
Abstract
Germ cells are a unique population of cells responsible for transmitting genetic information from one generation to the next. Our understanding of the key mechanisms underlying germ cell development in vivo remains scarce because of insufficient amounts of cell materials available for conducting biological studies. The establishment of in vitro differentiation models that support the generation of germ cells from mouse pluripotent stem cells provides an alternative means for studying reproductive development. The detection and analysis of stem cell-derived germ cells, however, present technical challenges. Methods for determining the developmental stage of germ cells ex vivo, such as gene expression and/or immunochemical analyses are inadequate, frequently necessitating the use of alternative, elaborate methods to prove germ cell identity. We have generated putative oocytes and granulosa cells in vitro from mouse embryonic stem cells and utilized electron microscopy to characterize these cells. Here, we report on the striking ultrastructural similarity of in vitro-generated oocytes and granulosa cells to in vivo oocytes developing within follicles.
Introduction
A
Materials and Methods
Electron microscopy
For transmission electron microscopy (TEM), cells were fixed with 2.5% glutaraldehyde (Merck) in 0.1 M sodium cacodylate buffer (pH 7.4), postfixed in 1% aqueous osmium tetroxide, dehydrated stepwise in a graded ethanol series, and embedded in Epon 812 (Fluka, Buchs, Switzerland). Ultrathin (50 nm) sections were prepared with an ultramicrotome (EM UC6; Leica), stained first with 1% uranyl acetate and then with 3% lead citrate, and subsequently examined using a Zeiss EM 109 electron microscope (Zeiss). Images were taken on 70 mm films (Maco ORT 25c orthochromatic; Hans O. Mahn & Co., Photo Division).
For scanning electron microscopy (SEM), cells were fixed with 2.5% glutaraldehyde in 0.1 M phosphate buffered saline (pH 7.4) and dehydrated stepwise in a graded ethanol series. Samples were dried with 100% ethanol via CO2 in a critical-point apparatus (Balzers). Dried samples were mounted onto aluminum stubs with leit-tabs and coated with gold film to a thickness of 40–50 nm in a sputter coater (Leitz). Cells were viewed under a Hitachi S-530 scanning electron microscope operated in a secondary mode at 20 kV.
Cell culture
Mouse ESC lines gcOct4-GFP (XY) [1] and OG2 (ΔPE-Oct4-GFP, XY; XX) [18] were differentiated into oocytes according to 3 protocols: a previously described serum-containing protocol [1], a serum-containing/serum-free differentiation combination protocol, and a serum-free differentiation protocol. Briefly, for the serum-containing differentiation procedure, cells were grown in tissue culture plates in Dulbecco's modified Eagle's medium containing 4.5 g/L glucose and supplemented with 15% fetal calf serum (Hyclone), 2 mM
RNA isolation
RNA was isolated using the RNeasy Micro Kit (QIAGEN) with on-column DNase digestion as per the manufacturer's instructions. RNA quality was determined using a Nanodrop (ND-1000) spectrophotometer and an Agilent RNA 2100 Bioanalyzer.
For single-cell analysis of oocytes, the Applied Biosystems kit was used for preamplification according to the manufacturer's recommendations.
Reverse transcription–polymerase chain reaction and real-time quantitative reverse transcription–polymerase chain reaction
Two hundred fifty nanograms of RNA was reverse-transcribed to cDNA using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Real-time quantitative TaqMan reverse transcription–polymerase chain reaction (q-RT-PCR) was performed on 7900ht and ht fast devices (Applied Biosystems) with milliQ H2O and TaqMan Universal PCR Master Mix (Applied Biosystems). Raw data were analyzed using SDS 2.3 software (Applied Biosystems). Gene expression was normalized to the mouse housekeeping gene Hprt. Quantification of relative gene expression was calculated using the ΔΔCt method. Three technical replicates were used for each real-time q-RT-PCR reaction; a reverse transcriptase blank and a no-template blank served as negative controls. The following primers were used: Oct4: Mm00658129_gH; Blimp1: Mm00476128_m1; Ddx4: Mm00802445_m1; Figla: Mm00488823_m1; Nobox: Mm00453743_m1; Prdm14: Mm01237813_m1; Sohlh1: Mm01338424_g1; Stella: Mm00836373_g1; Sycp3: Mm00488519_m1.
Isolation of germinal vesicle-stage oocytes from ovaries of adult mice
Eight- to 10-week-old CD1 female mice were superovulated by injection of 7.5 IU pregnant mare serum gonadotropin (PMSG) and, 48 h later, by injection of 7.5 IU human chorionic gonadotropin (hCG). Females were sacrificed by cervical dislocation at 17–18 h post-hCG stimulation. Ovaries were dissected and digested for 10 min at 37°C in 0.5 mg/mL collagenase type 1A (Sigma), washed in PBS, and placed in Hepes-buffered CZB medium containing 1 g/L glucose and 0.1% (w/v) polyvinylpyrolidone (ICN). Follicles were released into the medium by mechanical dissociation of the tissue. Cumulus cells were removed mechanically and oocytes with a clearly visible germinal vesicle were collected, washed 3 times in Hepes-buffered CZB medium, and lysed in RNeasy lyses buffer (RLT) to extract RNA for gene expression analysis or further processed for EM. Mice were maintained and used for experiments according to institutional guidelines.
