α6 proteins could not be found in SF [38]. However, the TGF-β1-induced modification of the integrin expression pattern in SF is involved in the pathology of rheumatoid arthritis, where TGF-β1 enhances the attachment of the fibroblasts to LM-111. The co-signaling processes between TGF-β1 and integrin receptors induce the expression of matrix-degrading proteases in SF, leading to destruction of the rheumatic tissue [49]. In contrast, the TGF-β1-enhanced attachment of MSC to col-I, which is mediated by integrin α2β1, might contribute to the embedding of MSC into defective tissues during wound healing.
MSC are mobilized into peripheral blood after skeletal muscle injury [50] and acute large skin damage [51], but it is not known whether TGF-β1 is involved in this process. A recombinant TGF-β1 fusion protein with a collagen-binding domain was shown to induce the migration of MSC [52]. But the effect of TGF-β alone on the migration of MSC remains controversial [53,54]. It is possible that additional factors stimulate mobilization of MSC into the peripheral blood, while TGF-β1 is locally secreted and promotes their engraftment into the defective tissue by enhancing the expression of specific integrins.
Migration of MSC can be stimulated by growth factors such as BMP-2, BMP-4, bFGF, EGF, PDGF-ab, and PDGF-bb [53,54]. This process is often coupled to changes in the integrin expression pattern of the target cells. PDGF-ab enhances the expression of integrin β1 in dermal fibroblasts, thus affecting attachment to col-1 and fibronectin [55]. bFGF, which enhances the expression of integrin α10 in MSC [56], regulates migration of MSC in a concentration-dependent manner [53,57]. Hypoxia is another stimulus for the mobilization of MSC into the peripheral blood [58]. This could be interesting in the context of myocardial infarction. After ischemia, the expression of TGF-β is induced in cardiomyocytes. Analysis of the intramyocardial migration of MSC showed that integrin β1 is responsible for the engraftment of MSC in the ischemic myocardium [59].
Thus, TGF-β1 might be an important factor in MSC homing into the damaged tissue by enhancing the expression of integrins on MSC and thereby improving their attachment to col-1. Furthermore, col-1 seems to be a suitable scaffold material for tissue engineering, integrating well into the tissue and keeping the MSC at the site of injury during wound healing under the influence of TGF-β.
Footnotes
Acknowledgments
This project was supported in part by a DFG grant (Ai 16/19). We thank Anis Larbi, Ph.D. for support with the flow cytometry analyses; Tino Felka, MS for optimization of chondrogenic differentiation protocols; Diane Blaurock, MA for her invaluable help in proofreading of the manuscript; and Bernd Rolauffs, MD for help in preparation of the artwork.
Author Disclosure Statement
No competing financial interests exist.