Results
Granulosa cells and the GC–oocyte interface of ESC-derived follicles
The entry of PGCs into meiosis in vivo and in vitro appears to be a cell autonomous process. Subsequent development of PGCs into competent oocytes and ultimate meiotic arrest during folliculogenesis is dependent on 2-way signaling interactions between the oocyte and surrounding GCs [21,22]. Figure 1A–D shows ESC-derived follicle-like structures. As aggregation of PGCs and granulosa-like cells takes place randomly in this system, we obtained spherical aggregates with centrally located single as well as multiple putative germ cells surrounded by various layers of attached presumptive GCs and extracellular matrix (ECM). Light microscopy revealed germ cell clusters surrounded by loosely attached somatic granulosa-like cells (Fig. 1A). SEM analysis demonstrated follicle-like structures, that is, germ cells surrounded by a dense layer of cuboidal cells (Fig. 1B). Figure 1C shows ESC-derived putative GCs in direct proximity to a zona pellucida-like matrix. The granulosa-like cells exhibit a polygonal shape and a smooth surface with few microvilli (Fig. 1C). SEM and TEM analysis revealed a structure specific for GCs, that is, TZPs (Fig. 1C, 1D, arrow) that extend toward the putative oocyte.
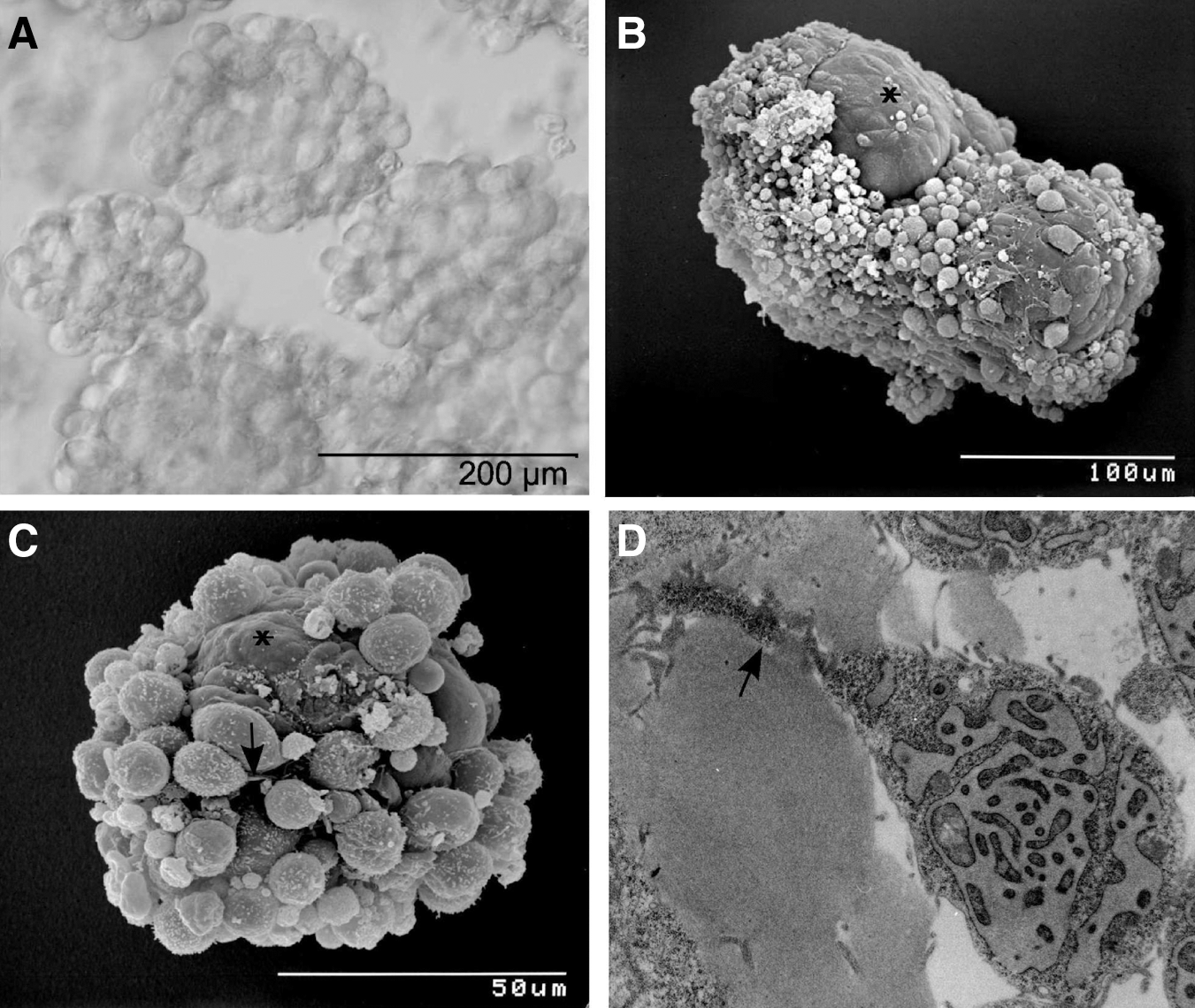
ESC-derived follicle-like structures.
Figure 2A shows a light microscopy image of an ESC-derived follicular structure, with presumptive GCs surrounding the green fluorescent protein (GFP)-positive oocyte-like cell. To analyze the GC–oocyte interface by SEM, in vitro-derived follicles were cracked. Figure 2C and D shows both parts of a cracked aggregate, revealing their morphological similarity to natural follicles. Even though the presumptive oocyte was destroyed during the cracking procedure, a round cavity of 50–60 μm in diameter with residual oocyte cell material is evident (Fig. 2C, D). Multiple layers of GCs surround the cavity. The most central ones exhibiting the typical cuboidal-elongated phenotype of natural polarized cumulus cells, that is, those cells in direct contact to the oocyte (Fig. 2C), whose TZPs orientate toward the oocyte (Fig. 2B). TZPs have been well characterized in many mammals by EM [23,24], and their number and form have been demonstrated to change dynamically during follicular development [12]. To investigate the GC–oocyte interface in more detail, we performed TEM analyses and compared GCs of natural follicles to ESC-derived presumptive GCs. The ultrastructure of TZPs of in vitro-derived GCs appeared to be indistinguishable from the TZPs of their natural counterparts, indicating generation of the interface between the oocyte and GCs, the major control structure for healthy follicle development [12]. ESC-derived GCs of high electron density exhibited long cellular processes containing the same dark fine granulation as the cytoplasm (Fig. 2E). These processes penetrated the matrix, with some reaching a diameter of ∼1 μm, very similar to the branched processes of polarized cumulus cell in natural follicles. The detection of organelles, for example, mitochondria [12,25], within the TZPs of in vitro-derived GCs (Fig. 2F, G) is indicative of recapitulation of folliculogenesis in vitro. Further, the ultrastructural texture of the pale amorphous matrix in direct proximity to the GCs in vitro (Fig. 2E) resembles the zona pellucida of the GC–oocyte interface in vivo (Fig. 2H).
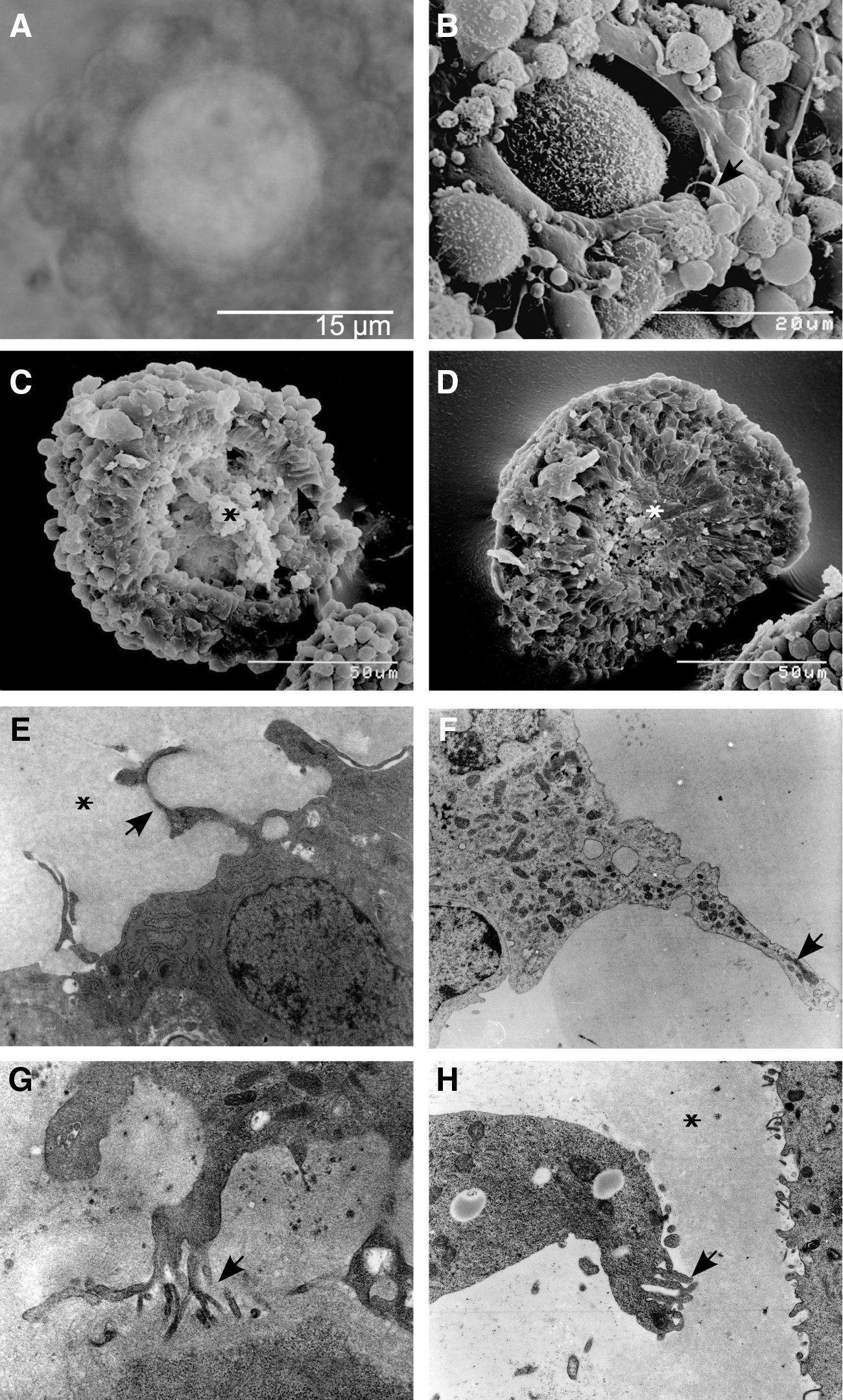
Granulosa cell–oocyte interface of ESC-derived follicles.
Ultrastructural characterization of in vitro-generated GC-like cells revealed additional analogies to characteristics of natural GCs described in the literature, for example, the presence of both dark- and light-appearing GCs. Dark GCs possess a dark background nucleoplasm and background cytoplasm (Fig. 3A, C). The electron density is due to the fine granulation of the cytoplasm, which does not correspond to free ribosomes, as free ribosomes would be much larger and stand distinctly out from the granular background (Fig. 3E, F). The cytoplasm of the GCs appears to be partly vacuolated and is populated by mitochondria, ribosomes, and the most prominent organelle of ESC-derived GCs, the rough endoplasmic reticulum. Cisternae of swollen rough endoplasmic reticulum containing granular material wind through the cytoplasm of dark and light in vitro-derived GCs (Fig. 3A, E) as well as pale in vivo GCs (Fig. 3F). The ribosomes of natural dark and light GCs and ESC-derived GCs are either free or associated with rough endoplasmic reticulum (Fig. 3A, B, E, F). The presence of free ribosomes is a characteristic feature of protein-secreting cells, such as the GCs of growing follicles. The mitochondria of both dark in vivo and in vitro-derived GCs appear irregular and elongated, containing pale cristae and an electron-dense matrix (Fig. 3A, B). The mitochondria of light GCs exhibit an oval shape and have tubular–vesicular cristae (Fig. 3E, F). Lipid droplets filled with a transparent gray substance and empty vacuoles frequently surrounded by a membrane (Fig. 3B, D) are distributed throughout the cytoplasm of in vivo GCs. Vacuolization appears to be a prominent feature of ESC-derived GCs (Fig. 3C), manifested mainly by empty white vacuoles and vacuoles containing a residual gray substance (Fig. 3C). Empty vacuoles appear to be an artifact of sample manipulation, created when the gray substance of lipid droplets is washed out during the dehydration step. The nuclei of both in vitro-derived and in vivo GCs are large and have an irregular contour (Fig. 3A–D). The nucleoplasm of ESC-derived GCs, enclosed by the inner nuclear membrane, contains finely dispersed chromatin and clumps of heterochromatin scattered throughout (Fig. 3A, C, E); the nucleoplasm of in vivo GCs is arranged similarly (Fig. 3B, D, F). Taking these data together, the ultrastructural features of in vitro-derived GCs are indistinguishable from natural GCs. Most importantly, to the best of our knowledge, there is no other cell in the body capable of forming unique and typical TZPs. The observed activated rough endoplasmatic reticulum, indicative for metabolically active and steroid-producing cells, further supports their GC-like identity.
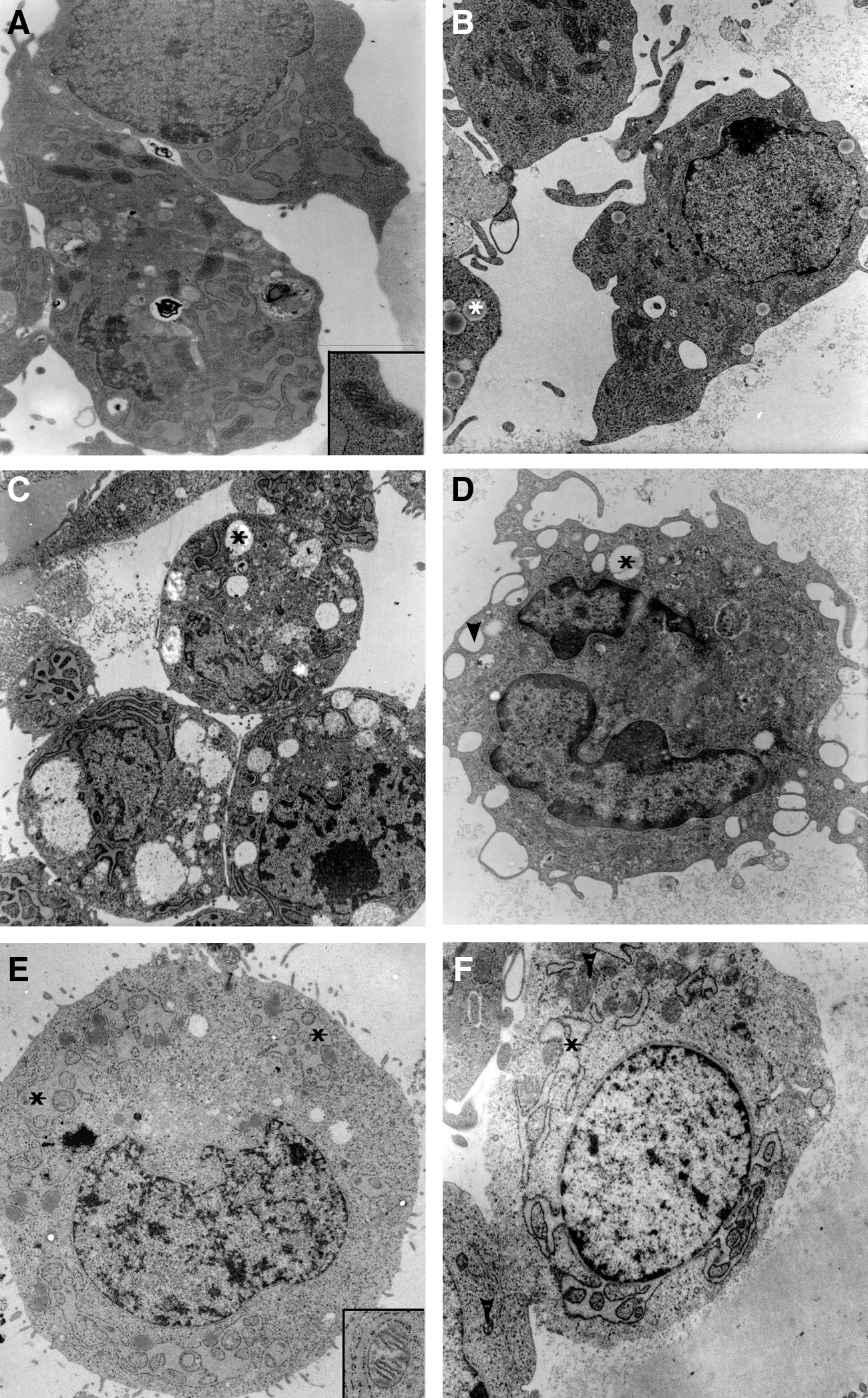
TEM analyses of ESC-derived dark- and light-colored granulosa cells.
ESC-derived oocytes
We have developed a suspension culture system that supports the generation of ESC-derived oocytes of different follicular stages. Independent of the differentiation media used to initiate differentiation of ESCs, we obtained putative oocytes of various developmental stages at different yields. Serum-free conditions appeared best to promote germ cell differentiation, and XY-ESCs yielded a higher percentage of oocytes than XX-ESCs. Expression of specific oocyte marker genes was assessed by qRT-PCR in groups of 3 in vitro-derived oocytes and compared with superovulated germinal vesicle-stage oocytes and ESCs. As expected, the generated oocytes (Fig. 4A) exhibited a gene expression pattern very similar to that of control samples (Fig. 4B). Morphologically, in vitro-generated oocytes (Figs. 4A and 5A) and in vitro-matured oocytes generated from 18.5 days postcoitum (dpc) germ cells under the same culture conditions (Fig. 5C) exhibited a very similar shape, size, and microvillous cell surface (Fig. 5C, D). Interestingly, the surface topography of some in vitro-derived oocytes analyzed by SEM resembled exactly that of unfertilized oocytes in vivo, with the typical sparse and relatively uniform microvillous area and a clear mosaic surface containing a polar microvillous-free region (Fig. 5B–D). TEM analyses of generated oocytes revealed large, slightly eccentric nuclei with a characteristic pale background nucleoplasm (paler than the ooplasm) (Fig. 5E). Free ribosomes appeared individually or arranged in clusters of 5–10 (Fig. 5E, insert). Elongated staples of fibrils not associated with ribosomes were found throughout the cytoplasm (Fig. 5E, insert, F). Oocyte-specific cortical granules containing a characteristic electron-dense center and a pale outer ring (Fig. 5F, arrow) were mostly distributed throughout the cytoplasm and appeared only sporadically at the cell periphery (Fig. 5F and inserts), a distinct feature of immature oocytes. Further, we detected round as well as elongated mitochondria with a dense matrix and pale, irregularly arranged cristae and intermitochondrial vacuoles, all features of natural oocytes. Noteworthy is that the majority of ESC-derived oocytes showed signs of apoptosis and autophagy, which might be indicative of atresia (Fig. 5E, H). The oocyte shown in Fig. 5E lost its spherical shape. Numerous lysosomes, clear vesicles, and autophagosomes as well as dark cytoplasmic structures were detected. The ESC-derived oocyte shown in Fig. 5H contains numerous apoptotic bodies and secondary lysosomes within the dark condensed cytoplasm. Mitochondria and other organelles cannot be identified. The zona pellucida and most of the microvilli at the oocyte surface appear to have been lost. In comparison, oocytes derived from natural 18.5-dpc PGCs grown for 13 days in the same culture medium as in vitro-derived oocytes (Fig. 5G) also exhibit dark-stained mitochondria without cristae, numerous vacuoles, and secondary lysosomes; other organelles cannot be identified. Microvilli either have been lost or have retracted from the zona pellucida. These observations indicate that our culture conditions support the maturation of PGCs into oogonia, but further optimization of these conditions is required for the generation of functional oocytes.
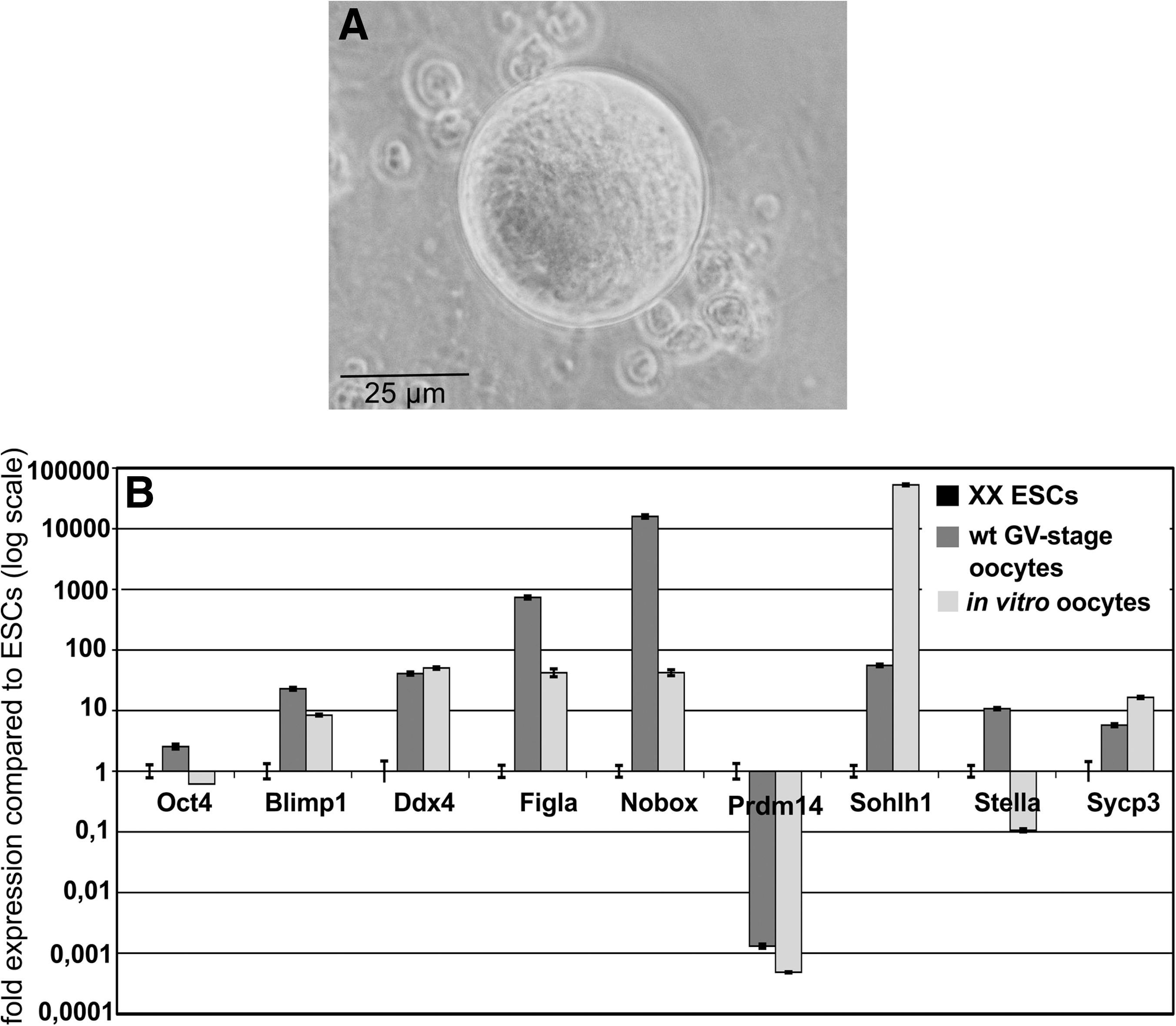
Gene expression of oocyte markers in a pool of 3 in vitro-derived oocytes.

Ultrastructural analysis of in vitro-derived oocytes.
Discussion
The first systematic study of the culture and growth of mouse oocytes in vitro was published by John Eppig in 1977 [26]. Eppig and coworkers demonstrated that mouse oocytes from primordial follicles could be grown and matured in vitro to produce live offspring after fertilization [27,28]. These data marked a major breakthrough in the field of reproductive biology. Based on this success, various in vitro culture systems for female gametes of different mammalian species were established, such as rat [29], hamster [30], cat [31], pig [32], sheep [33], goat [34], cow [35,36], and human [37]. All these maturation systems, including the mouse model, are still under investigation in an effort to broaden our understanding of the complex mechanisms involved in germ cell development and maturation. In this study, we performed comparative ultrastructural analyses of follicle-like structures and bona fide oocytes from mouse ESCs with their natural counterparts. With our data, we supplement the genetic and immunological data obtained by standard analyses with a description of the unique ultrastructural properties of oocytes and their supporting GCs during gametogenesis in vivo. Specifically, we performed SEM analysis on in vitro-derived follicle-like structures and compared their morphology with that of natural mammalian follicles described by Makabe and coworkers in 2006 [13]. Striking morphological similarities indicate that our culture system supports the derivation of germ cells of different developmental follicular stages. During the early growth of follicles, the shape of GCs progressively changes from flattened to cuboidal and to columnar [38] [39]. Well-defined in vitro-derived follicles (Fig. 1B), covered with a layer of densely attached polyhedral cells, form the stratum granulosum, indicating that the follicles are in the late phase of growth. Cracked in vitro-derived follicles, such as the one shown in Fig. 2C and D, exhibit the same morphological features as cracked mammalian follicles, with a residual macerated ooplasm as previously described [13]. We utilized TEM analyses to assess differences in the dynamic structure of TZPs, which form the morphological basis of the oocyte–GC interface and play a crucial role in the structural integrity of developing follicles and maturing oocytes [12]. Many of our in vitro-derived GCs developed microvilli, cytoplasmic invaginations, pseudopodia, and cytoplasmic protrusions (not shown here), which have been associated with cell expansion and are considered indirect signs of GC luteinization both in vitro and in vivo [40]. Further, we observed morphologically different subpopulations of ESC-derived GCs, that is, those exhibiting differences in the darkness of background cytoplasm and nucleoplasm as well as in content and distribution of lipid droplets and rough endoplasmic reticulum. This heterogeneity of a given GC population has also been reported by Nottola and coworkers [16] for in vivo follicular aspirates cultured in vitro. Further, cuboidal GCs with a dark background cytoplasm and nucleoplasm have been found in primordial polyovular and unilaminar follicles of the golden hamster in vivo [41]. We also detected these light and dark GCs in natural mouse follicles (Fig. 3B, D, F). Both dark and light GCs are frequently found in developing endocrine tissues and reflect a change in the complex secretory cycle [41]. The fine dispersion of chromatin and the peripheral patches of heterochromatin, which we found in the nuclei of ESC-derived GCs (Fig. 3A, C, E), are features typical of metabolically active cells [40,42]. Mitochondria of ESC-derived GCs vary in size and shape and have mostly tubular–vesicular cristae (Fig. 3A, E). These types of mitochondria are frequently found in luteinizing GCs in vivo [43] and in human GCs of follicles containing a fertilizable oocyte [16]. In vitro-derived dark GCs contain mitochondria with a dense matrix and pale cristae (Fig. 3A, insert), whereas light GCs contain mitochondria with a pale matrix and dark cristae (Fig. 3E, insert). Makino et al. [44] found an association between mitochondria of human GCs exhibiting a pale matrix with poor fertilization outcome in vitro. Mitochondria with longitudinally oriented cristae have been found in human ovarian carcinoma and in the lungs, liver, and ureter of different mammals [16]. Researchers have hypothesized that this morphological transformation of mitochondria could be due to negative environmental conditions, such as high oxygen concentration, presence of cytostatic drugs or antibiotics and aging [45 –47]. We did not observe such mitochondria in our culture-derived GCs, indicating that we successfully generated functional GCs in vitro. A most prominent feature of in vitro-derived GCs is a rich network of swollen rough endoplasmic reticulum and a well-developed Golgi apparatus. These features indicate that there is ongoing luteinization [40,43,48,49]. Isolated rough endoplasmic reticulum and ribosomes of luteinizing GCs are responsible for synthesizing the enzymes involved in steroid production [50]. The presence of numerous lipid droplets within the cytoplasm of in vitro-derived GCs (Fig. 3C), an early sign of luteinization [50], correlates with active steroidogenesis [16]. Taken together, in vitro-derived GCs exhibit ultrastructural characteristics typical for metabolically active and steroid-producing cells.
The ovarian follicle represents a morphological and functional unit wherein both somatic and germ cells play a pivotal role in follicle maturation and formation of fully competent, fertilizable oocytes [51]. Under appropriate culture conditions, in vitro-matured cumulus-enclosed oocytes of mouse, rat, cow, and sheep have been shown to possess a similar developmental capacity to mature oocytes in vivo [52 –54]. SEM analysis clearly demonstrated that ESC-derived cuboidal-shaped GCs closely surround the oocyte (Fig. 2C, D) and extend long TZPs toward the oocyte (Fig. 1D). In vivo, TZPs are most abundant in preantral follicles. By TEM analysis, we demonstrated that ESC-derived cumulus cells extend typical TZPs, some of which contain mitochondria, an observation consistent with literature [12,25]. Albertini et al. [12] demonstrated the active bidirectional movement of mitochondria within the TZPs by vital staining of intact mouse follicles with MitoTrackerTM, a cell-permeant probe that contains a thiol-reactive chloromethyl moiety for labeling mitochondria. TZPs are an essential requirement for healthy follicle development [12] and the ultrastructure of TZPs of our in vitro-derived GCs demonstrates a profound conformity to that of natural GCs. This data demonstrate the establishment of an interface, that is, the major control site between the oocyte and GCs in vitro. TEM analysis showed that TZPs penetrate a mucous substance, which appears as a gray homogeneous material (Fig. 2E, F). Cumulus cells synthesize and secrete hyaluronic acid as well as produce a hyaluronan-rich ECM matrix [55,56]. The ultrastructure of the ECM matrix of the in vivo cumulus–oocyte complex (Fig. 2H) closely resembles that of the matrix penetrated by the TZPs of the in vitro-derived GCs (Fig. 2E).
Comparative morphological analysis of ESC-derived oocytes and in vivo oocytes revealed remarkable similarities (Fig. 5A–H). The oocyte surface varies greatly from one stage of the cell cycle to the other and even within a given stage, suggesting a close association between the surface characteristics and the maturation status of the oocyte [57]. A mature, unfertilized mouse oocyte is characterized by small blebs and sparse, relatively uniform microvilli [58] as well as a clear mosaic surface, that is, a microvillous membrane with a smooth, microvillous-free polar region. This region is relatively free of organelles [59] and has fewer CGs [60]. SEM analysis of in vitro-derived oocytes from late cultures revealed a mosaic surface topography typical for mature unfertilized oocytes (Fig. 5D).
TEM analysis demonstrated a background nucleoplasm that was paler than the background cytoplasm (Fig. 5E). ESC-derived oocytes exhibited slight vacuolization of ooplasm, a feature typical for immature and aged oocytes [61]. Mature oocytes showed spontaneous or vesicle-fused vacuolization, with the vacuoles usually filled with fluid [62]. Culture-derived oocytes had numerous mitochondria of varied shapes (Fig. 5F). These mitochondria exhibited an electron-dense matrix and pale, irregularly arranged cristae, a typical feature of fetal mouse oocytes [63]. Some mitochondria showed a vacuolated matrix, which has been described for oocytes of polyovular and unilaminar follicles in the golden hamster [41]. Round, oval, and elongated mitochondria are found in mouse growing oocytes [63] and contain large intermitochondrial vacuoles, as shown in Fig. 5F (arrowhead). Numerous fibrillar arrays were detected among the mitochondria of our in vitro-derived oocytes (Fig. 5E, insert). This arrangement is seen in mature oocytes and can only be detected in glutaraldehyde-fixed samples [25]. In the mouse, these fibrils form polysomes and a hexagonal crystalline pattern. The fibrillar bundle seen in Fig. 5E (insert) appeared long, without any pattern, and was not associated with ribosomes as described by Zamboni [25]. Future work should address whether this nonassociation with ribosomes can be attributed to a smaller size, different metabolic activity, or a thinner and atypical zona pellucida of the oocyte.
The formation of cortical granules begins during oocyte maturation [25] and the distribution of cortical granules in the mouse oocyte cortex changes dynamically during meiotic maturation [60]. Cortical granules are round or elliptical in shape, measure 0.2–0.5 μm in diameter, and consist of a highly dense matrix surrounded by a single smooth membrane [25]. As described by Ducibella et al. [60], in immature oocytes, cortical granules are distributed asymmetrically throughout the cortex, whereas in mature oocytes, they are localized mainly to the cortex periphery and are fewer in number. TEM analysis of ESC-derived oocytes showed numerous cortical granules distributed over the entire cortex (Fig. 5F), with only a few granules at the oocyte periphery (Fig. 5F, insert), which is indicative for immature oocytes. The distribution of cortical granules and their competence to undergo cortical reaction are important indicators of oocyte cytoplasmic maturation and provide valuable information about the fertilizable lifespan of the oocyte [60]. Overall, these ultrastructural data suggest that the analyzed ESC-derived oocytes resembled immature and growing wild-type oocytes. We carried out some IVF experiments with oocytes derived under the tested differentiation conditions and at later culture time points, but were not able to achieve fertilization. The immature nature and the low yields of oocytes impeded the appropriate experimental setup for oocyte activation and subsequent IVF and ICSI. For the ultimate proof of functionality, our differentiation conditions need to be further improved to obtain higher yields of oocytes.
In vivo, only few follicles ovulate; the vast majority degenerate and die [64] during atresia, a process that can occur at any stage of follicular development [65]. This process assures adequate oocyte quality, prevents the survival of oocytes with crossover chromosome defects, and limits the number of oocytes within the growing follicle [66]. TEM analysis of in vitro-derived oocytes revealed signs of apoptosis and autophagy (Fig. 5E, H), which are indicative of atresia. As shown in Fig. 5E, ESC-derived oocytes had lost their spherical shape, a structural feature that was used by Escobar et al. to select for atretic rat oocytes. These oocytes showed clear vesicles, numerous lysosomes, and autophagosomes as well as dark cytoplasmic structures, all signs of autophagy [67]. Signs of apoptosis, such as clumps of compact chromatin, apoptotic bodies, and membrane blebbing, were not observed, consistent with the results of Devine and coworkers [65]. Ribbon-like fibril bundles were found around the nucleus, a finding previously reported by Ortiz et al. [68] in atretic oocytes of rat antral follicles. In comparison, in vitro-derived oocytes, such as the one shown in Fig. 5H, exhibited signs of apoptosis. Numerous apoptotic bodies and secondary lysosomes were distributed within the breaking, condensed cytoplasm. Mitochondria could not be identified and the zona pellucida, organelles, and most of the oocyte surface microvilli were lost. Escobar et al. [67] proposed that oocyte cell death probably starts with autophagic degradation of cytoplasmic components, such as mitochondria. It therefore appears that the in vitro-derived oocyte shown in Fig. 5E was analyzed at an initial phase of cell death, whereas the oocyte shown in Fig. 5H was isolated at an advanced stage of apoptotic cell death. Natural mouse oocytes derived from 18.5-dpc PGCs (Fig. 5G) exhibited similar features when grown under the same culture conditions, indicative of ongoing atresia. All oocytes shown in Fig. 5 were freely floating in the culture media at the time of collection, that is, had already been extruded from the follicle-like structures. At this time, it is not known whether the autophagic and/or apoptotic signs shown by our in vitro-derived oocytes are a consequence of the culture conditions or are caused by apoptotic-inducing stimuli present in the cellular milieu.
Although the derivation and culture of female germ cells from pluripotent stem cells in vitro may lead to ultrastructural alterations in oocytes, this study demonstrates that a simple in vitro culture system can create the essential components required for oocyte development: oocytes, an ECM-based interface, and granulosa-like cells. Future efforts to optimize our in vitro differentiation conditions should bring us closer to our ultimate goal of generating fully functional oocytes and demonstrate whether all key events of germ cell development in vivo can be truthfully recapitulated in vitro.
Footnotes
Acknowledgments
The authors thank J. Lange, M. Sinn, C. Ortmeier, and G. Verberk for providing technical assistance, L. Gentile and T. Esteves for providing helpful discussions, and the Department of Zoology (University of Osnabrück) for granting our group access to their electron microscope. This work was supported by the Max Planck Society and the DFG grant for Research Unit Germ Cell Potential (FOR 1041).
Author Disclosure Statement
The authors declare that no competing financial interests exist.